
豫南薄壳山核桃生长规律研究
2018-09-05 03:44章理运申明海周传涛张英姿吴文竹
广东农业科学 2018年7期
章理运,邱 林,申明海,杨 博,周传涛,张英姿,吴文竹
(1. 信阳市林业科学研究所,河南 信阳 464031;2. 信阳鸡公山自然保护区管理局,河南 信阳 464133)
薄壳山核桃(Carya illinoensis)又名长山核桃、美国山核桃、碧根果,为胡桃科(Juglandaceae)山核桃属(CaryaNutt.)落叶乔木,是世界上重要的干果树种之一,原产于美国和墨西哥北部[1]。其果仁色美味香,无涩味,营养丰富,果仁油脂中不饱和脂肪酸的含量高达 940 g/kg,优于茶油(910 g/kg)和山核桃油(890 g/kg)[2-3]。不饱和脂肪酸可以降低血液中胆固醇和甘油三酯的含量,能调节心脏功能、降低血液黏稠度、增强记忆力和思维能力,有利于人体健康,因此薄壳山核桃油脂是一种珍贵的营养保健油,是理想的保健食品,极具市场竞争力[4]。近年来,我国大力发展木本油料产业,作为油料、木材、园林绿化兼具的薄壳山核桃备受关注,浙江、江苏、云南、安徽、江西、湖北等地出现种植热潮。
薄壳山核桃在我国虽有百年的引种历史,但到目前为止仍未实现产业化生产,坚果消费基本依赖进口,造成这种现象的一个重要原因是缺乏科研和生产的积累[5],多数果树结果迟、品质差、产量低而不稳定,使得薄壳山核桃应有的生产潜力没有得到充分发挥[6]。苗木生长规律是苗木遗传特性与环境条件相适应的反映,也是进行生产管理的基础,根据苗木生长过程中不同时期的生物学特性科学管理苗木,可以避免管理上的盲目性,降低管理成本,同时可以人为地施加“促生”和“抑生”条件,如施肥、剪枝、摘心、环剥、环割等调控措施来抑制营养生长,促进生殖生长[7-8],达到早产丰产的目的。目前,国内对薄壳山核桃的研究主要集中在繁殖技术、开花与授粉能力观察[9-10]、光合作用[11-12]、果实生长和品质特征[13-14]以及人为调控对其生长的影响[15-16]等方面,而在生长规律方面的研究较少,也缺少生长时期划分的有关资料,不利于对该树种的持续动态管理。本研究以种植2年后的波尼薄壳山核桃苗木为试验材料,通过年生长周期内连续不间断的监测记录,采用曲线模型对其年生长变化情况进行模拟分析,同时对生长时期进行划分,以探索其生长规律,旨在为薄壳山核桃在豫南地区的丰产栽培与科学管理等提供理论和实践依据。
1 材料与方法
1.1 试验材料
供试材料为波尼薄壳山核桃嫁接苗,2016年3月从浙江金华引进700余株,苗高70~120 cm。
试验地设在信阳市浉河区游河乡和罗山县周党镇,为北亚热带和暖温带的南北过渡区,属北亚热带季风类型,雨热同季,气候温和,四季分明,年平均气温15.1~15.3℃,最冷月平均气温10℃,最热月平均气温29.9℃,极端最高气温40.9℃,极端最低气温-20.3℃,无霜期220~230 d,年平均降水量1 100 mm左右,≥10℃的年积温4 820~4 970℃。属低山丘陵区,海拔100~200 m,地势稍有起伏,土壤为黄棕壤,土层厚度一般在50 cm以上,pH值为6.5左右,呈微酸性。
1.2 试验方法
试验于2015年10月底开始整地挖穴,树穴规格80 cm×80 cm×70 cm,造林密度设6 m×6 m、6 m×8 m、8 m×8 m 3种 规 格。2016年3月将薄壳山核桃嫁接苗按3种种植密度定植于试验地中,经1年栽植缓苗后,2017年春季开始观测。
自2017年3月15日芽萌动至11月30日树木开始落叶,选择30株生长发育正常的薄壳山核桃树苗为标准株并挂牌,每隔15 d测定1次,主要测定株高、新梢长、冠幅(钢卷尺测量,精确到0.1 cm)和地径(游标卡尺测量,精确到 0.01 mm)。
1.3 数据处理
1.3.1 生长时期的划分 采用有序样本聚类法(最优分割法)[17],借助 DPS2000有序样本聚类分析程序计算。
1.3.2 生长过程的拟合 应用SPSS 17软件进行回归分析[18-19],采用的数学模型包括二次曲线、等比级数曲线、S形曲线、三次方程、指数方程、幂函数曲线、Logistic方程模型。
二次曲线:Y=A+BT+CT2
等比级数曲线 :Y=e(A+BT)
S 移曲线 :Y=e(A+B/T)
三次方程:Y=A+BT+CT2+DT3
指数方程 :Y=Ae(BT)
幂函数曲线:Y=ATB
Logistic 方程 :Y=1/(1/u+ABT)
式中,Y为生长量,A、B、C、D为随机参数。分别将薄壳山核桃的苗高、地径、分枝(新稍)生长数据用上述7个模型进行拟合,通过回归分析,从中选择最优模型。
2 结果与分析
2.1 河南薄壳山核桃株高年生长规律
薄壳山核桃栽植的第1年,由于起苗时根系损伤、生长量较少,处于保活和树体恢复阶段,第2年开始正常生长。在豫南地区,山核桃于3月15日左右开始萌动,4月上旬开始抽稍、展叶,4月中、下旬新稍迅速伸长,生长较快,形成第1次生长高峰期,6月下旬至7月上旬有一个减慢生长阶段,持续时间在15~20 d;7月中旬至8月下旬生长加快,形成第2次生长高峰期,9月上旬生长量减慢,9月15日以后株高生长基本停止,但叶片在树上一直保持至10月底,11月上旬叶片开始变色,11月中旬逐渐脱落。全年株高净生长量为77.6 cm(平均值),呈现出明显的“快-慢-快-慢”的生长节律。
2.1.1 株高年生长时期划分 薄壳山核桃的苗高生长随着时间的推移出现动态变化规律,对其生长时期的划分需按顺序把样本分割成若干段,因此本研究采用有序样本聚类分割法(Fisher分类方法)对薄壳山核桃的生长时期进行划分,从3月下半月开始至11月上半月止,每15 d净生长量的数据按顺序连续编号为x1,x2,x3,…x16。通过计算机软件程序可将这16个有序样本分为4类:{ x1},{ x2,x3,x4,x5,x6,x7,x8},{ x9,x10,x11},{ x12,x13,x14,x15,x16}。将分类结果与实际测量结果相结合,可将薄壳山核桃的生长时期划分为4个时期(表1)。从表1可以看出,薄壳山核桃在豫南的年株高生长时间达180 d,年株高总生长量为77.6 cm,每15 d的净生长量见图1。
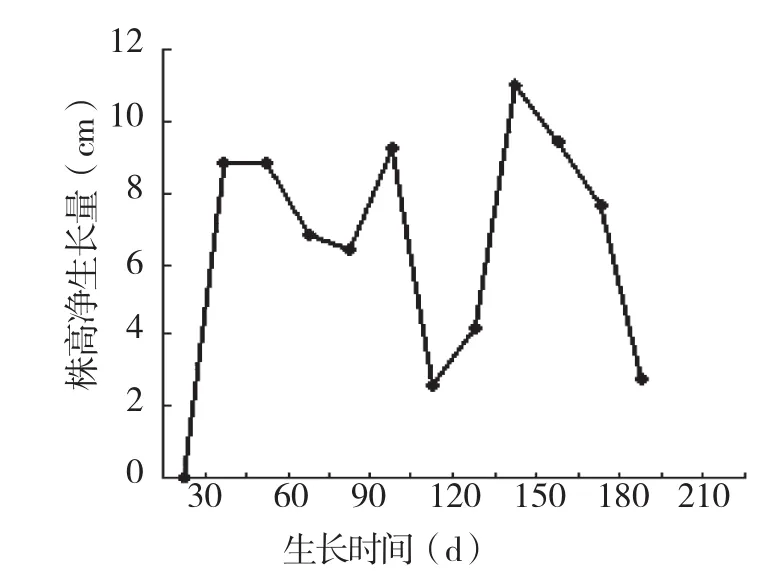
图1 河南薄壳山核桃株高净生长量变化
2.1.2 株高年生长模型 由表2可知,用三次方程曲线模型拟合的决定系数(R2=0.991)最大,拟合效果最好,因此薄壳山核桃造林第2年株高的生长模型为:
H=82.145+4.923T+0.515T2-0.032T3
经差异显著性检验,模型的方差分析结果达到极显著水平(F=465.876,P<0.01,表3),表明回归结果较可靠,推测的实际生长过程具有较高的准确性(图2)。
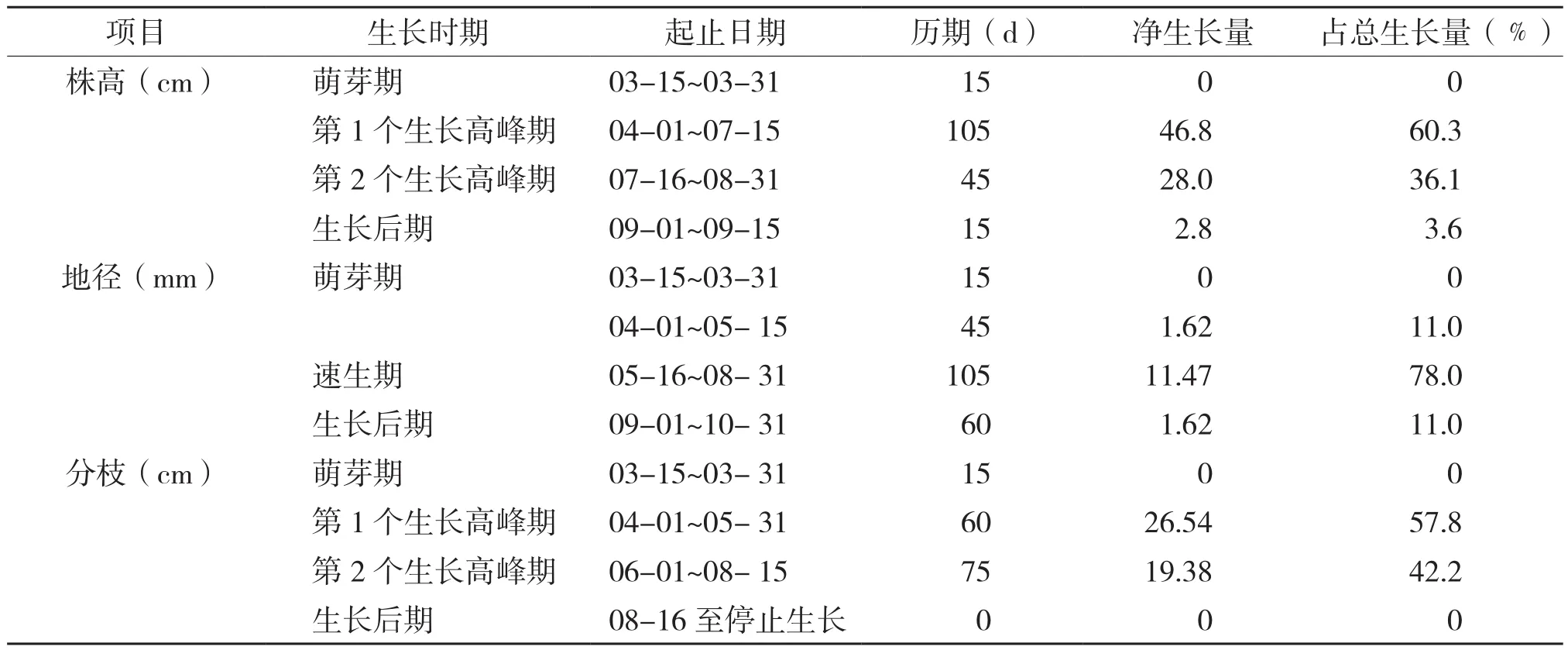
表1 河南薄壳山核桃株高、地径、分枝各生长时期的划分及生长情况
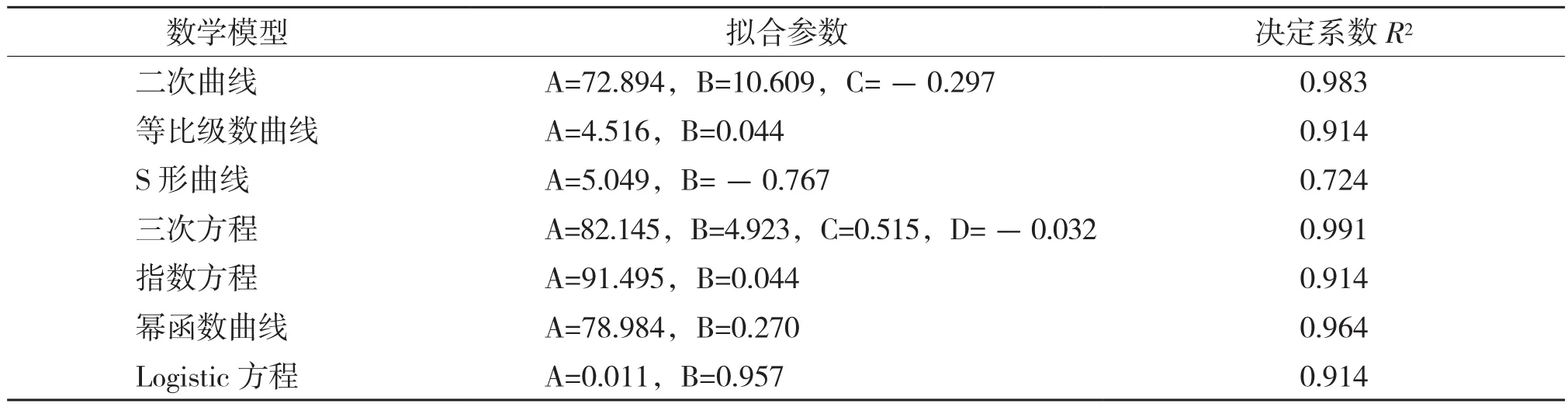
表2 河南薄壳山核桃树高连续生长数学模型拟合结果
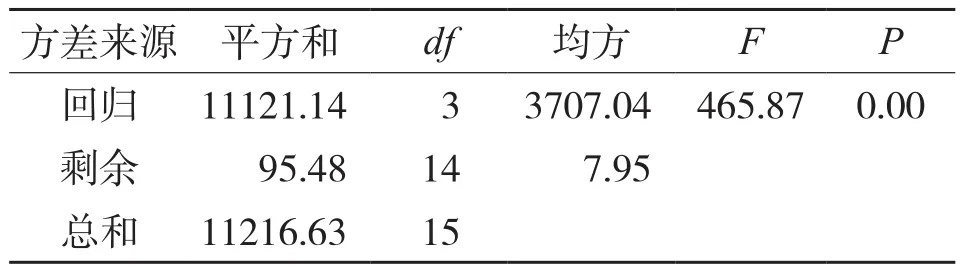
表3 河南薄壳山核桃株高连续生长回归方差分析
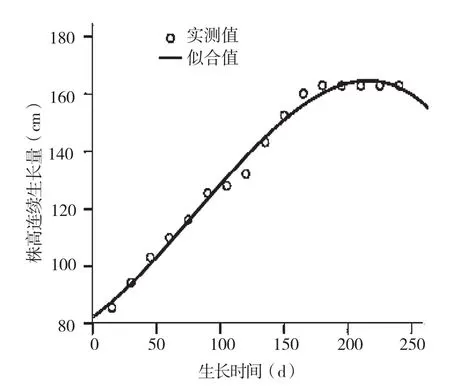
图2 河南薄壳山核桃株高连续生长量及三次曲线拟合结果
2.2 河南薄壳山核桃地径年生长规律
薄壳山核桃地径的生长与株高生长相关,随着苗高的增长地径也相应地增长。地径生长进入速生期的时间较株高生长进入高峰期的时间延后50~60 d,生长期也相应较株高生长延后约45 d,说明株高生长停止后地径仍会维持较长一段时间的生长过程。用上述7个模型对薄壳山核桃地径连续生长量进行模拟,结果见表4,表明薄壳山核桃地径的连续生长模型为:
D=18.416-0.957T+0.324T2-0.013T3
地径模型的方差分析结果见表5,从表5可以看出,回归结果较为理想,拟合模型的方差分析结果达到极显著水平(F=910.332,P<0.01),模拟拟合曲线见图3,表明地径生长与生长时间存在极显著相关关系。
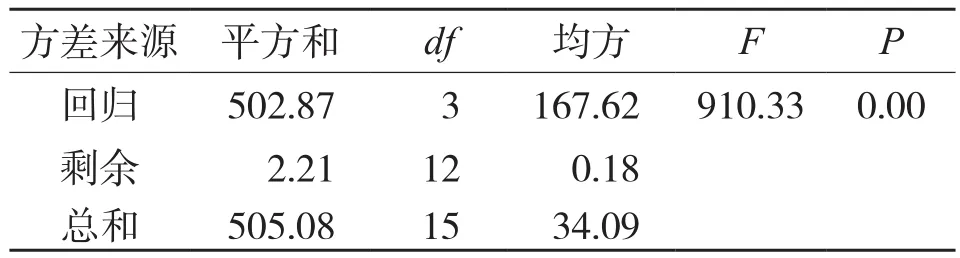
表5 河南薄壳山核桃地径连续生长回归方差分析

表4 河南薄壳山核桃地径连续生长数学模型拟合结果
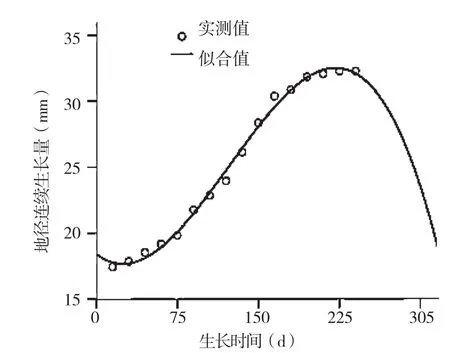
图3 河南薄壳山核桃地径连续生长量及三次曲线拟合结果
利用DPS软件对观测的16组地径数据进行有序样本的聚类分析结果,可将地径生长分为萌芽期、生长初期、速生期、生长后期4个阶段,各个生长时期的生长量以及占全年地径的生长比例见表1。地径的生长情况与株高生长不同,地径生长表现出“慢-快-慢”的生长过程,速生期集中在5月中旬至8月底,年生长时间达225 d,年净生长量为14.71 mm(平均值)。由图4可知,地径生长在开始生长的4月至5月上旬生长较慢,时间约45 d左右,然后进入快速生长阶段,在整个快速生长阶段与株高生长不同,生长减慢阶段生长量降低的程度较株高生长小。
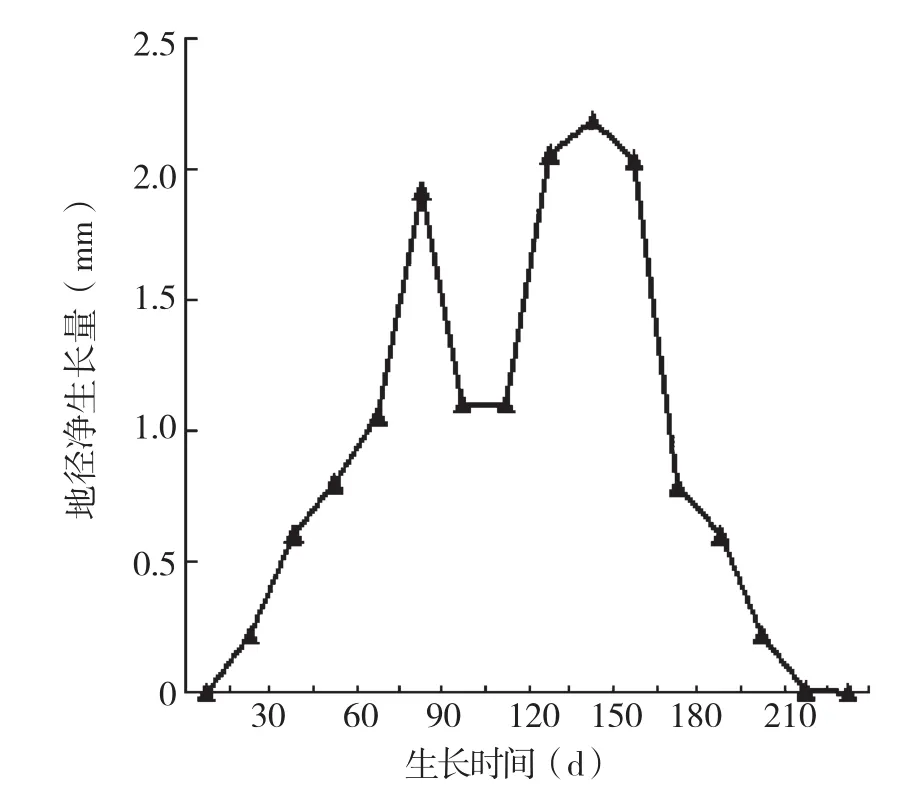
图4 河南薄壳山核桃地径净生长量变化
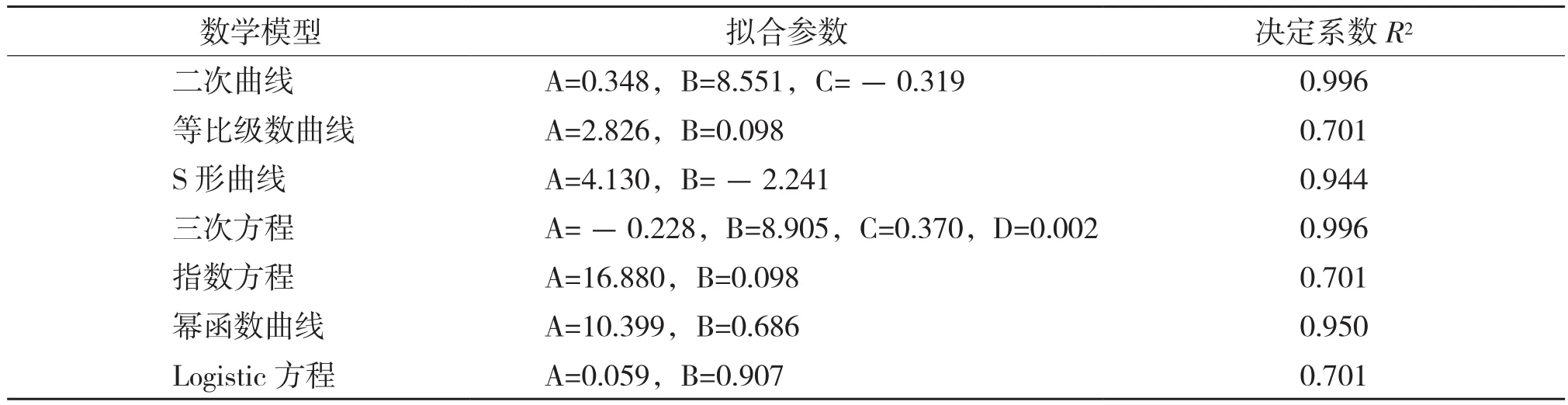
表6 河南薄壳山核桃分枝连续生长数学模型拟合结果
2.3 河南薄壳山核桃分枝(新梢)年生长规律
薄壳山核桃分枝的年生长规律与株高生长相似,但与年高生长节律又有所不同,分枝的第1个生长高峰期在5月31日前,6月上中旬生长减慢,6月下旬至7月上旬生长加快,7月下旬达到第2个生长高峰,分枝生长表现出“快-慢-快”的生长节律。分枝的生长停止时间与株高、地径不同,较株高生长停止时间提前1个月,较地径提前2个月。用上述7个曲线模型进行模拟,结果见表6。从表6可以看出,二次曲线、S形曲线、三次方程和幂函数曲线等4个模型均能较好地拟合,为了简单方便,本研究选择二次曲线模型为薄壳山核桃分枝的连续生长模型:
Y=0.348+8.551T-0.319T2
方差分析结果(表7)表明,拟合结果达到极显著水平(F=1626.872,P<0.01),分枝累积生长量及二次曲线拟合结果见图5,可见用此回归模型描述分枝的生长规律是合适的。

表7 河南薄壳山核桃分枝连续生长回归方差分析

图5 河南薄壳山核桃分枝连续生长量及二次曲线拟合结果
用有序样本聚类分割法对薄壳山核桃的分枝生长时期进行划分,可以将薄壳山核桃的分枝生长分为4个时期:萌芽期、第1个生长高峰期、第2个生长高峰期、生长后期。由图6可知,分枝的生长与株高生长同时进行,分枝的年生长时间为150 d、年净生长量为45.92 cm(平均值)。生长后期虽然枝条上有叶片,但较少伸长,8月15日后分枝生长基本停止(表1)。
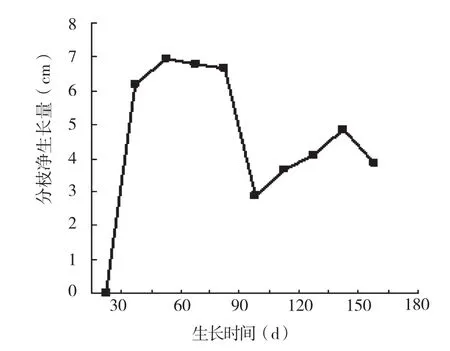
图6 河南薄壳山核桃分枝净生长量变化
3 结论与讨论
本研究结果表明,在豫南地区,薄壳山核桃幼龄林阶段,其株高、地径的生长规律遵循三次方程曲线,分枝(新梢)的生长规律遵循二次曲线,利用这3个方程可以模拟株高、地径和分枝(新梢)的生长过程,拟合效果较好。与陈炜青等[20]对油橄榄(Olea europaeaL.)、张琳等[7]对银雀树(Tapiseia siriensisOliv.)、蒋燚等[21]对黑荆树(Acacia mearnsii)的研究结果不同,他们用Logistic方程拟合树高、地径的效果较好,而本试验通过7个曲线模型拟合结果表明,薄壳山核桃的株高、地径生长规律更适合三次方程曲线;与周祥斌等[22]对香椿(Toona sinensis)的研究相比,株高的拟合结果不同,而地径的拟合结果基本相同。说明薄壳山核桃的高生长规律与这4个树种的高生长规律存在差异,而地径的生长规律与香椿的地径生长规律相似,原因可能是由于树种本身生物学特性的差异引起,或者是由于树龄、地理气候因素引起。从分枝的拟合结果看,生长规律既符合二次曲线,亦符合三次方程,而用Logistic方程拟合效果不理想,说明用Logistic方程模拟树木生长规律有局限性。随着树龄的增长,或人为施加管理等因素的影响是否仍遵循这一规律需进一步研究。
通过观测发现,波尼薄壳山核桃在豫南的年生长时间可达220 d以上,达到薄壳山核桃生长发育对时间要求。其中,年株高净生长量为77.6 cm,年地径净生长量为14. 71 mm,与吕运舟等[23]的研究结果以及美国原产地生长表现相似。用Fisher分类方法对波尼株高、地径、分枝的生长时期进行划分,结果表明三者的生长时期均不相同,株高和分枝的生长均有2个生长高峰期,但时间不同;株高的第1个生长高峰期从4月1日至7月15日(105 d),生长量占全年的60.3%;而分枝的第1个生长高峰期为4月1日至5月30日(60 d),生长量占全年的57.8%,较株高提前45 d结束;株高的第2个生长高峰期从7月15日至9月15日,而分枝生长从6月1日至8月15日,较株高提前45 d进入高峰期,提前30 d结束。这与熊新武等[24]的研究结果不尽相同,其中,2次生长高峰相同,但株高生长高峰期的时间不同,生长量占比也不同;分枝高峰期的时间基本相同,生长量占比不同。也与殷巧等[25]、姚小华等[26]研究的高峰期时间结果不同,原因可能是由于品种或树龄或地理位置不同等引起,有待进一步研究。
本研究结果表明,株高、地径、分枝(新梢)生长规律的3个曲线方程可以为薄壳山核桃在豫南地区幼龄林时期的生长动态预测与科学管理提供依据,对生长阶段的划分亦可为造林后实际生产阶段的管理提供指导,按照不同阶段的生长特性采取相应的管理措施。生长高峰期对树木的生长起着关键作用,因此,要加强水、肥管理,促进苗木的快速生长及根系发达,而在生长减慢期如5月下旬至6月下旬可进行修剪、拉枝、扭伤等管理,促进苗木提前结果。
猜你喜欢
农家参谋(2021年10期)2021-12-05
河北果树(2021年4期)2021-12-02
广东蚕业(2021年9期)2021-12-01
数学大王·中高年级(2021年3期)2021-03-10
中国食品(2020年21期)2020-12-07
今日农业(2019年13期)2019-01-03
小学生作文(中高年级适用)(2018年4期)2018-05-14
阅读(中年级)(2018年11期)2018-01-19
学生天地·小学中高年级(2015年11期)2016-01-14
环球时报(2009-09-11)2009-09-11
