
基于晋汾52和晋谷21解析谷子NBS类型抗病基因变异
2018-09-05 11:12韩彦卿郑洁武彩娟元香梅韩渊怀
山西农业大学学报(自然科学版) 2018年9期
韩彦卿,郑洁,武彩娟,元香梅,韩渊怀*
(1.山西农业大学 农学院,山西 太谷 030801;2.山西农业大学 农业生物工程研究所,山西 太谷 030801)
谷子(Setariaitalica(L.) P. Beauv.),去壳后称为小米,是起源于我国黄河流域的古老作物,由青狗尾草驯化而来,现广泛栽培于欧亚大陆的温带和热带地区[1]。2015年我国谷子栽培面积1 200万亩,占世界80%,总产量占90%,位居杂粮作物之首。然而,近几年来,谷子各种病害发生和流行,严重降低了谷子的产量和品质。例如,谷子白发病在大发生年份可造成产量减产70%以上[2]。由此可见,选育抗病高产优良谷子品种是解决谷子免受病害侵袭的首选[3]。基于本研究室田间多年调查统计发现,晋谷21虽然品质优良,但田间表现极易感白发病。然而其亲本晋汾52在田间表现为抗病。这为我们研究2个品种间的抗感分子机制提供了非常好的研究材料。
随着测序技术的突飞猛进和发展,基因组重测序技术已经成为发掘重要抗病相关基因的重要手段,已经广泛应用到各个研究领域[4~6]。Jia等2013年对916份来自世界各地的谷子核心资源进行了基因组重测序,分析获得了200多万个SNP标记和50万个插入缺失(InDel)的变异图谱,为谷子的遗传研究和抗病改良奠定了坚实基础[7]。施阳选用高抗白粉病片段代换系和高感的受体亲本2个黄瓜品种进行全基因组重测序,鉴定出了具有LRR结构域的抗病基因,初步明确了黄瓜抗白粉病机制[6]。本研究基于本实验室对晋谷52 和晋汾21全基因组重测序的基础上,依据已鉴定NBS家族基因的SNP、InDel、CNV和SV位点,对NBS(核苷酸结合位点)类抗性基因进行比较分析,筛选出6个突变位点较多,接着克隆差异较大的基因并测序分析,拟挖掘鉴定出导致晋谷21及其亲本晋汾52抗病性差异的关键基因,旨在为谷子抗白发病分子辅助育种的培育提供理论指导。
1 材料与方法
1.1 试验材料
本试验选用感病谷子品种晋谷21(JG21),抗病谷子品种晋汾52(JF52),对照品种豫谷1号(YG1)为试材。3个品种在温室盆栽种植,谷子生长条件模拟田间条件进行,待谷子长到3~5叶期时,取其幼嫩叶片,立即放入液氮速冻备用。
1.2 试验方法
1.2.1 谷子基因组DNA提取及PCR扩增
2个品种基因组DNA的提取,采用北京康为世纪 CW0553植物基因组提取试剂盒。所提取DNA的纯度和质量分别用Nanodrop2000检测核酸浓度和纯度,1.0%琼脂糖凝胶电泳检测DNA质量。用Primer Premier5.0设计PCR扩增引物。引物合成由生工生物工程(上海)股份有限公司北京DNA合成部合成。PCR扩增体系:采用50 μL的总反应体系,其中2×GC PCR Buffer 25 μL,dNTP Mixture 8 μL,LA Taq酶0.5 μL,正反向引物各2 μL,模板DNA 2 μL ddH2O 10.5 μL。PCR程序设置为:94 ℃预变性5 min,94 ℃变性30 sec,退火温度57~59 ℃不等1 min,72 ℃延伸1~4 min,72 ℃复延伸10 min,循环38次,4 ℃下保存(表1)。
1.2.2 NBS类型基因比对分析
基于前期基因组重测序及测序拼接的基础上,本实验室获得了完整的NBS类型基因序列。随后利用序列比对软件ClustalX对基因的核苷酸和氨基酸序列进行同源比对,寻找差异位点。并对含有突变位点进行多次测序验证,确认确实是由于辐射诱变导致的突变。蛋白特性分析采用Compute pI/Mw软件,蛋白的二级结构分析用SOPMA和ScanProsite软件,三级结构预测和分析分别软件SignalP和ProtScale软件。
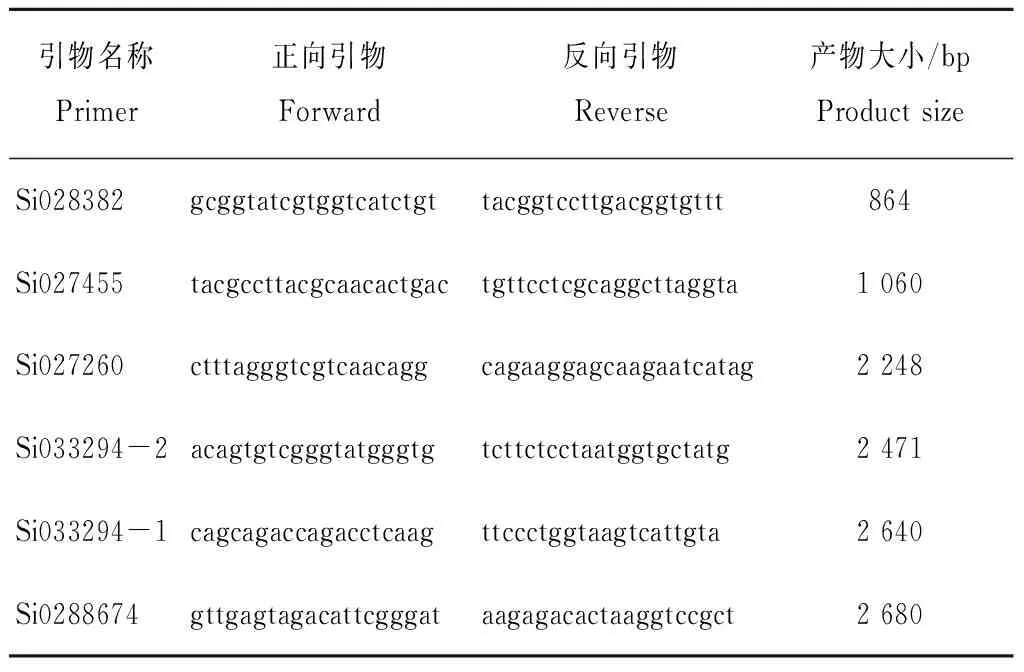
表1 引物序列Table 1 Primer sequences
2 结果与分析
2.1 NBS类基因的扩增片段比较
利用上述设计的6对PCR引物,对3个不同谷子品种进行PCR扩增,分析扩增条带发现,4个基因Si028867、Si033294-1、Si033294-2和Si028382在3个谷子品种中均扩增出单一条带;对Si027260基因扩增发现,在YG1号和JG21中扩增出单一清晰的片段,JF52中没有扩增出条带;而Si027455中均未扩增出条带。通过对3个品种的Si027260基因片段扩增结果我们推测认为,JF52未扩增出条带可能与其抗病性有关(图1)。
2.2 NBS类基因的核苷酸序列比对
基于扩增结果,我们试图分析3个品种中均扩增出条带的2个基因Si028867和Si033294在DNA序列上存在哪些碱基差异。经核苷酸序列比对后,在Si028867基因中发现有5处单核苷酸突变(图2A),依次分别是T-C、G-A、C-G、T-C、A-C,其中包含3个位置的转换和两个位置的颠换,前3个位置的碱基突变发生在中间部位,位于1 000~1 500 bp之间,剩余的2处碱基突变发生末端;比对Si033294基因碱基序列发现,在2个品种中存在1处颠换,为T-G,该突变发生在基因的起始位置,此外,在基因的260 bp处有一个单碱基A的插入(图2B),其余部分序列未发现突变。

图1 NBS类基因PCR产物电泳图谱Fig.1 Electrophoretic map of PCR product of NBS gene

图2 (2A, 2B)Si028867和Si033294两个核苷酸序列同源比对Fig.2 (2A and 2B)Homologous comparison of two nucleotide sequence in Si028867 and Si033294A.Si028867;B.Si033294
2.3 NBS类基因的的氨基酸序列分析
为明确JF52和JG21品种的2个NBS类基因Si028867和Si033294差异,对其氨基酸序列同源比对分析。发现2个品种的Si028867蛋白中有4处氨基酸发生突变,分别在338、416、704和737位氨基酸处,依次是L-S、L-V、F-L和H-P;发现 2个品种的Si033294蛋白的氨基酸差异较大,同源性较低,推测可能是由于诱变而来的JG21可能由于该基因中核苷酸片段的插入导致(图3A和3B)。
2.4 差异基因的蛋白结构分析与预测
蛋白二级结构特征分析发现,Si028867蛋白在2个品种中,无论是蛋白特性,还是二级结构特点均没有明显差异。而对Si033294的蛋白分析发现存在较大不同,其在JF52中蛋白为亲水性,其二级结构为无规则卷曲,其次为α-螺旋;在JG21中的蛋白为疏水性,二级结构则与其相反(表2)。对Si028867蛋白的三级结构预测发现,JF52和JG21中蛋白的三级结构十分类似。而Si033294蛋白的三级结构分析发现,2个谷子品种之间的差异较大(图4)。
2.5 抗性基因的系统进化分析
为了明确抗性基因的遗传进化,我们将Si028867和Si033294基因编码的氨基酸序列同水稻、高粱、小麦等禾本科植物蛋白序列比对,发现这2个蛋白与所选植物蛋白序列的同源性较高,Si028867与比对的几种植物序列的高度同源区段相对集中,而Si033294则比较分散,分布在几个区段(图5A和5B)。随后,我们分别构建了Si028867和Si033294进化发育树,谷子中Si028867蛋白与玉米的同源蛋白的亲缘关系最近,其次是水稻和高粱;而谷子中Si033294蛋白与谷子和高粱的同源蛋白聚在一起,而其它物种亲缘关系较远(图6)。
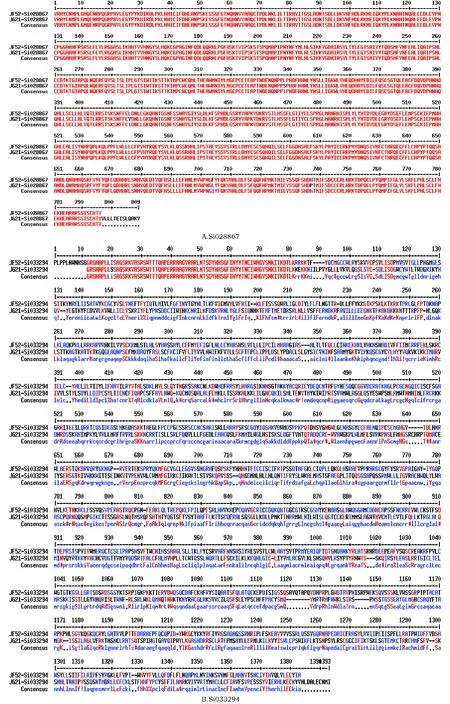
图3 (3A,3B)Si028867和Si033 294两个基因氨基酸序列同源比对Fig.3 (3A and 3B)Homologous comparison of two gene’s amino acid sequence in Si028867和Si033294A.Si028867;B.Si033294
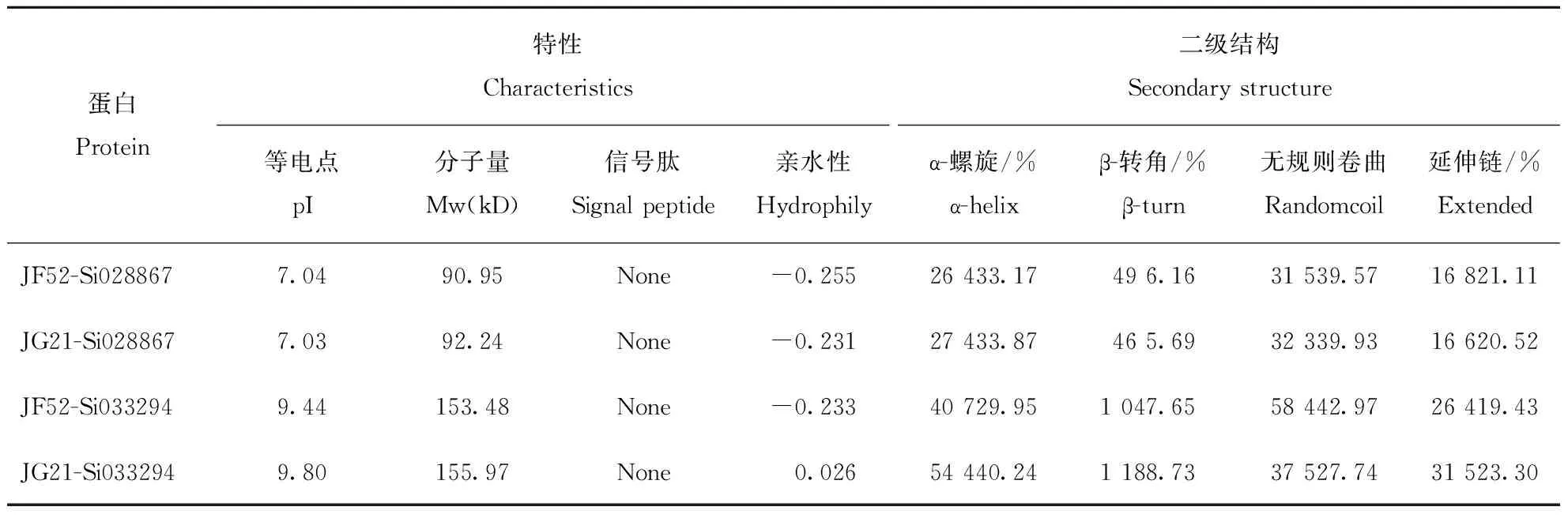
蛋白Protein特性Characteristics二级结构Secondary structure等电点pI分子量Mw(kD)信号肽Signal peptide亲水性Hydrophilyα-螺旋/%α-helixβ-转角/%β-turn无规则卷曲Randomcoil延伸链/%ExtendedJF52-Si0288677.0490.95None-0.25526 433.1749 6.1631 539.5716 821.11JG21-Si0288677.0392.24None-0.23127 433.8746 5.6932 339.9316 620.52JF52-Si0332949.44153.48None-0.23340 729.951 047.6558 442.9726 419.43JG21-Si0332949.80155.97None0.02654 440.241 188.7337 527.7431 523.30
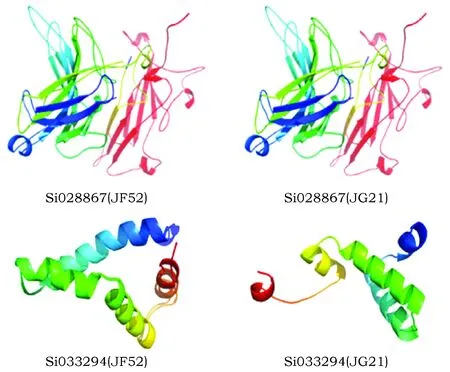
图4 Si028867和Si033294的三级结构预测Fig.4 Tertiary structure prediction of Si028867and Si033294
3 讨论与结论
植物中的NBS类型基因是抗病基因家族中数目最庞大的一类家族,抗病基因多成簇分布。抗病基因的LRR(富含亮氨酸的重复序列)位点可与病原菌相互识别。相关研究表明,LRR区域的核苷酸突变是导致抗感差异的重要因素[8~10]。Bryan 等研究发现在水稻NBS-LRR序列中单核苷酸变
异是导致水稻抗/感稻瘟病的原因所在[11]。
我们基于前期全基因组重测序数据分析获得的NBS类型抗病基因,进一步克隆比对筛选,最后发现2个基因:Si028867和Si033294。经对晋谷21、晋汾52的Si028867和Si033294序列比对发现,Si028867在LRR区域有5个核苷酸位点发生突变;对2个品种抗病基因的氨基酸序列比对发现,氨基酸在4个位置上发生了非同义突变:T-C、C-G、T-A、A-C。非同义突变很可能造成蛋白结构发生一定变化,蛋白构象变化会直接影响抗病性表现。由此推测氨基酸的非同义突变可能是造成2个亲缘关系较近的品种抗病和感病的原因。这一研究与施阳的研究结果一致[6]。对另一个基因Si033294分析,其突变也由核苷酸的插入而导致非同义突变,由于该碱基的插入导致2个氨基酸序列发生了很大差异,这些突变是否导致2个谷子品种对病原菌抗性差异的主要原因有待于进一步研究。
本研究通过对前期重测序结果分析筛选出的6个NBS类型基因通过鉴定和比较分析,晋谷21和晋汾52两个品种中发现Si028867和Si033294两个基因的突变位点较多,主要表现为非同义突变,二级结构和三级结构中也存在蛋白特征和构象变化,推测可能是2个品种间的抗病基因序列发生非同义突变,是造成抗病性差异的主要原因。本研究结果为研究谷子抗病分子机理和抗病分子辅助育种提供了理论指导。

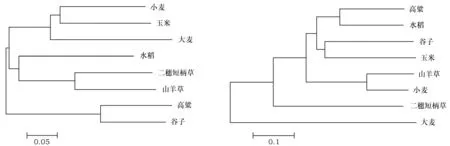
图6 (6A, 6B)Si028867和Si033294的系统发育进化树Fig.6 (6A and 6B)Phylogenetic tree of protein in millet and other plant注:左图为Si028867的系统发育进化树;右图为Si033294的系统发育进化树Note:Phylogenetic tree on the left map was Si028867; Phylogenetic tree on the right map was Si033294
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
今日农业(2022年4期)2022-06-01
当代陕西(2021年21期)2022-01-19
作物研究(2020年2期)2020-04-04
发明与创新·大科技(2019年5期)2019-07-31
中国饲料(2019年19期)2019-03-25
延河(2018年6期)2018-06-11
中学生物学(2008年3期)2008-06-03
