
毛竹入侵森林对固氮微生物群落结构和丰度的影响
2018-08-16 10:03刘彩霞徐秋芳梁辰飞陈俊辉李永春
植物营养与肥料学报 2018年4期
周 燕,刘彩霞,徐秋芳,梁辰飞,陈俊辉,秦 华,李永春
(浙江农林大学环境与资源学院/浙江省森林生态系统碳循环与固碳减排重点实验室,浙江临安 311300)
毛竹 (Phyllostachys pubescens) 是中国的乡土物种[1],栽培历史悠久、分布面积广,主要生长于闽、赣、浙、湘等南方省份[2],是十分重要的经济树种之一。因其在食品、工艺美术、建筑等方面均具有重要的经济价值,从300多年前开始,被陆续引种至日本及一些欧美国家[1]。有关毛竹入侵周边林地的现象,最早在1996年报道于日本[3]。中国同样存在着各种程度的毛竹入侵现象,有越来越多的阔叶林、杉木林等天然林地被毛竹林所替代。江西的大岗山[4]、井冈山自然保护区[5],江苏的龙背山森林公园[6]以及浙江的天目山自然保护区等都有关于毛竹入侵到周边林地的报道。根据国内外不同地区的调查显示,毛竹林前沿向周边林地蔓延的速度基本可达到每年1~2 m[3,7–8]。
毛竹以无性繁殖为主,地下竹鞭具有很强的繁殖能力,横向蔓延至周边长笋、成竹。母竹通过竹鞭运输同化产物提供给新竹,未展叶新竹以及矮小的新竹不会因无法进行光合作用而死亡[9],在毛竹的成竹阶段,土壤—竹叶—大气间形成了强大的水势差,及时补充新叶充足的水分,促进毛竹快速生长[10]。另外有研究发现,毛竹产生的一些化感物质对其他植物的生长产生潜在的抑制作用[11]。以上这些毛竹的性质都是引起毛竹向周边林地不断扩张、最后完全代替原树种的重要因素。植物入侵会影响入侵地的原有生态系统,甚至导致本地植物灭绝,使得生态系统失衡[12],这种情况也可能发生在毛竹入侵的过程中。
在中国,已有不少研究者关注毛竹入侵后对原有生态系统所造成的各方面影响。杨清培等[4]研究发现大岗山因毛竹入侵常绿阔叶林导致森林生态系统总碳储量减少了15.37%,其中土壤碳储量减少了17.30%;欧阳明等[5]报道毛竹入侵导致井冈山自然保护区次生常绿阔叶林群落组成和结构简化、物种多样性下降;史纪明等[6]调查发现毛竹入侵限制了龙背山杉木的生长。位于浙江省杭州市临安区西北部的天目山国家自然保护区,从1985年起就有学者研究毛竹入侵周边林地与生物多样性之间的矛盾问题[13]。在动物方面,杨淑贞等[14]调查发现保护区内毛竹蔓延导致了鸟类种类和数量的减少。在植物多样性方面,林倩倩等[15]发现受毛竹入侵影响,保护区的阔叶林、针叶林等林地内,乔木层的物种丰富度、多样性和均匀度呈现降低趋势,而灌木层物种的各项多样性指数则上升,草木层无变化。吴家森等[16]研究毛竹入侵对土壤肥力的影响,发现土壤的各个肥力指标在毛竹入侵过程中变化并不一致,其中碱解氮含量上升,而有效磷和速效钾则下降。土壤中的微生物也会受到不同程度的干扰。Xu等[17]通过不同实验手段研究土壤微生物群落,表明在毛竹入侵影响下土壤微生物的生物量和多样性均增加,但功能多样性并无显著改变。潘璐等[18]研究报道,毛竹扩张不会减弱菌根侵染林木的频率和强度。土壤中细菌的群落结构和多样性同样不受毛竹入侵的显著影响[19],但真菌的数量在毛竹入侵后显著下降[20]。
土壤固氮微生物是土壤中一类重要的功能性微生物,能够直接将空气中的氮还原成氨供给植物吸收利用[21]。氮是植物生长的必需营养元素,与毛竹的出笋、成竹过程有着密切的关系[22],在毛竹入侵周边林地的过程中,土壤的氮素矿化和有效性都会受到影响[23]。在没有人为经营的林地中,生物固氮是主要的氮素来源,因而,土壤固氮微生物的群落结构和多样性是评价森林生态功能十分重要的参数。因此,我们假设,毛竹入侵会引起林地土壤固氮微生物群落的变化。
本研究选择天目山保护区作为试验地,分别采集毛竹林、毛竹与原林混交林、原有林 (阔叶林或针阔混交林) 土壤,利用末端限制性片段长度多态性(terminal restriction fragment length polymorphism,T-RFLP)技术和荧光定量 PCR (quantitative PCR,qPCR) 方法,揭示毛竹入侵过程中土壤固氮微生物群落结构和丰度的变化,为毛竹入侵对生态功能影响评价提供理论依据。
1 材料与方法
1.1 研究区概况
天目山国家自然保护区 (30°18′~30°25′N,119°23′~119°29′E),属中亚热带向北亚热带过渡型气候区,海拔300~1500 m,年平均气温8.8~14.8℃,多年平均降水量1390~1870 mm,无霜期209~235 d,年太阳总辐射3770~4460 MJ/m2,这些气候条件适合毛竹的生长繁衍。保护区内的植被类型分为常绿阔叶林、常绿落叶阔叶混交林、针叶林、落叶阔叶林、竹林 (主要为毛竹林) 等[24],土壤母质主要为凝灰岩类、花岗岩类、砂泥质岩类、碳酸盐岩类和红色砂砾岩类五种风化残坡积物类型[25]。天目山自1956年林区禁伐建立为自然保护区后,原先由人为种植的毛竹在没有人为干扰下不断蔓延生长,至2014年近60年间,毛竹林面积已从55.1 hm2扩大至 105.4 hm2[26]。
在保护区内,选择三个典型的毛竹入侵带 (大约相距2公里),入侵带一为青龙山,原林地为针阔混交林,土壤母质为泥质灰岩,林下灌木比较丰富;入侵带二为石门洞,原林地为阔叶林,母质为泥质灰岩,纯毛竹林中灌木很少,有大量死竹,而混交林和阔叶林中有较多灌木;入侵带三为进山门,原林地为阔叶林,母质为石英斑岩,林下植被和样地二类似。
1.2 土壤样品采集与处理
在三个入侵带中,依次沿毛竹入侵的方向分别选取三种林分:毛竹纯林 (BB)、混交林 (BBBL)、阔叶林或针阔混交林 (BL),每种林分中各划定6个10 m × 10 m 的样地,样地间隔距离 40 m,各个样地中选取5个采样点,刮去表层枯枝落叶,采集0—20 cm的表层土壤[20]。5个采样点的土壤充分混匀后过2 mm筛作为一个土壤样品,共计54个土样。新鲜土样取一小部分经冷冻干燥后保存用于提取微生物DNA,其余自然风干用于测定土壤基本理化性质等。
1.3 样品分析方法
1.3.1 土壤基本化学性质的测定 基本的土壤化学性质参考文献[27]进行测定:土壤pH值测定采用1∶2.5土水质量比,用复合电极测定;有机碳含量采用K2Cr2O7–浓H2SO4外加热法测定;碱解氮采用碱解扩散法;有效磷采用Bray法,HCl-NH4F溶液浸提,钼锑抗比色法测定;速效钾采用NH4Ac浸提,火焰分光光度计测定;硝态氮、铵态氮用2 mol/L KCl溶液浸提,前者用紫外分光光度法,后者用靛酚蓝比色法测定。
1.3.2 土壤总 DNA 的提取 用 PowerSoil®DNA Isolation Kit试剂盒 (Mo Bio,美国) 提取土壤总 DNA。用NanoDrop核酸蛋白测定仪 (ND-1000)(ThermoFisher,美国) 测定DNA的浓度和纯度,所有样品浓度平均值为 40~120 ng/μL。
1.3.3 土壤固氮微生物的群落分析 土壤固氮微生物的群落分析采用T-RFLP技术,PCR反应体系如下:Premix TaqTM(TaKaRa,大连) 25 μL,采用引物nifHF (AAAGGYGGWATCGGYAARTCCACCAC)和 nifHRb (TGSGCYTTGTCYTCRCGGATBG GCAT)[28](10 μmol/L) 各 0.5 μL,BSA (Bovine Serum Albumin Solution)(TaKaRa,大连)0.5 μL,模板 DNA 1 μL,用灭菌水补充至 50 μL。其中,nifHF 的 5′端带有FAM荧光标记。PCR反应采用Touch-down PCR 程序,步骤如下:95℃ 5 min,94℃ 1 min,60℃ 1 min,72℃ 1 min,之后退火温度每个循环降低1℃,直到54℃;接下来按照54℃退火温度循环28 次,72℃ 10 min。
采用SanPrep柱式PCR产物纯化试剂盒 (生工,上海) 纯化PCR产物。选择限制性内切酶HaeⅢ[29](TaKaRa,大连)。酶切体系如下:酶 1.5 μL,Buffer 3 μL,纯化产物 20 μL,用无菌双蒸水补充至 30 μL。酶切过程:37℃ 12 h,65℃ 20 min。酶切完成,将产物送至生工生物工程 (上海) 股份有限公司进行检测分析。
1.3.4 土壤固氮微生物的丰度分析 以土壤总 DNA为模板,采用qPCR法测定固氮微生物nifH基因拷贝数。反应体系如下:SYBR®Premix Ex TaqTM10 μL,采用同 T-RFLP 的引物 (50 μmol/L) 各 0.2 μL,模板 DNA 1 μL,用无菌双蒸水补充至 20 μL。使用Bio-Rad CFX96TMThermal Cycler仪器 (Bio-Rad,美国) 进行扩增,每个样品进行3次重复。反应程序如下:95℃ 10 min;95℃ 30 s,56℃ 30 s,72℃ 30 s,循环39次。标准曲线制作:用EasyPure Plasmid MiniPrep Kit (全式金,北京) 提取含有插入 nifH 基因片段的质粒 DNA,按梯度 (10–1~10–7) 进行稀释,选择其中的10–2~10–7梯度做标曲。三个入侵带的扩增效率分别为95.9%、104.8%、101.3%,R2分别为0.999、0.993、0.998。
1.4 数据处理
采用SPSS18统计软件进行显著性检验和Pearson相关性分析,使用Duncan法进行多重比较。T-RFLP数据处理方法:计算每一个T-RF的百分比 (± 1.5 bp 的可归为一个 T-RF);去除小于 1% 的部分,以及低于 45 bp (杂峰、引物峰) 和高于 400 bp(引物长度) 的部分;6个重复间重现性不高的部分也去除;做接下来的分析[29]。使用BIO-DAP软件计算微生物的Shannon指数、Simpson指数和均匀度指数。采用R软件进行多响应置换过程分析 (multiresponse permulation procedures,MRPP),采用Canoco 4.5软件进行微生物与土壤化学性质之间的冗余分析 (redundancy analysis,RDA),其中均采用999次的蒙特卡洛置换检验显著性。
2 结果与分析
2.1 毛竹入侵对土壤化学性质的影响
如表1显示,在毛竹入侵过程中,土壤pH和有机碳含量在三个入侵带中变化规律一致,表现为毛竹纯林最高,混交林次之,原林地最低,其中青龙山和石门洞入侵带毛竹与原林之间差异显著 (P <0.05);土壤碱解氮和铵态氮含量在青龙山入侵带均随毛竹入侵显著上升,毛竹与原林的差异显著 (P <0.05),而在石门洞和进山门入侵带总体上毛竹高于原林,但差异不显著;土壤有效磷含量在石门洞和进山门入侵带随毛竹入侵上升,而青龙山则入侵前后差异不大;毛竹入侵后速效钾含量在三个入侵带均呈下降趋势,其中石门洞和进山门差异显著 (P <0.05);硝态氮的变化趋势均不显著。总体上,入侵带土壤的养分含量在毛竹入侵后有所上升。
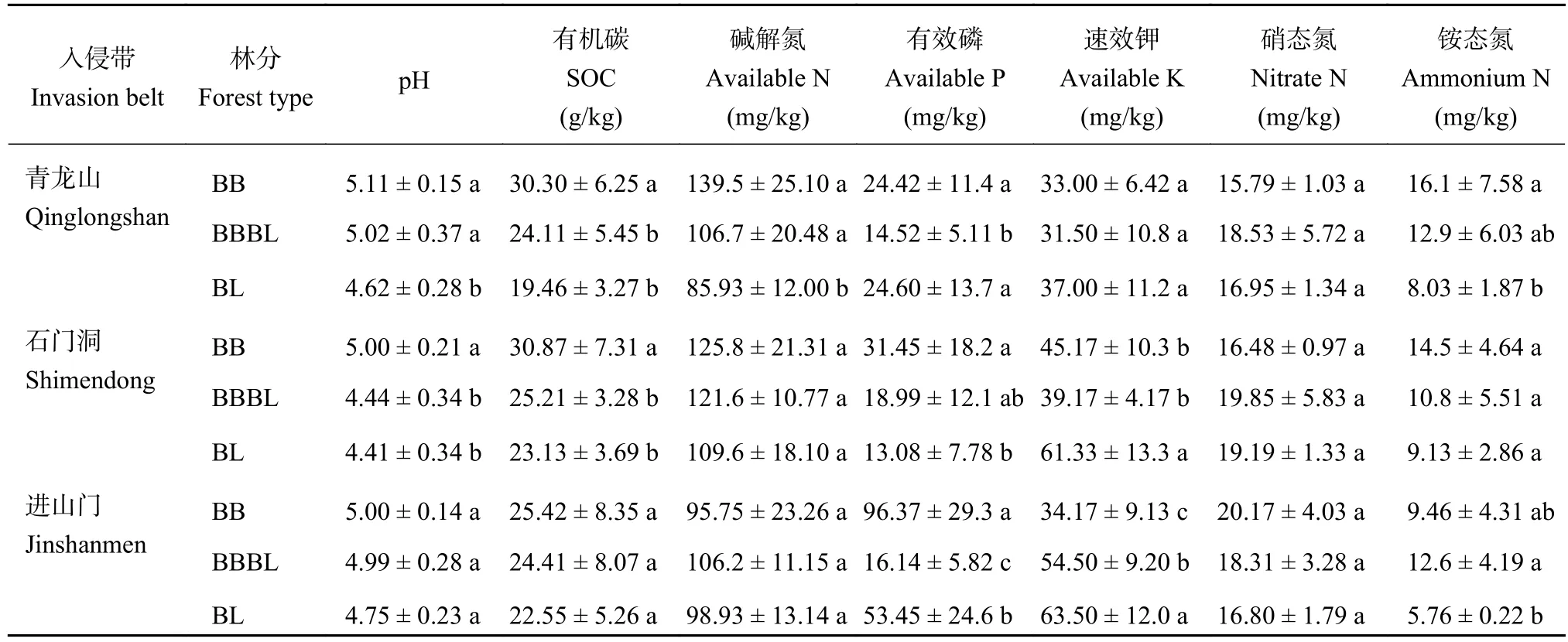
表1 三个入侵带土壤化学性质Table1 Soil chemical properties of the three invasion belts
2.2 毛竹入侵对土壤固氮微生物群落结构的影响
如图1显示,青龙山、石门洞和进山门三个入侵带土壤中分别筛选出18、16和19个T-RFs (terminal restriction fragments),其中 48 bp 在三个入侵带中均占绝对优势,青龙山和石门洞土壤的相对百分比高达40%左右 (36%~45%),进山门相对较低 (13%~30%),青龙山和进山门入侵带毛竹低于阔叶林,石门洞则相反,毛竹入侵前后土壤48 bp的变化趋势并不一致。三个入侵带土壤中除了共有的T-RFs 外,还拥有各自独特的T-RFs,这些T-RFs在毛竹入侵过程中变化各异。青龙山入侵带土壤减少2个T-RFs(267 bp在混交林土壤中消失,182 bp在毛竹纯林中消失),增加1个 T-RF (84 bp 在毛竹纯林中出现),其他T-RFs变化不显著;石门洞减少4个T-RFs (141 bp、182 bp 和 237 bp 在混交林中消失,125 bp 在毛竹纯林中消失),增加1个T-RF (109 bp在混交林中出现),其他T-RFs变化不显著;进山门减少3个TRFs (69 bp、304 bp 和 341 bp 均在毛竹纯林中消失),增加2个 T-RFs (74 bp、122 bp 在混交林中出现),182 bp 显著减少 (P<0.05),57 bp 显著上升 (P<0.05)。
将三个入侵带的土壤固氮微生物T-RFLP数据分别按林分分组 (BB、BBBL、BL) 进行MRPP分析(表2)。青龙山入侵带的土壤固氮微生物群落结构在混交林和原林地之间差异最大,其次是毛竹纯林和混交林之间,但均无显著差异;石门洞入侵带中,毛竹纯林和原林地 (P = 0.005)、混交林和原林地 (P =0.021) 之间的土壤固氮微生物群落结构有显著差异;进山门入侵带中,毛竹纯林和混交林 (P = 0.002)、原林地 (P = 0.032) 之间都有显著差异。将土壤固氮微生物T-RFLP数据按不同入侵带分组 (青龙山、石门洞、进山门) 进行MRPP分析,结果表明,三个入侵带的土壤固氮微生物群落结构两两之间均具有极显著差异 (P = 0.001)。
2.3 毛竹入侵对土壤固氮微生物多样性指数影响
根据T-RFLP数据计算得出土壤固氮微生物的Shannon 指数 (H)、Simpson 指数 (D) 和均匀度指数(E)(表3),分别表征微生物群落的丰富度、优势度和均匀度。青龙山入侵带土壤固氮微生物的三个多样性指数在毛竹入侵影响下均无显著变化;石门洞和进山门的Shannon指数和均匀度指数在原林地中最高,毛竹纯林中最低,Simpson指数的变化相反,其中石门洞的变化显著 (P<0.05)。
2.4 毛竹入侵对土壤固氮微生物丰度的影响
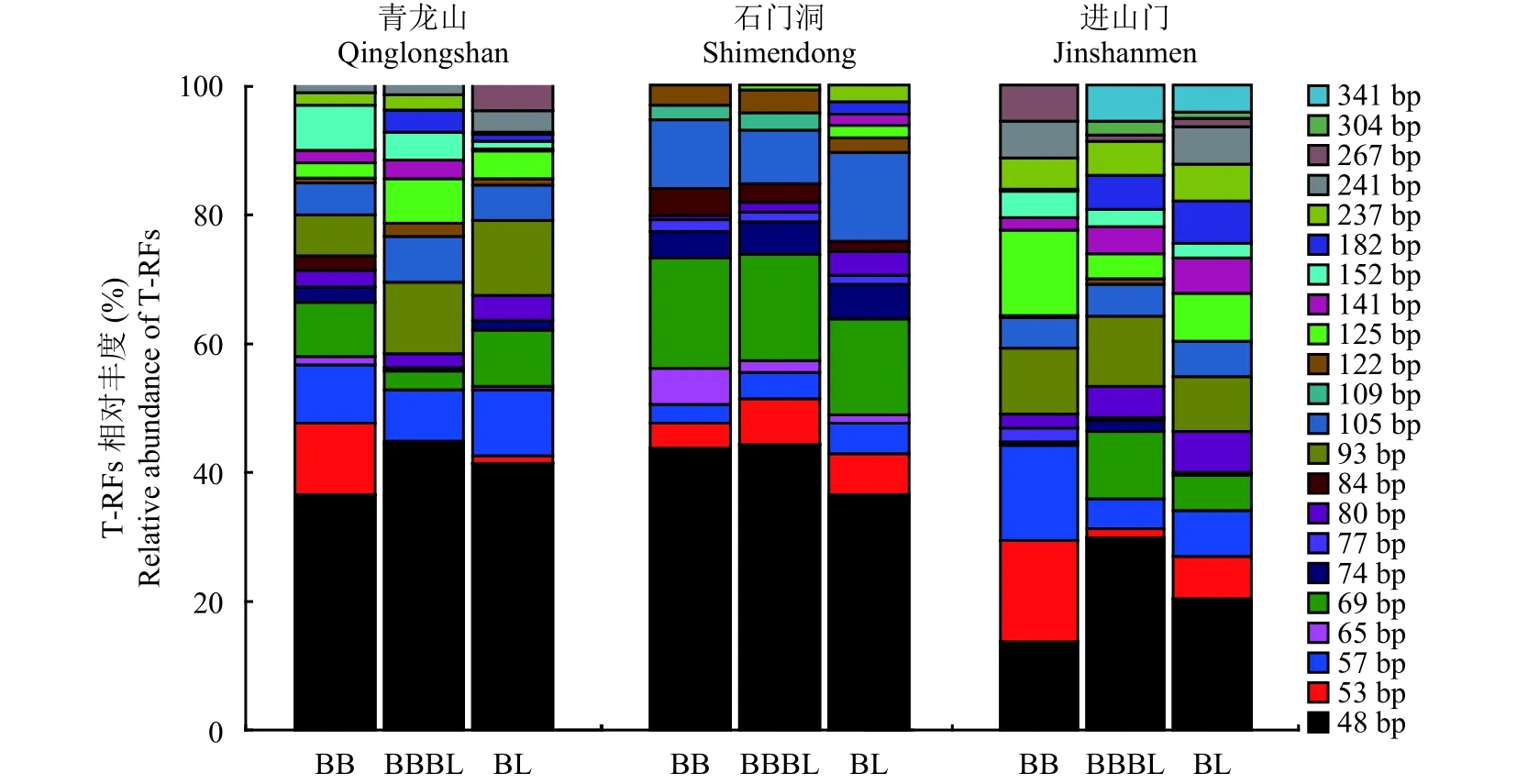
图1 三个入侵带土壤nifH基因T-RFs (bp) 相对丰度Fig.1 Relative abundance of T-RFs (bp) of soil nifH genes in the three invasion belts[注(Note):BB—毛竹林 Bamboo forest; BBBL—毛竹阔叶混交林 Mixed forest of bamboo and broadleaf or needle;BL—阔叶林 Broadleaf or needle forest.]
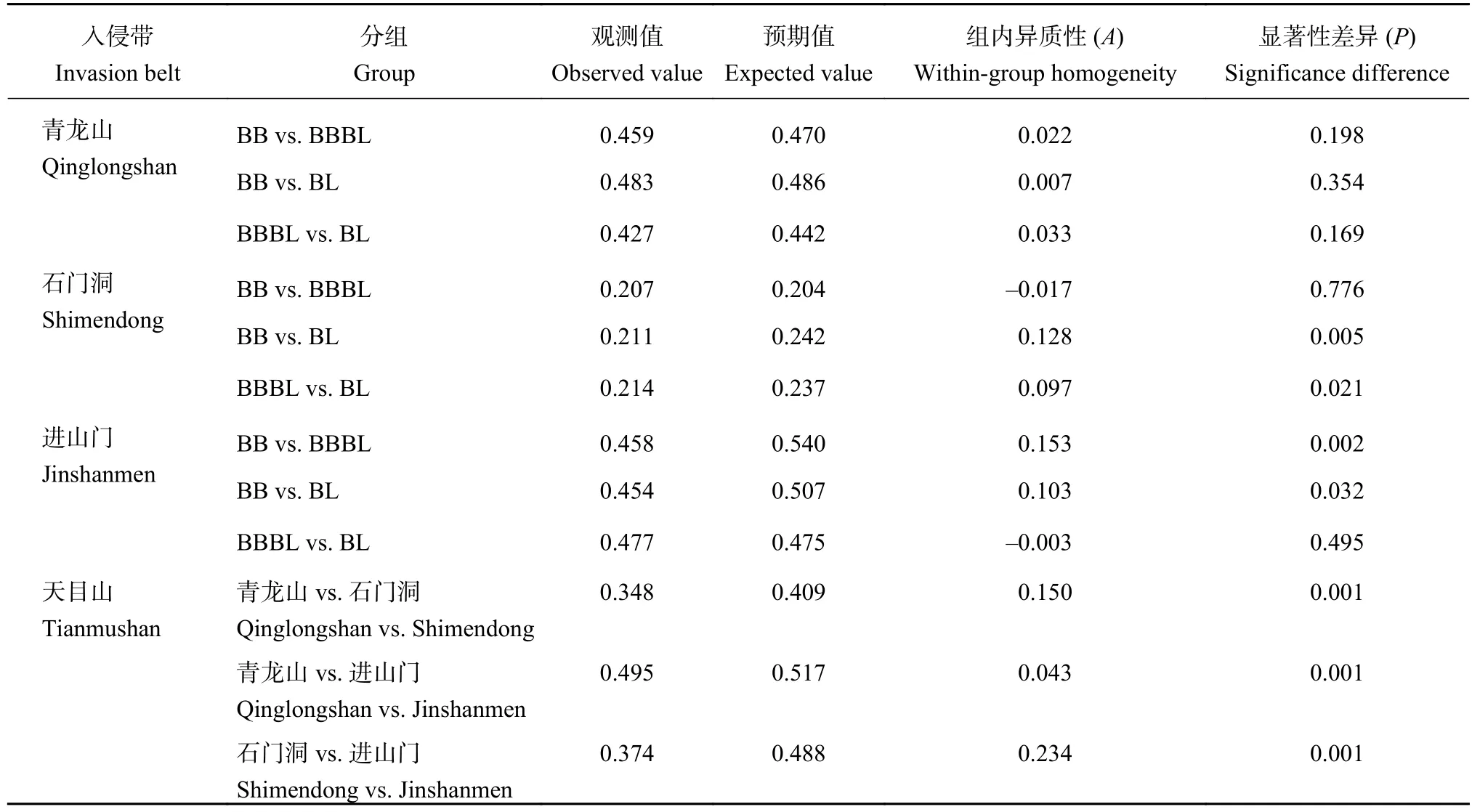
表2 三个入侵带不同林分土壤固氮微生物群落结构的多响应置换过程分析Table2 Multi-response permutation procedure analysis of soil nitrogen-fixing microbes community structure between vegetation and invasion belts
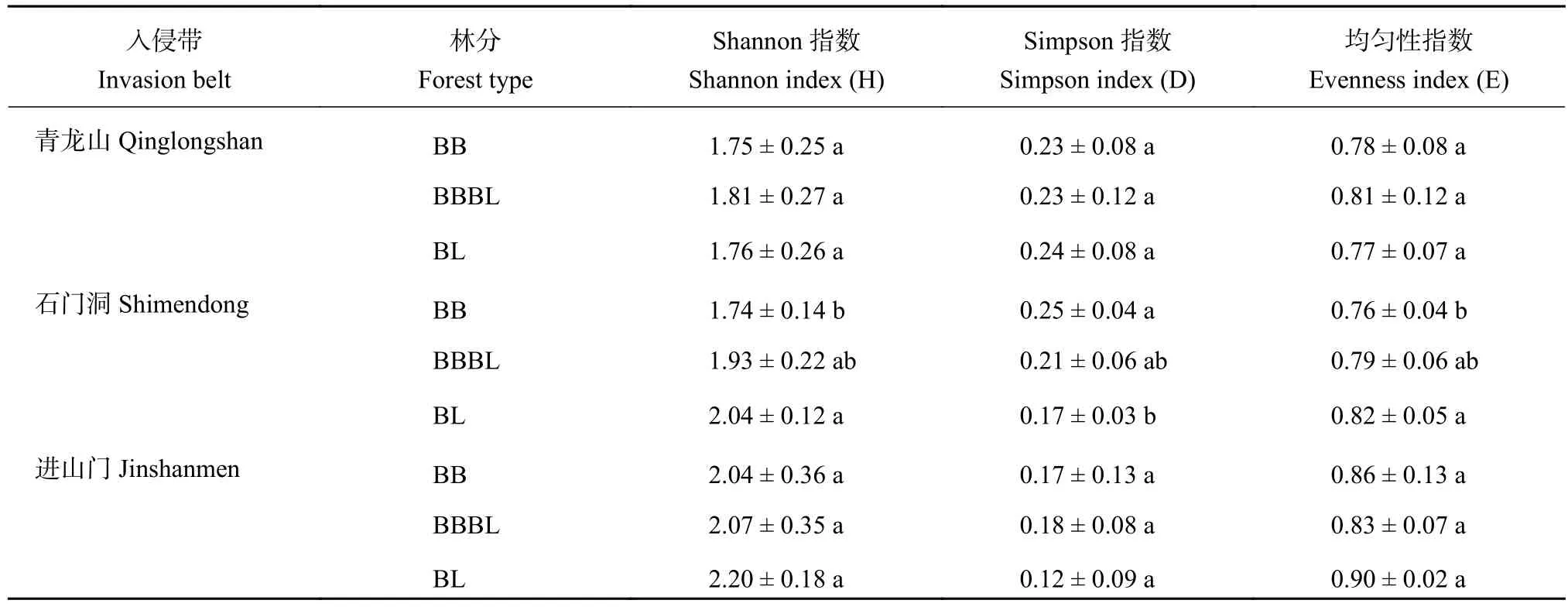
表3 三个入侵带土壤nifH基因多样性指数Table3 Biodiversity indices of soil nitrogen-fixing microbes community in the three invasion belts
如图2显示,青龙山、石门洞和进山门入侵带的土壤 nifH 基因拷贝数分别为 2.68 × 107~3.48 ×107、3.59 × 107~4.52 × 107和 4.45 × 107~11.9 × 107copies/g干土,进山门入侵带土壤nifH基因丰度高于其他两个。随着毛竹入侵,土壤nifH基因丰度均呈下降趋势 (BB<BBBL<BL),但显著差异只存在于进山门的毛竹林与混交林之间 (P<0.05),而青龙山和石门洞的nifH基因丰度在三种林分中均无显著性差异。
2.5 土壤固氮微生物与土壤化学性质的关系
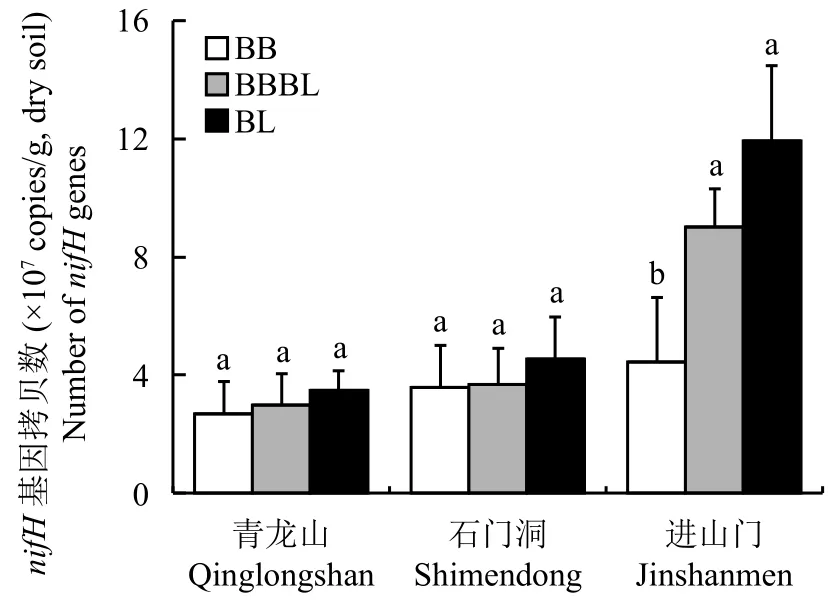
图2 三个入侵带土壤nifH基因拷贝数Fig.2 Soil nifH gene copies in three invasion belts[注(Note):BB—毛竹林 Bamboo forest; BBBL—毛竹阔叶混交林 Mixed forest of bamboo and broadleaf or needle; BL—阔叶林Broadleaf or needle forest.方柱上不同字母表示同一入侵带不同林分间差异显著 (P<0.05) Different letters above the bars indicate significant differences among different forest types in the same invasion belt (P<0.05).]
将三个入侵带土壤固氮微生物的nifH基因丰度、多样性指数分别与土壤化学性质作相关性分析,表4列出了有显著性差异的部分,青龙山入侵带的土壤固氮微生物的各项指标中,仅均匀度指数与土壤硝态氮含量有显著的相关性;石门洞的三个多样性指数均与土壤有机碳含量有显著相关性,与有效磷含量有极显著相关性;进山门仅nifH基因丰度与有效磷含量有显著相关性,与速效钾含量有极显著相关性。
将三个入侵带土壤固氮微生物群落结构与土壤化学性质分别和综合进行冗余分析,结果如图3所示,土壤化学性质对青龙山和进山门 (图3a、图3c)两个入侵带土壤固氮微生物群落结构变异的影响均不显著,且不同林分之间未形成明显的聚类差异;石门洞入侵带中 (图 3b),速效钾 (F = 3.963,P =0.003) 和铵态氮 (F = 2.248,P = 0.049) 含量显著影响该入侵带土壤固氮微生物群落结构的变异;合并三个入侵带的土壤固氮微生物群落结构信息和土壤化学性质的RDA结果表明 (图3d),有三个土壤性质达到显著水平,分别是碱解氮 (F = 3.016,P = 0.004)、有效磷 (F = 2.671,P = 0.024) 和速效钾 (F = 2.838,P = 0.005) 含量,说明它们是毛竹入侵过程中土壤总固氮微生物群落结构变异的主要影响因素。综合以上结果,可以看出不同入侵带之间没有统一的规律性分布格局,可能与入侵带的土壤性质、植被类型等不同有关。
3 讨论与结论
土壤作为植物生长的介质,能够提供植物生长繁殖所需的水分和养分[30],反之,植物也能够对土壤的性质产生影响[31]。植物入侵改变了植被组成,不同植物对土壤养分的差异吸收,以及林地的凋落物质[32]、根系分泌物[33]等随之发生改变,从而改变土壤的性质。本研究结果表明,在毛竹入侵周边阔叶林以及针阔混交林过程中,三个入侵带土壤的pH值均有上升趋势,而速效钾含量均有所下降,这与已有的研究结果一致[16–17,20],其中,毛竹对于土壤钾素的偏好吸收可能是引起速效钾下降的原因[34],而不同营养元素以及根系分泌物组成的改变可能引起了土壤pH值的上升。在毛竹入侵过程中,虽然多种类的阔叶树或针叶树被单一的毛竹所替代,但毛竹具有较强的固碳能力[4]以及拥有大量的地下凋落物[35],因此出现本文有机碳量随毛竹入侵过程增加的现象。无机氮成分—硝态氮和铵态氮之间含量相近,铵态氮有显著上升的情况,这与宋庆妮等[36]毛竹扩张对无机氮影响较小,且铵态氮是主要组分的研究结果有所差异,可能与研究区域本身氮素含量不同有关。三个入侵带的有效磷含量在毛竹入侵后有上升的现象,可能与pH值的上升有关,磷素在酸性土壤尤其是pH<5的环境中,容易形成Fe-P、Al-P等难溶性物质[37],降低土壤中有效磷含量。总体上,各个入侵带的土壤养分含量在毛竹入侵后有所上升,说明毛竹入侵并不一定只单纯地对生态系统造成危害。

表4 三个入侵带土壤nifH基因丰度和多样性指数与土壤化学性质的相关性分析Table4 Correlation analysis between nifH gene abundance, diversity indices and soil chemical properties in the three invasion belts

图3 土壤固氮微生物群落结构与土壤化学性质的冗余分析Fig.3 RDA of soil nitrogen-fixing microbes community and soil chemical properties[注(Note):BB—毛竹林 Bamboo forest;BBBL—毛竹原林混交林 Mixed forest of bamboo and broadleaf or needle;BL—原林地 Broadleaf or needle forest;SOC—土壤有机碳 Soil organic carbon;AN—碱解氮 Available nitrogen;AP—有效磷 Available phosphorus;AK—速效钾Available potassium.]
土壤为微生物生长提供主要的养分和能量,土壤环境的变化导致微生物生物量的变化[38],而不同植物类型、生长阶段、基因型以及植物多样性等则可直接影响或通过改变土壤环境间接影响土壤微生物的群落结构、多样性和活性[39]。植物入侵过程中植物的组成及多样性均发生变化,戴莲等[40]发现紫茎泽兰的入侵提高了土壤中固氮菌和氨氧化细菌等有益功能细菌的种群数量,Xu等[17]发现毛竹入侵天然阔叶林地后土壤微生物生物量和细菌多样性上升,但利用碳源的功能多样性没有发生变化。本研究结果发现,毛竹入侵原有林分后土壤固氮微生物功能基因丰度呈下降趋势,而土壤固氮微生物的丰富度、优势度和均匀度仅在石门洞有显著变化。对固氮微生物来说,固氮过程会消耗大量用于生长的能源物质[41],偏好养分充足的土壤[42],一般认为固氮微生物数量与有机质含量呈正相关[43],虽然土壤氮素含量与有机质含量呈正相关,但土壤氮素含量并不与固氮微生物数量有正相关性,因为其中高浓度的无机态氮素对固氮菌生长反而具有抑制性[44]。毛竹替代原林后固氮微生物丰度下降,可能是毛竹林有效氮 (碱解氮和铵态氮) 增加对土壤固氮细菌生长起到抑制作用,而土壤速效钾下降可能是另一诱因。土壤固氮微生物nifH丰度与土壤化学性质之间的显著相关性只发生在进山门入侵带 (nifH基因丰度分别与有效磷显著负相关、与速效钾极显著正相关) 的结果说明,固氮微生物与土壤性质的关系比较复杂。毕江涛等[39]指出,土壤微生物多样性与植物多样性呈正相关,毛竹入侵后改变了植物多样性,尤其是乔木层物种多样性降低[15],使得一部分固氮微生物种类消失 (土壤固氮微生物丰富度下降,表3),这可能是导致固氮微生物丰度下降的原因。同时,阔叶树或者针阔混交林被单一的毛竹取代后,嗜好毛竹凋落物和根系分泌物的固氮微生物种类增加,可解释毛竹入侵后土壤固氮微生物丰富度下降和优势度上升的现象,这与王纪杰[45]研究发现竹叶加入到阔叶林土壤中,导致微生物利用碳源的功能多样性指数随时间延长下降,微生物群落逐渐简单化的结果类似。本研究中毛竹入侵取代原树种形成毛竹纯林,与沈秋兰等[46]的阔叶林改种为毛竹纯林情况相似,两个研究中阔叶林与毛竹林土壤pH和有机碳之间的差异类似,但土壤固氮细菌丰度变化与本研究结果相反。两个研究中阔叶林与毛竹林之间土壤速效钾和有效磷的变化正好相反,可能是引起结果不同的原因。从表4三个入侵带中土壤性质与功能基因丰度及固氮微生物多样性的相关关系结果完全不一致,可以证明毛竹入侵过程中影响固氮微生物的因素比较复杂。这一结论可以从RDA结果得到进一步证明,三个入侵带土壤固氮微生物群落结构与土壤化学性质关系各不相同,仅石门洞入侵带有显著影响因子 (铵态氮和速效钾),并且土壤化学性质对这三个入侵带的土壤固氮微生物群落结构变化的解释度较低,说明在本研究的三个入侵带中,土壤性质对土壤固氮微生物群落结构影响度不高,还需结合其他因素进行探讨。
从T-RFLP图谱的片段组成看 (图1),进山门入侵带与其他两个入侵带差异明显,可能因为进山门土壤母质主要为石英斑岩,属于花岗岩类,而其他两地主要为泥质灰岩,属于碳酸盐岩类,两者在化学性质上并不相同[47],导致土壤的化学性质存在差异(表1)。三个入侵带的共同优势片段48 bp在进山门入侵带中相对比例明显较低,而毛竹入侵后变化却很大。董志新等[48]发现农田的不同土壤类型是影响土壤固氮微生物群落结构的显著影响因子。MRPP分析表明,毛竹入侵形成的三个林分之间土壤固氮微生物群落结构变化没有一致规律,石门洞和进山门入侵带不同林分之间的土壤固氮微生物群落结构有显著变异,而青龙山入侵带则变异不显著。张于光等[49]研究发现高寒草甸、高寒草原和高寒森林的土壤固氮微生物群落结构具有明显的差异,而文都日乐等[50]研究草原土壤固氮微生物时,发现土壤固氮微生物的群落结构在不同草地类型间的差异同样显著,说明植被是影响土壤固氮微生物群落结构的重要因素。青龙山毛竹入侵前后差异不显著,可能与其原有林地是针阔混交林有关,而石门洞和进山门入侵带是阔叶林。三个入侵带土壤固氮微生物的共同MRPP分析进一步证明,位于同一自然保护区的三个入侵带的土壤微生物之间有着极显著差异。概括分析得知,同为泥质灰岩的石门洞和青龙山入侵带,石门洞毛竹入侵后土壤固氮微生物群落结构有显著变异,而青龙山则变化不明显;发育于石英斑岩的进山门入侵带土壤固氮微生物群落结构与石门洞和青龙山入侵带明显不同、入侵过程中土壤固氮微生物群落结构变化明显;以上结果说明,天目山毛竹入侵过程中土壤固氮微生物群落结构的变化是土壤性质 (取决于母岩) 和植被交互作用的结果。
以上研究表明,即使同一自然保护区的三个毛竹入侵带,显著影响土壤固氮微生物群落变化的土壤化学性质并不相同。土壤母质和原有植被组成的综合作用导致土壤固氮微生物群落发生变化。因此,毛竹入侵周边林地导致的土壤化学性质以及固氮微生物的变化机制比较复杂,需要多层次、多尺度地来探讨毛竹入侵对土壤固氮微生物的影响。
猜你喜欢
农业科技与信息(2021年16期)2021-12-07
生态科学(2021年5期)2021-11-13
今日中国·西班牙文版(2021年4期)2021-04-09
安徽农学通报(2021年1期)2021-02-07
绿色中国(2019年18期)2020-01-04
科学与技术(2019年21期)2019-04-27
现代园艺(2018年2期)2018-03-15
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
自然保护地(2016年1期)2016-04-22
