
利用同源重组构建BMP6基因真核表达载体及在成纤维细胞中表达量的研究
2018-08-20 06:05:28张梦瑶刘开东刘积凤贺建宁
中国畜牧杂志 2018年8期
张梦瑶 ,杨 峰,刘开东,刘积凤,柳 楠,贺建宁*
(1.青岛农业大学动物科技学院,山东青岛 266109;2.内蒙古农业大学动物科技学院,内蒙古呼和浩特 010018;3.青岛畜牧兽医研究所,山东青岛 266121)
随着生活水平的提高,人类对羊毛的需求量和品质要求不断提升,羊毛的产量与质量问题越来越引发关注。羊毛产量由毛囊组织结构和性状共同决定,探究毛囊发育的规律和结构特征是研究毛囊生长发育调控机制的基础,同时毛囊发育对羊毛性状有重要影响[1-3]。敖汉细毛羊作为最具有代表性的细毛羊之一,研究其毛囊发育的调控机制具有重要价值。近年来,Stenn等[4]在成年绵羊身上对毛囊的周期性生长变化展开了研究,得出初级毛囊的密度活动变化较次级毛囊恒定;柳楠等[5]在分子水平上研究了敖汉细毛羊羊毛性状的形成机理,证实毛囊始发、形成与循环均共用一套调控网络,均需经历生长期、退行期和静止期;王小佳等[6]在分子层面上对细毛羊羊毛的生长部位进行了研究,得出BMPs主要表达于表皮上皮细胞和内根鞘细胞中,在肩部和腹股沟部表达较明显,且在肩部表达量较高,差异倍数在2倍以上。
有研究报道,BMPs在大量动物的卵巢、前列腺、皮肤等组织中均能检测到。曾德会等[6]研究发现,BMP6在BMP6-HJV-Smad信号通路调节铁稳态中发挥着重要作用。BMP6同时也是TGF-β超家族中的一员[7],在TGF通路中可以促进基因表达的同时也抑制通路中某些基因的表达。杨燕燕等[8]研究蒙古羊是否双羔直接受BMP6基因的控制,发现BMP6在发情期表达尤其明显,特别是双羊羔群体中。目前,对BMP6基因与家畜皮肤毛囊关系的研究主要集中在分子水平上,因此对BMP6基因在细胞水平进行研究十分必要。本研究通过同源重组技术构建BMP6真核表达载体及瞬时转染成纤维细胞来进行BMP6基因表达的定量定性分析,为此后个体层次上的研究提供坚实基础。
1 材料与方法
1.1 实验材料 选取健康敖汉细毛羊40日龄的胎羊(由青岛畜牧所奥特种羊场提供)。 TRIZol(Roche)试剂、RIPA、抗体、EcoR1限制性内切酶、pcDNA3.1质粒、标准胎牛血清、Lipofectamine2000等均购自青岛鑫恒一有限公司。Leica DMIRB 型倒置显微镜购于青岛鑫恒一有限公司、Redbio CFX ConnectTM荧光定量PCR系统购自Bio-Rad公司。
1.2 实验方法
1.2.1 引物设计 利用试剂盒对RNA进行提取并反转录。选取绵羊BMP6基因组(GenBank登录号为XM-015102858.1)CDS区、结合pcDNA3.1载体序列,在CDS区外侧通过Primer Premier 5.0设计引物,在上下游引物5'端加入同源臂,完成后的引物序列:上游引物:5'-TTTAAACTTAAGCTTGGTACCATGATTCCTGGTA ACCGAATGCTG-3',下游引物:5'-TGGATCCGAGCTC GGTACCTCAGCGGCACCCACATCCCTCCACTA-3',然后进行PCR扩增及琼脂糖凝胶电泳检测,25 μL扩增体系:DNA模板1 μL(2 183.1 μg/mL),上、下游引物各1 μL(10 μmol/L),GreeMix 12.5 μL(4mmol/L),ddH2O 9.5 μL。
1.2.2 BMP6基因与pcDNA3.1质粒同源重组 切胶回收PCR扩增产物,pcDNA3.1质粒同时进行EcoR1限制性单酶切。酶切体系:EcoR1 1 μL,pcDNA3.1 5 μL,Buffer 5 μL,ddH2O 39 μL;37℃ 2 h。连接体系:BMP6基因 1 μL,pcDNA3.1 1 μL,SoSo 5 μL,ddH2O 3 μL,50℃ 10 min。转化 DH5ɑ感受态细胞,挑单菌摇菌并进行菌液PCR鉴定阳性率,将阳性组送往公司测序,并通过质粒提取试剂盒提取阳性质粒。
1.2.3 成纤维细胞的培养 取40日龄胎羊用75%酒精和PBS清洗。去除头部和四肢剩余躯干用眼科剪将其剪成1.5~2.0 mm3大小的组织块,滴加37.0℃预热的胎牛血清,放置到CO2培养箱(37℃,5.0% CO2)培养4 h后加培养液。24 h进行换液,每天定时在倒置显微镜下观察细胞生长情况,当细胞密度达到约95%进行传代。弃掉旧培养液,用PBS洗涤3次,0.25%的胰蛋白酶37.0℃消化4 min,培养液终止消化。按1:3传代,置于CO2恒温培养箱中待细胞密度达95%后,再次传代培养[9]。
1.2.4 细胞转染 用Lipofectamine2000进行瞬时转染。孵育体系:实验组为脂质体20 μL、DMEM(不含双抗)480 μL;对照组为脂质体20 μL、DMEM(不含双抗)480 μL,混匀,静置10 min。转染体系:实验组为pcDNA3.1-BMP6 40 μL、DMEM(不含双抗)460 μL;对照组为DMEM 500 μL,静置20 min;各加入4 mL DMEM,37℃培养6 h后换液,培养36 h。
1.2.5 细胞转染后实时荧光定量PCR 对转染的细胞进行培养,待细胞密度达到95%时用TRIZol(Roche)裂解细胞提取RNA。利用核酸测定仪测定浓度和纯度,用试剂盒(Roche)将RNA反转录成cDNA。根据敖汉细毛羊BMP6基因的全序列和绵羊GAPDH的全序列,使用PrimerPremier5.0软件设计BMP6和GAPDH基因的引物序列见表1。
实时荧光定量PCR反应程序:95℃预变性7 min;95℃变性 40 s,60℃退火40 s,72℃ 延伸30 s,共45个循环。样品各进行3次重复,采用Ct(2-ΔΔCt)值法计算BMP6相对于内参基因GAPDH的表达量。利用SPSS 19.0软件中t检验对数据进行差异显著性分析,结果以平均值±标准误表示,以P<0.01作为差异极显著性判断标准[9]。
1.2.6 细胞转染后Western Blotting检测BMP6蛋白表达 样品分为对照组和实验组,每个样品3个平行。首先用细胞裂解液RIPA提取细胞中蛋白,β-actin作为内参。对提取的蛋白进行转膜,用5%BSA对蛋白进行封闭2 h;TBST振荡洗膜4次/5 min;将一抗按1:2 000进行稀释,孵育PDF膜4℃ 12 h;TBST振荡洗膜4次/5 min;山羊抗兔的二抗按1:2 000稀释,孵育PDF膜1.5 h;曝光液1:1的比例配置,在暗室中进行曝光拍照。所得结果用Image J软件分析灰度值,灰度值的比值用SPSS19.0统计分析软件进行t检验。

表1 引物信息
2 结 果
2.1 BMP6目的基因扩增 以敖汉细毛羊cDNA为模板,对目的基因片段进行PCR 扩增。利用2%琼脂糖凝胶电泳分析,结果表明,引物扩增的条带明亮无杂带,产物大小为537 bp (图1),与已知的目的基因片段大小相同,可用于后期实验。

图1 BMP6基因PCR扩增
2.2 pcDNA3.1载体单酶切及纯化 对pcDNA3.1进行单酶切,酶切后利用2%琼脂糖凝胶电泳检测,如图2所示,1、2是对pcDNA3.1载体的EcoR1单酶切,酶切效果良好,pcDNA3.1载体5 428 bp,与图中条带所吻合,显示酶切成功,切胶回收可用于后期连接实验。

图2 pcDNA3.1载体单酶切鉴定
2.3 表达载体pcDNA3.1与目的片段重组及鉴定 对pcDNA3.1载体单酶切的片段进行胶回收后用SoSo试剂盒将目的片段和pcDNA3.1载体进行连接,对菌液进行PCR电泳鉴定,如图3所示,在537 bp处有单一明亮的条带,为BMP6基因。菌液送至公司进行基因测序,利用BLAST与已知目的基因的序列进行比对,表明目的基因与pcDNA3.1载体连接成功(图4)。同时对重组的质粒进行提取, 1~2为pcDNA3.1-BMP6重组质粒,大小为5 965 bp,与图中条带相符(图5),表明重组质粒构建成功。

图3 表达载体pcDNA3.1的鉴定-PCR扩增图

图4 表达载体菌液的测序对比

图5 pcDNA3.1-BMP6重组质粒的提取
2.4 细胞培养 定时用电子倒置显微镜观察成纤维细胞的生长情况。细胞生长24 h可观察到细胞慢慢贴壁,有圆形的,有梭形的,还有一些游离组织。培养4 d后,细胞生长良好,细胞密度达到大约95%时对细胞进行传代(图6)。经传代后细胞开始生长速度稍慢,12 h后增殖迅速加快,细胞形态仍然呈现梭形,生长情况良好,可用于下一步转染(图7)。
2.5 实时荧光定量PCR检测BMP6基因表达 以成纤维细胞的cDNA为模板,对目的基因BMP6和内参基因GAPDH片段进行PCR 扩增。2%琼脂糖凝胶电泳分析表明,引物扩增的条带明亮,GAPDH产物大小为125 bp ,BMP6产物大小为116 bp(图8),与已知的内参基因、目的基因大小均相同,可继续进行实时荧光定量PCR实验。

图6 成纤维细胞原代培养(×100)
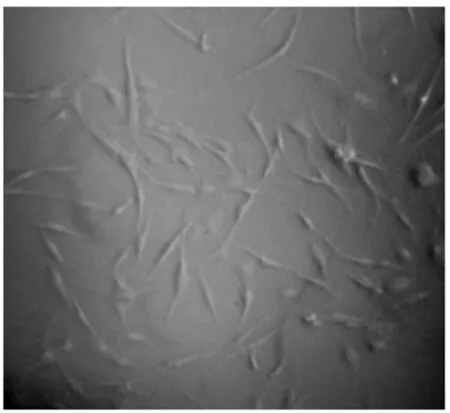
图7 成纤维细胞传代培养(×100

图8 内参基因GAPDH(A)目的基因BMP6扩增产物(B)
目的基因BMP6和内参基因GAPDH的扩增曲线、熔解曲线如图9所示,清晰整齐,并且目的基因和内参基因的熔解曲线上未出现杂乱的峰,表明在实时荧光定量PCR过程中引物特异性较高,没有产生二聚体,由此得到了特异性的扩增产物,因此设计的引物可以用来进行定量分析。如图10所示,BMP6基因经过质粒构建转染成纤维细胞后与未转染的细胞之间差异极显著(P<0.01)。
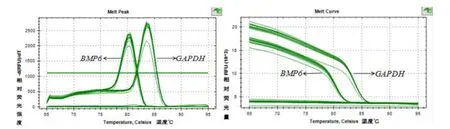
图9 BMP6和GAPDH 溶解曲线峰值图(A)和扩增曲线图(B)

图10 BMP6基因mRNA相对表达量
2.6 BMP6蛋白表达的检测 以成纤维细胞提取的蛋白为模板进行Western Blotting,结果显示实验组细胞瞬时转染BMP6基因的蛋白条带明显亮于未转染的对照组(图11),经ImageJ和SPSS19.0分析得实验组细胞瞬时转染BMP6基因蛋白的表达极显著高于未转染组(P<0.01)(图 12),说明质粒 pcDNA3.1-BMP6 能高效表达BMP6 基因。

图11 BMP6 基因表达蛋白条带
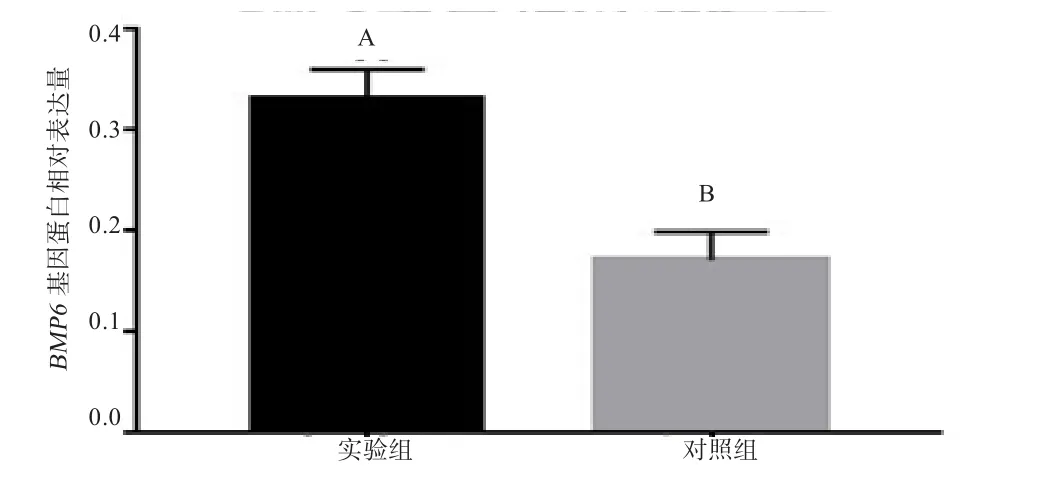
图12 BMP6基因蛋白相对表达量
3 讨 论
目前,调节毛囊形态发生的信号分子主要分为Wnt通路、TNF家族、FGF家族、BMP家族、Shh通路、TGF家族和NOTCH通路[10]7类,这些信号分子之间都会相互作用调控,有些是毛囊发育的促进物,有些是抑制物。崔向荣等[11]研究证明,BMP6参与颗粒细胞凋亡,BMP6蛋白突变易导致细胞的凋亡,从而引起一系列相应机制调节。以小鼠为模型研究,BMPs在毛球间质细胞和表皮里广泛表达[8],主要是在胚胎期毛囊形成和出生后毛囊周期循环过程中起作用[12]。本研究利用成纤维细胞,通过实时荧光定量PCR,在mRNA的水平上检测BMP6基因在成纤维细胞中过表达。王小佳等[13]验证了BMP4基因能够在细毛羊的腹股沟和肩部均抑制毛囊发育。BMP6基因和BMP4同属于BMP家族,是一种抑制毛囊发育的信号分子。本研究在细胞水平对基因的表达进行验证,经转染成纤维细胞后BMP6基因蛋白表达量明显升高。BMPs信号不仅调控着上皮细胞的增殖与分化,维持着上皮内环境的稳态[14],还控制着毛发角质细胞的细胞增殖,进而毛囊毛乳头的大小[15]、毛发类型和毛纤维直径将受到BMPs信号的控制。Noggin作为BMP信号通路中的抑制剂,进而诱导次级毛囊发育[16],Noggin缺失,BMP信号通路中的基因会增强表达,抑制毛发的生成,因而出现严重的光秃现象,这一现象极力阐述了BMP信号对毛发的抑制。BMP还会通过Smad类等细胞内信号分子以及Wnt等相邻信号通路对表皮和毛囊发育进行调节[17]。许多研究证明,BMP6基因在毛囊发育过程中起着十分重要的作用。
本文研究借助前人的研究结果对BMP6在细胞层次上进一步研究,选择40日龄的胎羊培养的成纤维细胞生长能力强,活力高,容易获得,结果稳定性更高,实验周期短。实验创新点主要是利用同源重组技术构建表达载体,相比TA克隆技术更为简便快速,不会引入多余的碱基,目的基因可以更准确地定位在靶位点上[18]。本研究结果显示,转染组的mRNA量极显著高于未转染组;其次,已知BMP6蛋白分子量约为19.6 ku,经过转染的细胞BMP6基因表达量明显高于空白对照。经过显色曝光后条带明显比对照组的条带黑、宽。经Image J分析,相对表达的灰度值也明显高于对照组。将目的基因与内参基因的灰度值的比值进行SPSS分析,结果显示差异极显著,这进一步说明了转染后BMP6基因的过表达。本研究中,经过转染,BMP6基因的mRNA和蛋白表达量明显升高,这从细胞层次上研究了与羊毛有关的基因。BMP6基因具有抑制毛囊发育的作用,本研究在细胞层次上验证了BMP6基因的过表达,其过表达的同时也将影响相关信号通路的调节。作为上、下调基因,BMP6基因过表达会影响它直接控制的基因也进行相应的表达增强、减弱,这也为以后个体水平上的研究提供坚实的基础,有助于解决绵羊羊毛生长的问题。
4 结 论
研究通过利用同源重组技术对载体pcDNA3.1-BMP6的构建、载体转染成纤维细胞后进行实时荧光定量PCR、Western Blotting等,成功地构建了表达载体pcDNA3.1-BMP6并转染成纤维细胞,转染后细胞生长状态良好,BMP6基因过表达,mRNA表达量和蛋白的表达量均极显著高于对照组,为进一步在个体水平上研究其功能奠定基础。
猜你喜欢
军事文摘(2023年2期)2023-02-17 09:20:24
昆明医科大学学报(2021年8期)2021-08-13 08:59:50
感染、炎症、修复(2021年1期)2021-07-28 06:18:08
云南医药(2021年3期)2021-07-21 05:40:30
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
保健与生活(2016年1期)2016-04-12 18:29:44
中国现代医学杂志(2015年26期)2015-12-23 11:04:22
中国光学(2015年1期)2015-06-06 18:30:20
中国当代医药(2015年33期)2015-03-01 02:09:20
海岸工程(2014年4期)2014-02-27 12:51:28
