
高寒沙区不同生境赖草的生长特征分析
2018-08-30 07:18:04张立恒贾志清王学全李清雪陈新均李少华
植物资源与环境学报 2018年3期
张立恒, 贾志清, 王学全,①, 李清雪, 陈新均, 李少华
(1. 中国林业科学研究院荒漠化研究所, 北京 100091; 2. 甘肃省治沙研究所, 甘肃 兰州 730070)
赖草〔Leymussecalinus(Georgi) Tzvel.〕隶属于禾本科(Poaceae)赖草属(LeymusHochst.),为多年生草本植物,在中国主要分布于新疆、青海、甘肃、宁夏、陕西、内蒙古、山西和河北等省(自治区)以及东北地区[1]。赖草是一种长根茎型植物,具有下伸和横走的根茎,秆通常单生或丛生且直立。赖草属于典型的无性系克隆植物,主要利用营养器官或营养体的一部分生长发育为新个体,其根茎繁殖迅速、耐寒耐旱,能在不同的环境条件下繁殖和更新[2-3]。赖草对不同生境的适应性均较强,在干旱和半干旱地区的草原和荒漠中均可生长,因其特殊的克隆整合性,在干旱地区大量生存扩张,常成为单优群落[4-5]。赖草根茎具有很强的可塑性,能够根据不同生境土壤水分和养分的空间异质性调整其根系的形态和分布范围,这种根系形态的变化和分布特征充分反映了赖草对恶劣环境的适应策略。
赖草为固定沙丘和丘间地的优良固沙植物之一,对干旱荒漠区的防风固沙和植被恢复具有重要作用[6]。在冬季和春季,赖草干枯枝挺立在沙丘表面,增大了地表粗糙度,削弱了地表风沙流作用,减轻了地表的风蚀程度,进而维持沙丘表面稳定;在农田田边或退耕荒地,密集的赖草根系交织分布在土壤中,对干裂地块起到缝合作用,防止土壤遭到侵蚀,在中国的防沙治沙和生态建设中发挥着其他草本植物不可替代的生态功能。近些年来,国内许多研究人员对平原、草地和沙地上生长的赖草种群进行了无性系繁殖、呼吸动态、构件结构、分株特征、生产性能及营养价值等方面的研究[7-10],但是对高寒沙区赖草的研究却鲜有报道。
本研究以青海省治沙试验站周边4种不同生境中的赖草为研究对象,对4种生境土壤的含水量、有机质和养分含量以及赖草的生长特征进行了初步研究,以期为当地的植被管护和生态修复重建提供参考,并为西部荒漠地区的防风固沙和禾草类植物抗逆新品种的选育与开发提供帮助。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于青海省治沙试验站周边,地处青藏高原东北缘,海拔2 871~3 870 m;年均气温约2.4 ℃;年均降水量311.1~402.1 mm,且降水主要集中在7月和8月,约占全年降水量的50%;年均蒸发量高达1 500~1 900 mm,属于典型的高寒干旱和半干旱区[11]。该地常年风沙活动频繁且多集中在冬季和春季,风向主要为西风和西北风,年均风速2.7 m·s-1,年均大风日数50.6 d,年均沙尘暴日数多达20.7 d。研究区内地带性土壤包括棕钙土和栗钙土,非地带性土壤包括草甸土和风沙土。研究区内的植被以人工植被为主,植物种类包括青杨(PopuluscathayanaRehd.)、乌柳(SalixcheilophilaSchneid.)、沙棘(HippophaerhamnoidesLinn.)、柠条锦鸡儿(CaraganakorshinskiiKom.)、柽柳(TamarixchinensisLour.)、小穗柳(SalixmicrostachyaTurcz. ex Trautv.)、白刺(NitrariatangutorumBobr.)、赖草、沙蓬〔Agriophyllumsquarrosum(Linn.) Moq.〕和黑沙蒿(ArtemisiaordosicaKrasch.)等[12]。
1.2 研究方法
1.2.1 样地设置和样品采集 于2017年7月下旬,在青海省治沙试验站周边分别选取低湿滩地、农田田边、丘间地和固定沙丘4种生境设置样地,4种生境样地的基本情况见表1。样地面积均为10 m×10 m,在每个样地内随机设置5个面积1 m×1 m的小样方,视为5个重复。
表1高寒沙区不同生境样地的基本情况
Table1Basicsituationofdifferenthabitatplotsinalpinesandyarea
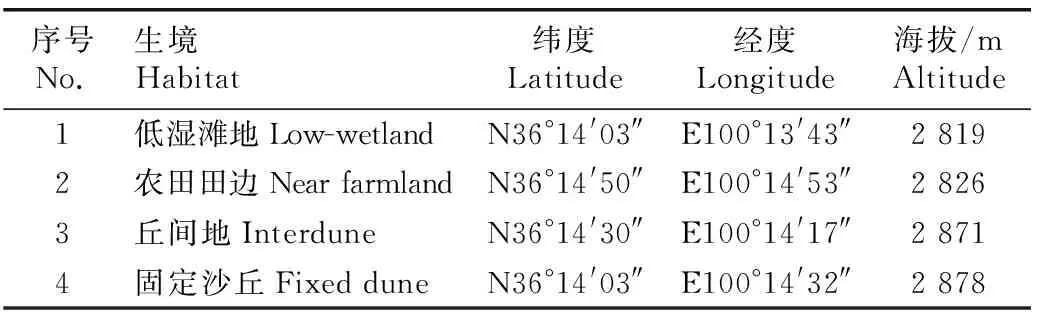
序号No.生境Habitat纬度Latitude经度Longitude海拔/mAltitude1低湿滩地Low-wetlandN36°14'03″E100°13'43″2 8192农田田边Near farmlandN36°14'50″E100°14'53″2 8263丘间地InterduneN36°14'30″E100°14'17″2 8714固定沙丘Fixed duneN36°14'03″E100°14'32″2 878
在每个小样方内选择长势良好的赖草10株,使用钢卷尺(精度0.1 cm)测量其株高,使用游标卡尺(精度0.01 mm)测量其地径,并统计其分株密度(1 m2内赖草的分株数),计算各指标的平均值。然后将各样方内赖草的地上部分全部收割带回实验室称量其鲜质量;同时以10 cm为一层由地表向下逐层挖掘直至看不到赖草根系,分别拣出每层中的根系,装入密封袋后带回实验室测定相关指标,并逐层采集土样带回实验室测定土壤含水量以及有机质和养分(全氮、全磷和全钾)含量。
1.2.2 样品处理和相关指标测定 将取回的地上部分和每层根系分别用清水冲洗干净,再用去离子水冲洗3次后吸水纸吸干水分,然后于105 ℃杀青15 min后再于70 ℃烘干至恒质量,使用天平(精度0.01 g)称量后计算生物量。选择代表性根茎,使用游标卡尺(精度0.01 mm)测量根茎直径和间隔子长等,计算各指标的平均值。
取回的土壤样品先称量其鲜质量,然后于105 ℃烘干8~10 h至恒质量,称量后计算土壤含水量。土壤有机质含量采用重铬酸钾-外加热法[13]30进行测定;经H2SO4-H2O2消煮后,土壤中全氮含量采用凯氏定氮法[13]42进行测定,全磷含量采用钼锑抗比色法[13]76进行测定,全钾含量采用火焰光度法[13]101进行测定,计算各指标的平均值。
1.3 数据处理和分析
采用EXCEL 2007和IBM SPSS Statistics 19.0统计分析软件进行数据处理和图表绘制,采用LSD法进行差异显著性分析。
2 结果和分析
2.1 高寒沙区不同生境不同土层土壤含水量以及有机质和养分含量的比较
2.1.1 土壤含水量的比较 高寒沙区不同生境不同土层的土壤含水量见表2。由表2可以看出:在0~30 cm土层范围内,低湿滩地各土层的土壤含水量均最高,农田田边各土层的土壤含水量次之,丘间地各土层的土壤含水量较低,固定沙丘各土层的土壤含水量最低;在30~40 cm土层范围内,农田田边的土壤含水量最高,低湿滩地的土壤含水量次之,丘间地的土壤含水量较低,固定沙丘的土壤含水量最低;而在40~60 cm土层范围内,农田田边各土层的土壤含水量均最高,低湿滩地各土层的土壤含水量次之,固定沙丘各土层的土壤含水量较低,丘间地各土层的土壤含水量最低。总体上看,低湿滩地和农田田边各土层的土壤含水量显著高于丘间地和固定沙丘(P<0.05)。


生境Habitat不同土层的土壤含水量/% Soil moisture content in different soil layers0-10 cm10-20 cm20-30 cm30-40 cm40-50 cm50-60 cm低湿滩地Low-wetland7.69±0.42Ac14.08±0.38Aa10.25±0.27Ab7.31±0.16Ac7.21±0.12Ac7.38±0.24Ac农田田边Near farmland6.19±0.30Bb6.36±0.25Bb7.65±0.07Ba7.52±0.19Aa7.38±0.05Aa7.69±0.18Aa丘间地Interdune0.80±0.03Cd1.52±0.02Cc1.60±0.01Cc2.44±0.01Bb2.92±0.00Cab3.11±0.03Ca固定沙丘Fixed dune0.34±0.02Cd0.86±0.01Dc1.20±0.02Cbc1.30±0.00Cb3.87±0.02Ba3.99±0.04Ba
1)同列中不同的大写字母表示不同生境间差异显著(P<0.05) Different capitals in the same column indicate the significant difference among different habitats (P<0.05); 同行中不同的小写字母表示不同土层间差异显著(P<0.05) Different lowercases in the same row indicate the significant difference among different soil layers (P<0.05).
由表2还可以看出:随着土层的加深,低湿滩地的土壤含水量基本呈先升高后降低的变化趋势,其中,10~20 cm土层的土壤含水量高达14.08%,显著高于其他土层的土壤含水量,该生境各土层土壤含水量的平均值为8.99%;农田田边、丘间地和固定沙丘的土壤含水量总体上呈升高的变化趋势,3种生境各土层土壤含水量的平均值分别为7.13%、2.07%和1.93%。
2.1.2 土壤有机质和养分含量的比较 高寒沙区不同生境不同土层土壤有机质和养分(全氮、全磷和全钾)的含量见图1。由图1可以看出:总体上看,在0~20 cm土层范围内,低湿滩地各土层土壤有机质、全氮、全磷和全钾的含量最高,显著高于其他3种生境;农田田边各土层土壤有机质和各养分含量次之;丘间地各土层土壤有机质和各养分含量较低;固定沙丘各土层土壤有机质和各养分含量最低。在20~40 cm土层范围内,低湿滩地和农田田边各土层土壤有机质和各养分含量较高,丘间地和固定沙丘各土层土壤有机质和各养分含量较低。
由图1还可以看出:随着土层的加深,低湿滩地土壤有机质和各养分含量呈逐渐降低的变化趋势,0~20 cm土层土壤的有机质和各养分含量总体上显著高于20~40土层土壤;农田田边和丘间地土壤有机质和各养分含量总体上呈波动下降的变化趋势;固定沙丘土壤有机质和各养分含量的变幅较小,且不同土层间总体上差异不显著。
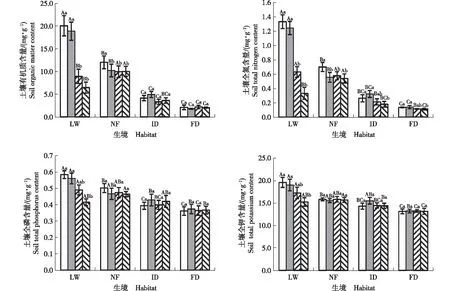
: 0~10 cm土层Soil layer of 0-10 cm; : 10~20 cm土层Soil layer of 10-20 cm; : 20~30 cm土层Soil layer of 20-30 cm; : 30~40 cm土层Soil layer of 30-40 cm. LW: 低湿滩地Low-wetland; NF: 农田田边Near farmland; ID: 丘间地Interdune; FD: 固定沙丘Fixed dune.不同的大写字母表示同一土层不同生境间差异显著(P<0.05) Different capitals indicate the significant difference among different habitats in the same soil layer (P<0.05); 不同的小写字母表示同一生境不同土层间差异显著(P<0.05) Different lowercases indicate the significant difference among different soil layers of the same habitat (P<0.05).图1 高寒沙区不同生境不同土层土壤有机质和养分含量的比较Fig. 1 Comparison on soil organic matter and nutrient contents in different soil layers of different habitats in alpine sandy area
2.2 高寒沙区不同生境中赖草地上部分生长特征的比较
高寒沙区不同生境中赖草地上部分的生长特征见表3。由表3可以看出:固定沙丘中赖草的株高和地径最大,分别为76.20 cm和2.57 mm,显著大于其他3种生境(P<0.05);农田田边中赖草的株高和地径次之,分别为72.56 cm和2.17 mm;丘间地中赖草的株高和地径较小,分别为69.38 cm和1.96 mm;低湿滩地中赖草的株高和地径最小,分别为58.37 cm和1.61 mm。
由表3还可以看出:低湿滩地中赖草的分株密度最大(150.3 m-2),显著大于其他3种生境;农田田边中赖草的分株密度较大(129.9 m-2),显著大于丘间地和固定沙丘;丘间地和固定沙丘中赖草的分株密度较小,分别为31.6和34.6 m-2。
由表3还可以看出:农田田边中赖草的地上部分生物量最大(208.74 g·m-2),显著大于其他3种生境;低湿滩地和固定沙丘中赖草的地上部分生物量居中,分别为120.01和107.36 g·m-2;丘间地中赖草的地上部分生物量最小(82.64 g·m-2),显著小于其他3种生境。
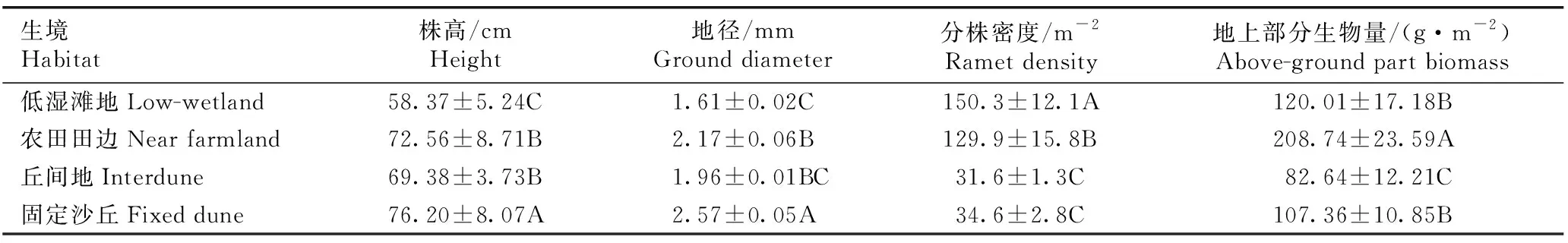
生境Habitat株高/cmHeight地径/mmGround diameter分株密度/m-2Ramet density地上部分生物量/(g·m-2)Above-ground part biomass低湿滩地Low-wetland58.37±5.24C1.61±0.02C150.3±12.1A120.01±17.18B农田田边Near farmland72.56±8.71B2.17±0.06B129.9±15.8B208.74±23.59A丘间地Interdune69.38±3.73B1.96±0.01BC31.6±1.3C82.64±12.21C固定沙丘Fixed dune76.20±8.07A2.57±0.05A34.6±2.8C107.36±10.85B
1)同列中不同的大写字母表示不同生境间差异显著(P<0.05) Different capitals in the same column indicate the significant difference among different habitats (P<0.05).
2.3 高寒沙区不同生境中赖草根系生长特征的比较
高寒沙区不同生境中赖草根系的生长特征见表4。由表4可以看出:低湿滩地、农田田边、丘间地和固定沙丘中赖草的根系分布深度分别为0~30、0~40、0~40和0~60 cm。低湿滩地中赖草的根茎直径最大(2.06 mm),显著大于其他3种生境(P<0.05);农田田边中赖草的根茎直径次之,为1.82 mm;丘间地和固定沙丘中赖草的根茎直径较小,分别为1.66和1.63 mm。固定沙丘中赖草的间隔子长最长(10.98 cm),显著大于其他3种生境;丘间地中赖草的间隔子长较长(9.06 cm),显著大于低湿滩地和农田田边;低湿滩地和农田田边中赖草的间隔子长较短,分别为7.69和7.52 cm。总体上看,从低湿滩地、农田田边、丘间地到固定沙丘,赖草的根系分布深度和间隔子长增加,根茎直径减小。
由表4还可以看出:农田田边中赖草的根系生物量最大(234.54 g·m-2),显著大于其他3种生境;低湿滩地中赖草的根系生物量较大(169.61 g·m-2),显著大于丘间地和固定沙丘;丘间地和固定沙丘中赖草的根系生物量较小,分别为110.44和123.10 g·m-2。

生境Habitat根系分布深度/cmRoot distribution depth根茎直径/mmRhizome diameter间隔子长/cmSpacer length根系生物量/(g·m-2)Root biomass低湿滩地Low-wetland0-302.06±0.38A7.69±0.21C169.61±20.36B农田田边Near farmland0-401.82±0.06B7.52±0.58C234.54±23.59A丘间地Interdune0-401.66±0.15BC9.06±0.83B110.44±15.25C固定沙丘Fixed dune0-601.63±0.12C10.98±1.05A123.10±6.46C
1)同列中不同的大写字母表示不同生境间差异显著(P<0.05) Different capitals in the same column indicate the significant difference among different habitats (P<0.05).
对高寒沙区不同生境不同土层中赖草根系生物量的分布特征进行进一步分析,结果见图2。由图2可以看出:在高寒沙区低湿滩地、农田田边、丘间地和固定沙丘4种生境中,赖草根系主要分布在浅层土壤。随着土层的加深,4种生境中赖草根系生物量均呈先增加后减少的变化趋势,且10~20 cm土层的根系生物量均最大,显著大于其他土层。低湿滩地和农田田边中赖草根系生物量集中分布在0~30 cm土层范围,而丘间地和固定沙丘中赖草根系生物量集中分布在10~30 cm土层范围。

LW: 低湿滩地Low-wetland; NF: 农田田边Near farmland; ID: 丘间地Interdune; FD: 固定沙丘Fixed dune.不同的小写字母表示不同土层间差异显著(P<0.05) Different lowercases indicate the significant difference among different soil layers (P<0.05).图2 高寒沙区不同生境不同土层中赖草根系生物量分布特征Fig. 2 Distribution characteristics of root biomass of Leymus secalinus (Georgi) Tzvel. in different soil layers of different habitats in alpine sandy area
3 讨论和结论
生境是生物的个体、种群或群落生活地域的环境,包括必需的生存条件和其他对生物起作用的生态因子[14],同一物种在不同生境中的形态和分布特征有所不同,表现出不同的表型。表型可塑性是生物体通过单个基因型产生多变的形态特征和生理特性以适应复杂环境的能力[15],是植物器官在复杂环境中产生一系列不同的相对适合的表型的潜能,是个体适应环境的方式[16],在物种的生长和进化过程中起至关重要的作用。赖草是一种长根茎型植物,根系拥有很强的表型可塑性,因生境土壤水分和养分的不同表现出不同的形态变化特征。本研究中,从低湿滩地、农田田边、丘间地到固定沙丘,土壤含水量以及有机质、全氮、全磷和全钾含量总体上呈逐渐降低的变化趋势,各生境中赖草的根系表现出明显的表型可塑性,如赖草的间隔子长逐渐增加,而分株密度逐渐减小。已有研究结果显示:当克隆植物处于资源条件好的斑块中时,其根茎节长和间隔子长通常会缩短,根系构型趋于“密集型”[17]。低湿滩地由于地势低洼,夏季暴雨后常有积水,且土层多为黏土,土壤含水量以及有机质和养分含量较高,该生境中生长的赖草根系能在基株周围获得满足生长的营养资源,根茎无需延长间隔子长和根茎长去更远更深的空间里吸取生长所需的水分和营养,而是在基株周围或较近的范围大量克隆繁殖,从而有更多分株可能被放置到资源条件较好的斑块中,有利于整个植株对资源的获取。密集分布的赖草为获得生长资源种内竞争相对激烈,最终导致植株高度相对矮小,茎秆相对较细。农田田边常年受农业灌溉和施肥的影响,土壤的水分和肥力条件也相对较好,赖草在该生境中的分株密度也相对较大,但是赖草周围伴有碱菀(TripoliumvulgareNees)、冰草〔Agropyroncristatum(Linn.) Gaertn.〕、芨芨草〔Achnatherumsplendens(Trin.) Nevski〕和甘草(GlycyrrhizauralensisFisch.)等田间草本植物,为占领更广阔的生态位以及获取生长所需的营养和光照资源,赖草与其他草本植物相互竞争,其分株密度略低于低湿滩地,但株高和地径略大于低湿滩地。从低湿滩地、农田田边、丘间地到固定沙丘,土壤渐趋干旱和贫瘠,赖草的根茎直径逐渐减小,由2.06 mm减小至1.63 mm,这与洪光宇等[18]的研究结果一致,说明随着生境条件变差,根茎型植物的根系主要表现为根茎直径的下降及其长度的增长,即根茎型植物主动牺牲根系增粗生长,而尽可能地向更远更深的空间延伸,以获取满足生长所需的资源。
丘间地和固定沙丘土壤表层植被稀少、蒸发强烈、风蚀严重,土壤含水量较低,土壤中有机质和养分流失严重,尤其是固定沙丘,土壤表层极其干旱,有机质和养分也极其匮乏。在资源条件差的斑块中,克隆植物的根茎节长、间隔子长和根茎长增加,根系构型趋于“游击型”,有利于基株向资源丰富的斑块伸展去获取资源[17]。在干旱和资源贫乏的丘间地和固定沙丘中,赖草根系无法在基株附近获取满足生长所需的资源,根茎通过增大间隔子长和根茎长尽力延伸到更大的范围去获取资源。赖草通过根茎延伸分株的定植范围,尽力将分株放置到不同资源斑块中实现资源共享[19],以便于根系穿过资源有效性低的生境斑块,向资源有效性高的生境斑块分布以吸取营养来提高种群个体的存活概率。任安芝等[20]的研究结果表明:干旱胁迫导致赖草分株数目明显降低,且总生物量也显著下降。本研究中,从低湿滩地、农田田边、丘间地到固定沙丘,随着土壤水分和养分的减少,赖草通过降低分株密度来减少自身的营养消耗,从而维持亲株的生长,减小亲株的死亡风险。丘间地和固定沙丘中赖草的生长长期受土壤水分、有机质和养分条件的限制,分株密度和生物量较小。但是丘间地有大量人工种植的黑沙蒿和柠条锦鸡儿,资源的限制和种间的竞争导致赖草分株密度和生物量低于固定沙丘。
植物根系在不同深度土壤中的分布因自身的生长特性和生长环境的高度异质性而有所差异[21]。本研究中,高寒沙区低湿滩地、农田田边、丘间地和固定沙丘4种生境中赖草根系生物量主要分布于浅层土,这与刘爽等[22]的研究结果一致,即赖草根系生物量倾向于分布在0~40 cm土层。根系是植物直接感受土壤水分变化和吸收土壤水分的最主要的器官[23],具有向地、向水和向肥生长的特性。本研究区地处高寒干旱、半干旱地区,浅层土壤易受不定降水的补给,且浅层土壤肥力相对较佳,赖草根系在浅层大量生长并积累生物量。从低湿滩地、农田田边、丘间地到固定沙丘,随着土壤环境渐趋干旱,赖草根系为获取满足生长的水分采取深扎稳长的策略,逐渐向土壤深层延伸发展,固定沙丘中赖草根系可分布于50~60 cm土层,在持续的干旱胁迫下某些斑块的赖草根系甚至可伸到60 cm以下土层,通过吸收深层土壤的水分来维持植株生长。
综合研究结果认为,从低湿滩地、农田田边、丘间地到固定沙丘,土壤渐趋干旱和贫瘠,形成一定的梯度,在此梯度下生长的赖草表现出不同的生长特征,尤其根系表现出较强的表型可塑性。随着土壤水分、有机质和养分的减少,赖草采用灵活的吸取营养方式和积极的适应策略应对复杂多变的生境,维持最佳生长状态,如:增加间隔子长以吸收更远空间里的生长资源,增大根系分布深度以利用更深土层的水分,减小分株密度以减少自身的营养消耗。赖草在高寒荒漠地带中的生存发展对当地的防风固沙和生态修复发挥着重要作用,建议今后应推动抗逆新品种的开发和培育,为中国西部荒漠的植被恢复提供良好的生态服务效益。
猜你喜欢
环境污染与防治(2023年11期)2023-11-25 01:22:38
热带作物学报(2022年8期)2022-09-16 06:06:00
小猕猴智力画刊(2022年3期)2022-03-28 01:37:47
落叶果树(2021年6期)2021-02-12 01:29:10
中国农村水利水电(2018年12期)2018-12-28 06:04:04
发明与创新(2017年3期)2017-01-18 05:14:04
中国水土保持科学(2016年4期)2016-09-21 00:40:04
农家科技中旬版(2016年12期)2016-04-16 03:41:29
体育科技(2016年2期)2016-02-28 17:06:07
卫星与网络(2016年12期)2016-02-05 09:23:26
