
幼年东方田鼠长江亚种对苔草的取食研究
2018-08-22 07:19徐正刚张美文赵运林
农学学报 2018年8期
徐正刚,冯 蕾,王 勇,张美文,张 婉,赵运林
(1中南林业科技大学/湖南省环境资源植物开发与利用工程技术研究中心,长沙410004;2湖南城市学院材料与化学工程学院,湖南益阳413000;3中科院亚热带农业生态研究所,长沙410125)
0 引言
东方田鼠长江亚种(Microtus fortis)是洞庭湖稻作区的主要害鼠,严重危害农作物,传播疾病。东方田鼠具有季节迁移的特性,在洞庭湖水位变动的压力下,东方田鼠会周期性地改变栖息场所——低水位时主要栖息在洞庭湖洲滩,高水位时则被迫向农田迁徙。东方田鼠不同栖息地植被结构差异较大,为东方田鼠食物改变提供了基础。东方田鼠是广食性物种,在不同的栖息环境条件下食物结构会发生改变[1-6]。对该鼠食性进行研究,了解东方田鼠对不同食物的反应,在种群生态学上具有重要意义。
国内外对啮齿类动物食性研究已进行广泛研究,并为鼠害防治提供理论基础。姚丹丹等[7]对笼养板齿鼠(Bandicota indica)对一些作物果实和种子的喜好程度及食量进行测定,认为板齿鼠食谱极广,可通过营造同质生存环境,清除田间杂草的方法破坏其食物条件,防止鼠害发生;钟文勤等[8]通过分析草原啮齿类动物食性,以及分析草原啮齿类动物的危害程度,为其综合防治提供了理论依据;宛新荣等[9]对布氏田鼠(Microtus brandti)非取食性牧草的消耗量进行研究,结果显示:布氏田鼠春夏季对牧草以取食主,而秋冬季以筑巢为主。
此外,东北鼢鼠(Myospalax psilurus)[10]、高原鼠兔(Ochotona curzoniae)[11-13]、三趾跳鼠(Dipus sagitta)[14-15]、长爪沙鼠(Meriones unguiculatus)[16]等重要鼠害的食性均有相关研究。Rowe-Rowe等[17]发现,杂食性的小家鼠(Mus musculus)和褐家鼠(Rattus rattus)等啮齿动物作为海岛引进物种,对岛上的原有物种往往有着很大的影响;Roux等[18]研究了亚南极洲岛屿上的小家鼠的食性;Lathiya等[19]将啮齿动物食性研究的方法与谷仓中的鼠害相结合,发现仓库中的啮齿动物主要取食稻谷和昆虫,因此建议在使用灭鼠剂防治谷仓中或者野外鼠害时,可以在毒饵中加入杂拟谷盗(Tribolium confusum)和谷斑皮蠹(Trogoderma granarium)等昆虫的粉末,增加诱饵对害鼠的诱惑力。除此之外,还有、纳亚里特松鼠(Sciurus nayaritensis)[20]、缨耳松鼠(Sciurus aberti)[21]的食性报道。李俊年等[22]对东方田鼠的摄食量及觅食行为进行了研究,发现在饥饿条件下,植食性小哺乳动物是通过增加口量大小,提高摄食率来满足营养需求的。
邢廷杰等[23]研究发现,单宁酸、蛋白质、纤维素是影响植食性动物选择食物选择的重要因素,尤其是食物蛋白含量高低会显著影响东方田鼠幼鼠的体重。洞庭湖区东方田鼠主要在冬季繁殖,该时期东方田鼠幼鼠主要取食苔草[24]。因此,笔者以幼年东方田鼠为研究对象,在试验条件下研究不同性别幼年东方田鼠对苔草的取食,了解苔草对幼年东方田鼠的影响,以期为深入了解食物在东方田鼠种群暴发中的作用提供参考。
1 材料与方法
1.1 试验时间、地点
鼠类喂食试验于2016年4—9月在湖南长沙中科院亚热带农业生态研究所动物生态研究组鼠类实验室进行。苔草成分测定于中南林业科技大学稻谷及副产物深加工国家工程实验室。
1.2 试验幼鼠饲养
使用东方田鼠种群为经历多代驯化,建立的实验室封闭群。研究从中挑选体重近似的20~30日龄的幼年东方田鼠,雌雄各15只,要求健康、无伤。每一只东方田鼠单独饲养于37 cm×27 cm×17 cm的塑料盒中,以锯木屑为垫料,实验室温度18~25℃。在正式试验开始前进行预试验,喂食7天新鲜苔草(Carex brevicuspis),使幼鼠适应对苔草的取食。东方田鼠主要取食苔草地上部分的幼茎和叶,研究定义该部分为苔草的可食用部分,其他为不可食用部分。为便于实验,研究取苔草的可食用部分扎成小捆,固定于取食槽,进行喂食。苔草采集于洞庭湖洲滩。
开始取食试验前,先称量每只东方田鼠的初始体重,并连续喂食7天新鲜苔草。试验过程中确保苔草供给充足。试验中称量所喂新鲜苔草重量及取食后剩余苔草重量。同时,从喂食苔草中取少量样本,置于80℃烘箱烘干至恒重,分别测量新鲜苔草及剩余苔草的含水率。每天收集每只试鼠粪便,并称量粪便重量,将粪便置于80℃烘箱烘干至恒重并测量粪便含水率。最后称量试鼠1周后体重。
1.3 苔草营养成分测定
取洞庭湖苔草样,测定苔草可食用部分和不可食用部分粗蛋白、粗脂肪、粗灰分、钙及总磷含量。测量方法分别按照“GB/T 6432—94饲料粗蛋白测定方法”、“GB/T 6433饲料粗脂肪测定方法”、“GB/T 6438饲料中粗灰分的测定方法”、“GB/T 5009.92—2003食品中钙的测定方法”、“GB/T 6437饲料中总磷的测定分光光度法”等国标方法进行测定。
1.4 数据处理
计算取食量、单位取食量及单位排泄量等指标。各指标计算分别见公式(1)~(3)。

数据处理均采用SPSS 18.0统计软件进行,除雌鼠和雄鼠各自喂食前后体重变化采用配对样本t检验外,其他均值比较采用独立样本t检验进行分析。
2 结果与分析
2.1 不同性别幼年东方田鼠取食苔草后体重变化
试验中,雌性幼鼠初始体重为(74.51±8.98)g,1周后体重为(50.42±5.72)g,减少了(24.09±10.72)g,体重减少极显著(t=7.107,P<0.001)。雄鼠初始体重为(57.41±5.97)g,1周后体重为(62.97±7.99)g,增加了(5.56±11.35)g,但差异不显著(t=-1.549,P>0.05)(见图1)。在条件一致的情况下,雌性幼鼠体重明显减少,可能是由于雌性幼鼠对食物的敏感性较高,单一食物不能满足其生理需求。雄性幼鼠体重略有增加,可能是由于雄性幼鼠对食物的敏感性较低,单独食用苔草可维持基本生活。

图1 幼年东方田鼠取食1周后体重变化
2.2 不同性别幼年东方田鼠代谢
统计不同性别幼年东方田鼠对苔草取食量,发现每天取食量在4.83 g左右,差异不大(P>0.05)。雌性幼鼠单位取食量为(0.098±0.055)g/g,雄性幼鼠单位取食量为(0.075±0.033)g/g,雌性幼鼠比雄性幼鼠的单位取食量稍高,但无明显差异(t=1.144,P>0.05)(见表1)。雌性幼鼠比雄性幼鼠的单位取食量稍高,说明雌性幼鼠对营养的需求较雄性高。

表1 幼年东方田鼠取食率与代谢率
收集幼年东方田鼠取食苔草后排泄的粪便,测得含水率为15.28%。雌性幼鼠单位排泄量为(0.083±0.045)g/g,雄性幼鼠单位排泄量为(0.064±0.027)g/g。雌性幼鼠代谢率亦稍高于雄性幼鼠代谢率,但无明显差异(t=1.142,P>0.05)(见表1)。由于雌性幼鼠的单位取食量和单位排泄量均较雄性高,可初步判断雌性代谢活动较雄性更为活跃。
2.3 洞庭湖苔草的营养成分
苔草属莎草科,多生长于洞庭湖洲滩,是东方田鼠最喜食植物之一。研究测得喂食新鲜苔草的平均含水率87.29%,取食后剩余苔草的平均含水率13.79%,含水率显著下降。苔草可食用部分粗蛋白、粗脂肪和总磷均含量分别为(96.76±0.015)g/kg、(35.42±0.012)g/kg、(2.16±0.002)g/kg,苔草不可食用部分粗蛋白、粗脂肪和总磷均含量分别为(81.4±0.01)g/kg、(16.53±0.012)g/kg、(1.67±0.001)g/kg,苔草可食用部分粗蛋白、粗脂肪和总磷含量均高于苔草不可食用部分,其中苔草可食用部分粗脂肪含量是苔草不可食用部分的2倍以上。苔草可食用部分的粗灰分与钙的均含量分别为(68.47±0.502)g/kg、(4.26±0.006)g/kg,苔草不可食用部分的粗灰分与钙的均含量分别为(122.12±0.95)g/kg、(4.95±0.006)g/kg,其可食用部分含量小于不可食用部分,且粗灰分含量差异极显著(t=-92.275,P<0.001)(见图2)。
3 讨论与结论
研究探讨了幼年东方田鼠对苔草的取食。研究发现雌性幼鼠喂食1周苔草后体重明显降低,而雄性幼鼠体重略有增加。幼年雌性东方田鼠的单位取食量及单位排泄量会稍高于成年雌性东方田鼠,这可能是由于幼年雌性东方田鼠对食物的营养要求比成年雄性东方田鼠对食物的营养要求更高。食物中所含蛋白质是有机体维持新陈代谢的主要营养物质之一,食物蛋白含量的高低显著影响幼体体重[25]。本研究中,幼年东方田鼠所食苔草经测定粗蛋白的含量为9.68%,与陈竞峰等[25]设定的10%低蛋白食物含量差异不大。因此,引起幼年雌鼠体重下降的原因,可能是由于其他营养物质不足所导致的。也进一步说明不同性别幼年东方田鼠对营养的需求不同。本试验中,幼年东方田鼠经过长期喂食苔草,发现在15~20天左右开始出现死亡现象,也可能是由于酚类化合物副作用导致的。胡忠军等的研究表明洞庭湖东方田鼠性比在野外是偏雄的,但在实验条件下是偏雌的[26]。可能是由于雌性幼年东方田鼠对环境更加敏感,死亡率较高导致的,与本研究结果一致。
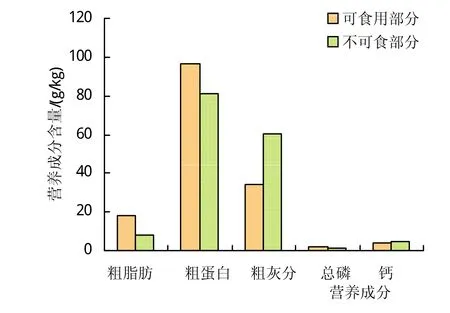
图2 洞庭湖苔草营养成分
研究认为,即使每天供应充足苔草作为食物,单一苔草作为食物来源无法满足幼年东方田鼠对营养的需求。同时,在幼年阶段,雌性东方田鼠比雄性东方田鼠对环境更加敏感。通过对不同性别幼年东方田鼠对其食性研究,能够从取食规律以及营养需求方面揭示东方田鼠暴发内在规律,为农业鼠害防治提供一些依据。
猜你喜欢
海南医学院学报(2022年18期)2022-10-13
广东药科大学学报(2022年4期)2022-07-22
当代水产(2021年10期)2022-01-12
作文大王·低年级(2021年11期)2021-12-22
小天使·一年级语数英综合(2021年2期)2021-05-08
郑州大学学报(医学版)(2021年1期)2021-03-03
小学生(看图说画)(2020年4期)2020-04-01
中国油脂(2019年4期)2019-04-29
教育教学论坛(2016年49期)2017-02-27
星星·散文诗(2016年23期)2016-12-29
