
100份朝天椒的农艺性状和SRAP标记遗传多样性分析
2018-09-05 13:00:16曾绍贵朱邦彤罗木旺邱胤晖林淑婷
江苏农业学报 2018年4期
曾绍贵, 朱邦彤, 罗木旺, 邱胤晖, 罗 英, 林淑婷
(1.三明市农业科学研究院,福建 沙县 365509; 2.沙县农业科学研究所,福建 沙县 365509)
朝天椒(Capsicumannuum.L)辣味足、产量较高,是优良的鲜食和加工蔬菜。朝天椒是按果实着生状态,对椒果朝天或斜朝天生长这一类群辣椒的统称,包括分类上辣椒栽培种的5个变种:簇生椒、圆锥椒(小果型)、长辣椒(短指形)、樱桃椒和灯笼椒[1]。
种质资源的遗传多样性分析是育种工作中的重要内容,在实际工作中育种家往往凭借经验进行品种的选配,这成为限制育种精准性的重要因素[2]。在常规育种中,由于育种开展研究时间长和选育技术的同质化及基础育种材料在各育种单位间广泛交换,使得育成品种的相似性很大。应用分子标记可以对种质资源、杂交亲本和后代个体进行大规模的基因型鉴定,构建遗传图谱,并与表型数据建立联系,从而大大提高育种效率和规模。利用分子标记对种质资源进行亲缘关系分析,准确性高、操作简单,在众多大田作物中得到了广泛的应用。马荣丽等[3]对225份类型多样的辣椒资源的11个表型性状进行遗传多样性分析。耿广东等[4]对92份辣椒材料的11个表型性状进行聚类分析,将一些来源相同或相近的聚在一起。张素勤等[5]通过对贵州省不同地区的61份辣椒种质资源进行表型和SRAP分析,以探索贵州辣椒种质的亲缘关系和遗传多样性。许先松等[6]采用形态学标记与序列相关扩增多态性(SRAP)分子标记相结合的手段,分别对49份和72份辣椒资源的遗传多样性及亲缘关系进行了分析。
为了提高亲本的选择效率,提高朝天椒新品种选育进程中的预见性和可靠性,在现有的朝天椒种质资源基础上,有必要对其进行进一步拓展。本试验以100份朝天椒材料为研究对象,采用SRAP标记技术和传统表型聚类分析对朝天椒进行亲缘关系的鉴定,分别从朝天椒的形态差异及起源进化上进行分析,为配制杂交组合、提高新品种选育效率提供理论依据。
1 材料与方法
1.1 试验材料
试验选取的100份朝天椒是三明市农业科学研究院蔬菜研究所近年来收集和自主培育的品种,分别来自中国(湖南、河南、福建等)、韩国、印度等主要国家(地区)的特色品种(表1)。试验于2016年8月-2017年2月在福建省三明市农业科学研究院蔬菜研究所试验田进行。试验于2017年1月15日按品种单株随机取材,采取新鲜嫩叶,每个品种单株进行3次重复,保存-80 ℃冰箱备用。试验于2017年2月20日分别对100份朝天椒具有本品种典型性状的10株植株进行主要农艺性状的调查。为保证实验的客观性,所有供试材料采用编号,试验结果统计后才将编号与供试材料名称对应。
1.2 试验方法
1.2.1 主要农艺性状观察记载与朝天椒主要农艺性状表型分析 表型性状调查和数据采集方法参考《植物新品种特异性、一致性和稳定性测试指南——辣椒》[7]进行,每份材料随机调查记录生长正常植株10株,共测量11个表型性状(表2)。
为了揭示各品种性状间的相互关系,为朝天椒新品种选育以及种质资源创新提供参考信息,对收集记载的100份朝天椒农艺性状数据,利用SPSS Statistics 19软件进行相关系数的计算,主成分分析和数量性状的表型变异分析;利用Hierarchical cluster进行分层聚类,构建100份朝天椒农艺性状表型层次聚类谱系图。
1.2.2 DNA提取 采用改良的CTAB法提取100份朝天椒基因组DNA。用1.5%琼脂糖检测质量,用紫外分光光度计将DNA质量浓度标准化为50 ng/μl。
1.2.3 SRAP反应体系及扩增程序 SRAP-PCR总反应体积为20 μl:2 μl 10×Buffer,0.4 μl dNTP(10 mmol/L),上下游特异性随机引物组合各0.5 μl(50 pmol/μl),0.6 μlTaqDNA聚合酶(1.5 U/μl),1 μl模版DNA(50 ng/μl)和15.0 μl无菌ddH20。SRAP-PCR反应程序为:94 ℃条件下预变性3 min;94 ℃变性40 s,56 ℃退火30 s,72 ℃复性5 min,40次循环,最后72 ℃延伸8 min,4 ℃条件下停止反应。
1.2.4 引物筛选 利用表型差异较大的朝天椒材料,进行SRAP特异引物组合的筛选,鉴定出条带清晰、多态性高的核心引物进行遗传多样性分析,淘汰扩增效果差、带型不易辨认的引物组合。
1.2.5 数据处理与统计分析 利用Microsoft Excel 2016和Ntedit1.2软件分析相关序列扩增多态性获得的特异条带,对于凝胶电泳图上的某个相同迁移位置上有DNA条带赋值为1,无特异性条带的赋值为0,缺项赋值为9。利用NTsys2.10e软件,根据Nei-Li计算相似系数,按非加权配对算数平均法(UPGMA)进行SAHN聚类分析,最后利用Tree plot程序生成构建100份朝天椒的亲缘关系树状图。
表1供试材料编号与名称
Table1Thenumberandnameofmaterial
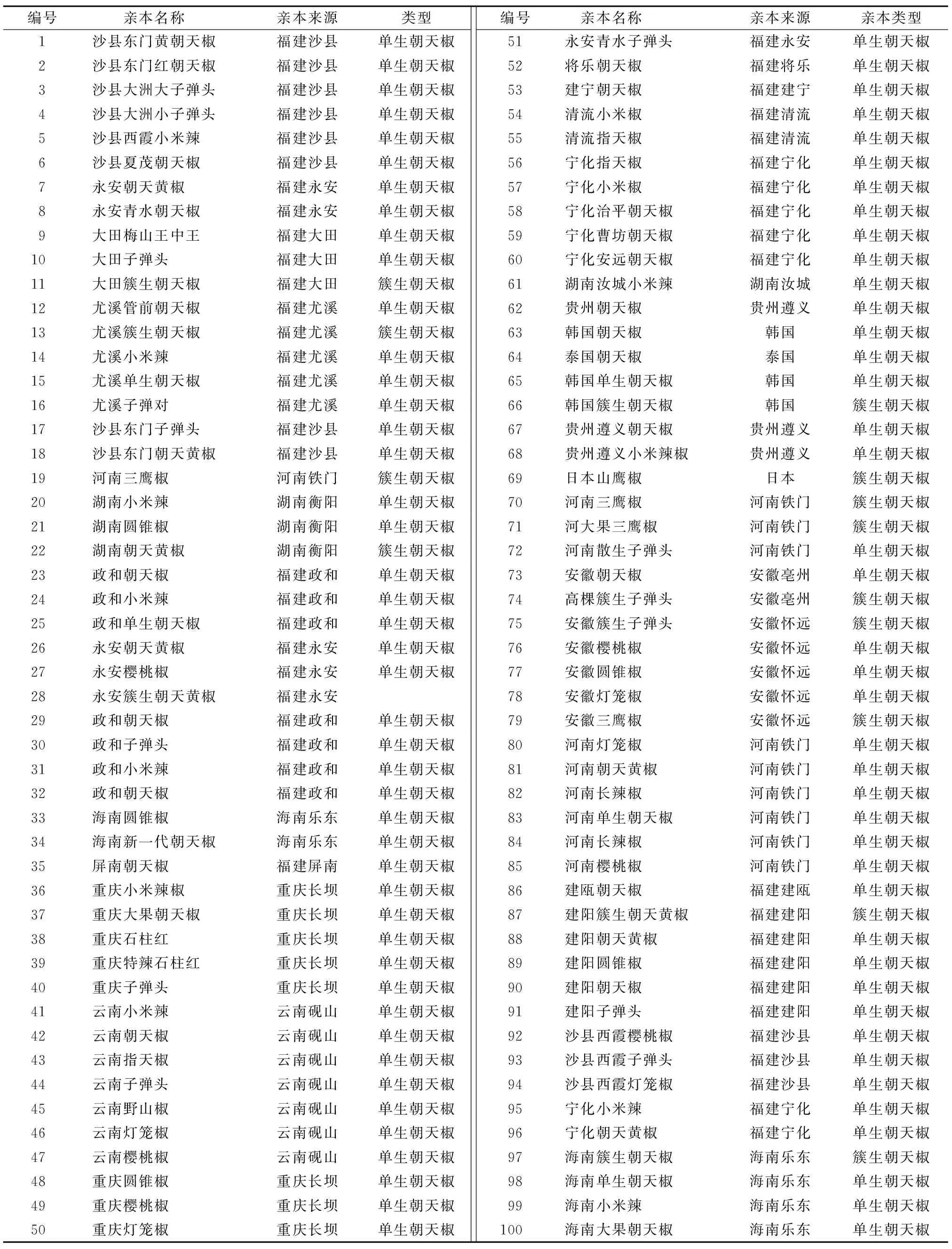
编号 亲本名称亲本来源类型编号 亲本名称亲本来源亲本类型1沙县东门黄朝天椒福建沙县单生朝天椒51永安青水子弹头福建永安单生朝天椒2沙县东门红朝天椒福建沙县单生朝天椒52将乐朝天椒福建将乐单生朝天椒3沙县大洲大子弹头福建沙县单生朝天椒53建宁朝天椒福建建宁单生朝天椒4沙县大洲小子弹头福建沙县单生朝天椒54清流小米椒福建清流单生朝天椒5沙县西霞小米辣福建沙县单生朝天椒55清流指天椒福建清流单生朝天椒6沙县夏茂朝天椒福建沙县单生朝天椒56宁化指天椒福建宁化单生朝天椒7永安朝天黄椒福建永安单生朝天椒57宁化小米椒福建宁化单生朝天椒8永安青水朝天椒福建永安单生朝天椒58宁化治平朝天椒福建宁化单生朝天椒9大田梅山王中王福建大田单生朝天椒59宁化曹坊朝天椒福建宁化单生朝天椒10大田子弹头福建大田单生朝天椒60宁化安远朝天椒福建宁化单生朝天椒11大田簇生朝天椒福建大田簇生朝天椒61湖南汝城小米辣湖南汝城单生朝天椒12尤溪管前朝天椒福建尤溪单生朝天椒62贵州朝天椒贵州遵义单生朝天椒13尤溪簇生朝天椒福建尤溪簇生朝天椒63韩国朝天椒韩国单生朝天椒14尤溪小米辣福建尤溪单生朝天椒64泰国朝天椒泰国单生朝天椒15尤溪单生朝天椒福建尤溪单生朝天椒65韩国单生朝天椒韩国单生朝天椒16尤溪子弹对福建尤溪单生朝天椒66韩国簇生朝天椒韩国簇生朝天椒17沙县东门子弹头福建沙县单生朝天椒67贵州遵义朝天椒贵州遵义单生朝天椒18沙县东门朝天黄椒福建沙县单生朝天椒68贵州遵义小米辣椒贵州遵义单生朝天椒19河南三鹰椒河南铁门簇生朝天椒69日本山鹰椒日本簇生朝天椒20湖南小米辣湖南衡阳单生朝天椒70河南三鹰椒河南铁门簇生朝天椒21湖南圆锥椒湖南衡阳单生朝天椒71河大果三鹰椒河南铁门簇生朝天椒22湖南朝天黄椒湖南衡阳簇生朝天椒72河南散生子弹头河南铁门单生朝天椒23政和朝天椒福建政和单生朝天椒73安徽朝天椒安徽亳州单生朝天椒24政和小米辣福建政和单生朝天椒74高棵簇生子弹头安徽亳州簇生朝天椒25政和单生朝天椒福建政和单生朝天椒75安徽簇生子弹头安徽怀远簇生朝天椒26永安朝天黄椒福建永安单生朝天椒76安徽樱桃椒安徽怀远单生朝天椒27永安樱桃椒福建永安单生朝天椒77安徽圆锥椒安徽怀远单生朝天椒28永安簇生朝天黄椒福建永安78安徽灯笼椒安徽怀远单生朝天椒29政和朝天椒福建政和单生朝天椒79安徽三鹰椒安徽怀远簇生朝天椒30政和子弹头福建政和单生朝天椒80河南灯笼椒河南铁门单生朝天椒31政和小米辣福建政和单生朝天椒81河南朝天黄椒河南铁门单生朝天椒32政和朝天椒福建政和单生朝天椒82河南长辣椒河南铁门单生朝天椒33海南圆锥椒海南乐东单生朝天椒83河南单生朝天椒河南铁门单生朝天椒34海南新一代朝天椒海南乐东单生朝天椒84河南长辣椒河南铁门单生朝天椒35屏南朝天椒福建屏南单生朝天椒85河南樱桃椒河南铁门单生朝天椒36重庆小米辣椒重庆长坝单生朝天椒86建瓯朝天椒福建建瓯单生朝天椒37重庆大果朝天椒重庆长坝单生朝天椒87建阳簇生朝天黄椒福建建阳簇生朝天椒38重庆石柱红重庆长坝单生朝天椒88建阳朝天黄椒福建建阳单生朝天椒39重庆特辣石柱红重庆长坝单生朝天椒89建阳圆锥椒福建建阳单生朝天椒40重庆子弹头重庆长坝单生朝天椒90建阳朝天椒福建建阳单生朝天椒41云南小米辣云南砚山单生朝天椒91建阳子弹头福建建阳单生朝天椒42云南朝天椒云南砚山单生朝天椒92沙县西霞樱桃椒福建沙县单生朝天椒43云南指天椒云南砚山单生朝天椒93沙县西霞子弹头福建沙县单生朝天椒44云南子弹头云南砚山单生朝天椒94沙县西霞灯笼椒福建沙县单生朝天椒45云南野山椒云南砚山单生朝天椒95宁化小米辣福建宁化单生朝天椒46云南灯笼椒云南砚山单生朝天椒96宁化朝天黄椒福建宁化单生朝天椒47云南樱桃椒云南砚山单生朝天椒97海南簇生朝天椒海南乐东簇生朝天椒48重庆圆锥椒重庆长坝单生朝天椒98海南单生朝天椒海南乐东单生朝天椒49重庆樱桃椒重庆长坝单生朝天椒99海南小米辣海南乐东单生朝天椒50重庆灯笼椒重庆长坝单生朝天椒100海南大果朝天椒海南乐东单生朝天椒
表2测量的农艺性状
Table2Theagronomictraitsmeasuredinthisstudy

标号名称说明备注X1株高 (cm)地面植株基部至全株最高处X2开展度 (cm)植株左右宽度X3茎粗 (cm)第一朵花下横径X4叶长 (cm)叶片的长度于植株中部随机取10片叶测量X5叶宽 (cm)叶片的宽度X6果纵径 (cm)果最长部分纵径于盛收期随机取10个果实测量X7果横径 (cm)果最长部分横径X8果肉厚 (cm)果肉的厚度在果实的果肩部横切后测量X9果柄长 (cm)果实和茎连接的部位的长度X10单果质量 (g)成熟期的果实质量X11单株果数单株上所有的果实数
2 结果与分析
2.1 朝天椒主要农艺性状表型遗传多样性分析
2.1.1 朝天椒表型变异分析 统计分析结果表明(表3),表型性状存在不同程度的变异,变异系数平均值为24.80%。
2.1.2 朝天椒表型相关性分析 通过对朝天椒11个表型性状间的相关性分析可知(表4),株高、果横径、果肉厚、茎粗和开展度间呈显著正相关,叶长、叶宽、果柄长和株高间呈极显著正相关,开展度与单果质量间呈极显著正相关,叶宽、果柄长、单果质量和叶长呈极显著正相关,叶宽和单果质量呈显著相关,叶宽和果柄长呈极显著正相关,果横径和果纵径呈显著负相关,果柄长和果纵径呈极显著正相关,单果质量和果纵径呈显著相关,果肉厚、单果质量和果横径呈极显著正相关,果横径和果柄长呈极显著负相关,果肉厚和果柄长呈显著负相关,果肉厚和单株果数呈极显著负相关,果肉厚和单果质量呈极显著正相关,单果质量和单株果数呈极显著负相关。由此可以推断,果肉较厚、果实横径和纵径较大的品种,对产量和单果质量影响较大。
表3朝天椒资源数量性状的表型变异分析
Table3PhenotypicvariationanalysisofquantitativecharactersinCapsicumannuum

形态学性状极小值极大值均值标准差株高 (cm)30.095.058.94011.930开展度 (cm)26.075.052.70010.520茎粗 (cm)0.71.50.9710.178叶长 (cm)6.521.511.9292.601叶宽 (cm)3.09.05.6101.186果纵径 (cm)0.613.56.0031.895果横径 (cm)0.83.51.9110.452果肉厚 (cm)0.10.30.1890.044果柄长 (cm)1.24.02.3100.555单果质量 (g)3.116.67.4672.569单株果数12.072.032.96011.171
表4朝天椒各性状间的相关系数
Table4CorrelationcoefficientofmainagronomiccharactersinCapsicumannuum
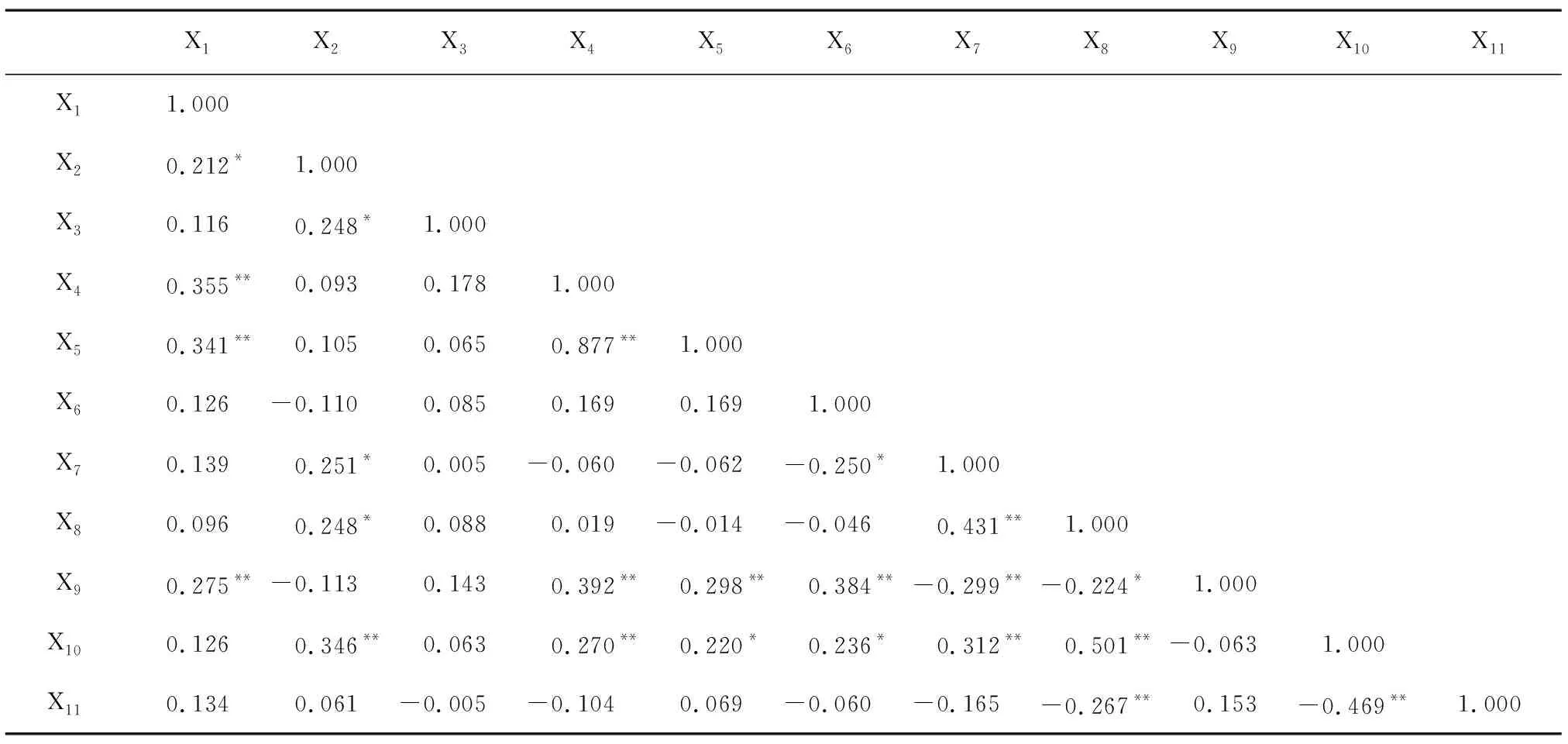
X1X2X3X4X5X6X7X8X9X10X11X11.000X20.212*1.000X30.1160.248*1.000X40.355**0.0930.1781.000X50.341**0.1050.0650.877**1.000X60.126-0.1100.0850.1690.1691.000X70.1390.251*0.005-0.060-0.062-0.250*1.000X80.0960.248*0.0880.019-0.014-0.0460.431**1.000X90.275**-0.1130.1430.392**0.298**0.384**-0.299**-0.224*1.000X100.1260.346**0.0630.270**0.220*0.236*0.312**0.501**-0.0631.000X110.1340.061-0.005-0.1040.069-0.060-0.165-0.267**0.153-0.469**1.000
X1~X11见表2。**表示在0.01水平相关性显著;*表示在0.05水平相关性显著。
2.1.3 朝天椒表型聚类分析 100份朝天椒材料的11个农艺性状聚类分析结果见图1,在欧式距离为20处,100份朝天椒材料划分为2大类群:第一类群包含42份材料,第二类包括58份材料。在欧式距离为14处则被分为4个群组,分别依次命名为Ⅰ、Ⅱ、Ⅲ和Ⅳ。
第Ⅰ群组包含18份材料(85、96、83、84、7、88、78、91、77、86、87、44、89、95、92、43、94和93)。第Ⅱ群组包含24份材料(60、70、65、33、53、39、61、64、74、29、30、31、45、90、56、79、72、81、17、76、36、50、35和6)。第Ⅲ群组包含5份材料(58、69、59、67和68)。第Ⅳ群组包含53份材料(57、82、18、51、10、32、49、14、9、16、34、20、38、52、11、54、13、47、12、22、48、55、73、71、42、75、41、1、26、23、62、40、46、97、98、99、100、3、27、66、28、80、8、37、15、19、4、5、2、21、25、24和63)。
2.2 朝天椒SRAP遗传多样性分析
2.2.1 分子标记在朝天椒群体中扩增产物的多态性分析 利用4份表型差异较大的朝天椒DNA样品对SRAP引物组合进行筛选,最终确定8对引物序列组合(表5)用于SRAP-PCR反应以研究100份朝天椒材料的遗传多样性。PCR扩增累计扩增出76条谱带,其中多态性谱带有61条,总的多态性比率为80.3%,各引物多态性比率为 71.4%~87.5%。
2.2.2 朝天椒遗传多样性分析及聚类分析 根据朝天椒种质资源材料的SRAP扩增结果,按非加权配对算数平均法(UPGMA)进行SAHN聚类分析,得到亲缘关系聚类分析树状图(图2)。以遗传相似系数0.77为阀值,利用NTsys2.10e软件,根据Nei-Li计算的相似系数为 0.77~1.00,在相似系数为0.815处,100份朝天椒材料被明显地分为2大类:第一类包括47份材料,第二类包括53份材料。在遗传相似系数为0.810处,朝天椒材料被分为7个群组,分别依次命名为Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ和Ⅶ。
第Ⅰ群组包含40份材料(1、2、4、16、17、18、3、19、14、15、13、5、25、27、26、28、31、66、76、37、73、38、64、62、63、53、89、6、7、41、42、43、61、54、49、51、52、50、88和90)。第Ⅱ群组包含5份材料(65、74、75、77和87)。第Ⅲ群组包含2份材料(29和39)。第Ⅳ群组包含37份材料(8、11、12、35、9、10、20、81、80、82、21、24、22、23、40、44、57、58、59、67、68、69、79、93、95、45、84、47、48、55、56、91、92、94、60、83和86)。第Ⅴ群组包含10份材料(30、32、33、34、36、46、97、98、99和100)。第Ⅵ群组包含3份材料(71、72和78)。第Ⅶ群组包含3份材料(70、96和85)。
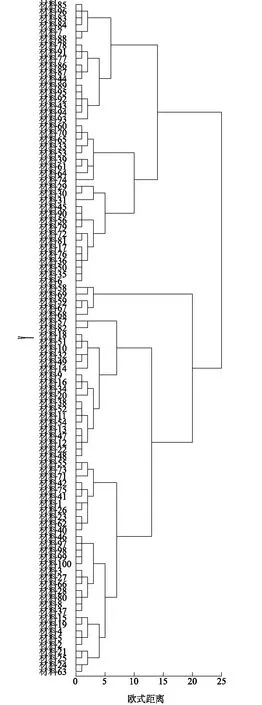
材料1~100见表1。图1 100份朝天椒材料表型性状聚类图Fig.1 Cluster graph of phenotypic traits in Capsicum annuum
表5不同SRAP引物组合和100份朝天椒种质中扩增出的多态性
Table5Polymorphismin100samplesofCapsicumannuumamplifiedbydifferentSRAPprimers
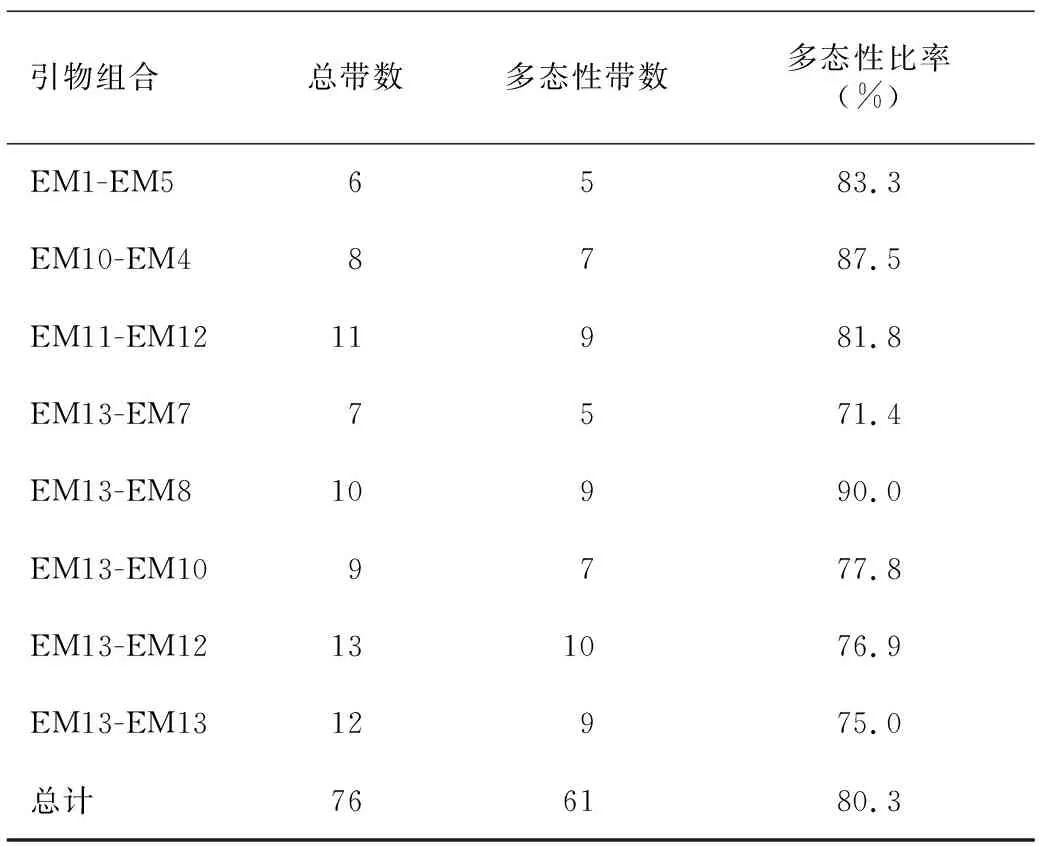
引物组合总带数多态性带数多态性比率(%)EM1-EM56583.3EM10-EM48787.5EM11-EM1211981.8EM13-EM77571.4EM13-EM810990.0EM13-EM109777.8EM13-EM12131076.9EM13-EM1312975.0总计766180.3
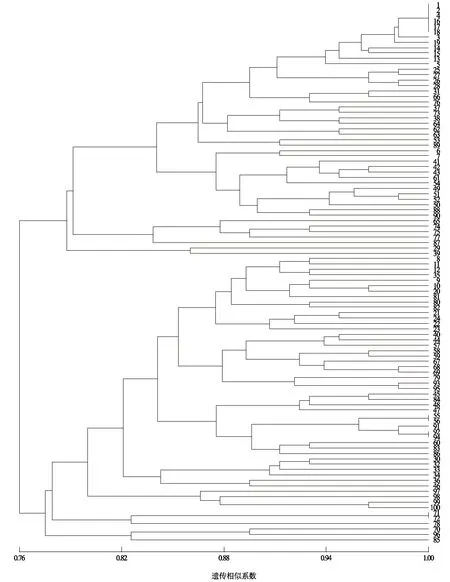
图2 100份朝天椒种质资源材料的SRAP聚类分析Fig.2 SRAP cluster analysis of Capsicum annuum
3 讨 论
植物种质资源是发展农业生产,开展作物育种和进行生物技术研究的物质基础[8]。植物的农艺性状是遗传因素和环境因素相互作用表现出的结果。
3.1 表型相关分析
表型性状兼具稳定性和变异性,既是遗传变异的特征,也易受环境的影响[9]。叶长和叶宽相关系数最大(0.877),呈极显著正相关,说明叶长较长的朝天椒品种其叶宽也较宽。单株果数和单果质量是朝天椒产量构成的重要因素,而单株果数与果肉厚和单果质量呈极显著负相关,单果质量与开展度、叶长、果横径、果肉厚呈极显著正相关,说明朝天椒产量是一个较为综合的指标。因此,从高产优质的育种目标出发,良种选育过程虽然产量受到诸多因素的影响,但也应优先选择单株果数、果肉厚、单果质量这3个性状指标。
3.2 表型变异分析
株高的标准差与方差最大,变异系数为 10%~100%,为中等变异,引进的材料可以为品种改良提供用处。单果质量的变异系数达到最大值,而变异系数值较大的性状可以作为遗传育种改良的首选特征,通过有目的的良种选育较容易获得单果质量较大的品种。
3.3 表型聚类分析
在第Ⅰ群组中编号为83、84、85、86、87、88、89这7份材料均由同一个系统选育的品种以及衍生自交分离后育成的品种,其农艺性状(株高、开展度和茎粗等)表现较为相似,在朝天椒果实色泽上表现不同(红色、黄色)以及果形指数表现不同。第Ⅰ群组中的沙县西霞樱桃椒、沙县西霞子弹头、沙县西霞灯笼椒、宁化小米辣、宁化朝天椒属于三明地区引进的亲本材料,在聚类分析中与河南朝天椒分在同一个群组,说明三明地区部分朝天椒材料是通过引进河南的亲本,通过人工选育后,进行多代自交纯化而成的。第Ⅲ群组中的宁化治平朝天椒、宁化曹坊朝天椒与贵州遵义朝天椒、贵州遵义小米辣、日本山鹰椒分在同一群组,这些材料的朝天椒主要农艺性状较为相似,例如单果质量、果纵径等方面。第Ⅳ群组中的三明大田、三明尤溪、三明将乐朝天椒材料分在同一群组,表现为生态环境相似,地理来源相邻地区的遗传关系较近。海南簇生朝天椒、海南单生朝天椒、海南小米辣、海南大果朝天椒分在同一群组,这些材料叶长、株高、开展度等农艺性状较为相似。第Ⅱ群组的24份样本材料只是将农艺性状相似的朝天椒材料聚在一起,没有体现出地理相似性。
3.4 SRAP分析
每对引物扩增出的谱带数为6至13之间,平均每对引物扩增出的条带为9.5个多态性位点,扩增位点的分子量主要分布于 200~2 000 bp。EM13和EM8标记的谱带存在明显多态性分布,能够较好地鉴别出不同的朝天椒DNA材料,EM13和EM8引物组合共扩增出10条谱带,其中9条属于多态性位点,多态性比率达90%。从引物组合EM13和EM8对100份朝天椒的SRAP扩增结果可以看出,有明显不同片段大小的光亮条带分布,不同品种材料之间存在差异,也有部分条带趋于一致,可能是由于选取的100份朝天椒材料有部分是通过杂交优势育种后自交提纯出的材料,由于基因组高度一致,因此经引物筛选出的条带大体一致。
3.5 SRAP聚类分析
SRAP聚类分析结果与表型聚类结果不同的是许多来源同一地区(县、市)的朝天椒材料并没有聚为同一类,这可能是由于在系统选育的过程中,不同种质材料相互间进行多个亲本复合杂交以及多项选择,产生了较大的遗传变异所致。第Ⅴ群组包括10份种质材料,这些材料具有高度的一致性,由于这些种质材料是由同一个单株分离出的,在人工选育时可能是更多地选择了父本的优良性状而较少选择母本性状造成的。经大田观察,这几个新品种在物候期等多个形状上与父本非常接近,这一事实也很好地解释了上述推测。这些种质材料的单株果数较少,对应的折算产量也较低,但其生长势较旺盛,可作为强株复壮的材料。
如今SRAP分子标记技术已经应用在辣椒的种质资源遗传多样性研究当中,本试验发现其多态性较高,能较准确聚类100份朝天椒材料,为解决朝天椒育种过程中亲本选择效率低、育种进程慢的技术问题提供了借鉴。
