
水分胁迫下马铃薯叶片光合特性的变化及其响应机制研究
2018-08-10 07:14:22刘素军蒙美莲陈有君焦瑞枣
西北农林科技大学学报(自然科学版) 2018年8期
刘素军,蒙美莲,陈有君,焦瑞枣
(内蒙古农业大学 a农学院,b生命科学学院,内蒙古 呼和浩特 010019)
我国是世界上13个贫水国家之一,人均水资源占有量少,不到世界平均水平的1/4,缺水严重[1],而且由于我国连续遭受旱灾,致使水资源短缺不断加剧。土壤水分是影响植物生理生态特性及生长发育的重要生态因子之一。水分胁迫已成为制约植物生长发育的主要逆境因子之一[2],直接影响着植株的光合性能[3-5]。光合作用是作物生长发育和产量形成的基础,作物95%以上的干物质由光合作用提供[6]。前人研究表明,水分胁迫下玉米叶片的光合特性下降,光合速率日变化的峰值提前,影响了玉米叶片的正常光合作用[7]。干旱胁迫下糜子气孔导度及胞间CO2浓度下降,叶片净光合速率降低,生物量减少,抗旱性强的糜子品种具有较高的电子传递速率和光能转化能力[8]。马铃薯对水分亏缺非常敏感,水资源短缺极大地影响了马铃薯生产。目前有关水分胁迫对马铃薯形态指标、抗氧化酶、渗透调节物质影响的研究较多[9-11]。研究表明,水分胁迫下马铃薯不同品种光合特性存在差异,随着胁迫程度的增加光合特性指标降低,从而影响了产量[12]。在分子水平上,前人对马铃薯差异蛋白[13]、马铃薯SnRK2基因[14]等方面进行了初步研究。但对水分胁迫和复水条件下马铃薯叶片叶绿素相对含量(SPAD值)和光合特性变化规律、光合特性相关差异基因的研究尚未见报道。为此,本研究以“克新1号”马铃薯为材料,研究马铃薯关键生育期内水分胁迫和复水对SPAD值、光合特性的影响,最后在室内利用高通量测序技术对水分胁迫后马铃薯的差异基因及代谢途径进行分析,拟揭示水分胁迫和复水条件下马铃薯光合特性的响应机制,以期为提高马铃薯抗旱性提供科学依据。
1 材料与方法
1.1 试验材料
田间试验材料为“克新1号”原种脱毒种薯,室内试验材料为“克新1号”原原种脱毒种薯,这2种材料均由内蒙古民丰薯业公司提供。
1.2 试验设计及方法
1.2.1 田间试验设计 田间试验在内蒙古武川县大豆铺室外遮雨棚进行,试验地土壤类型为壤土,0~20 cm土壤碱解氮含量41.68 mg/kg,有效磷含量11.23 mg/kg,速效钾含量104.59 mg/kg。采用膜下滴灌的种植方式,宽窄行种植,宽行行距0.7 m,窄行行距0.4 m,株距0.33 m,密度6 万株/hm2,小区面积5.5 m×4 m=22 m2。播种前撒施羊粪4 500 kg/hm2,氮磷钾复合肥(m(N)∶m(P2O5)∶m(K2O)=13∶17∶15)750 kg/hm2,人工起垄后,2014-05-16人工点播播种。试验在马铃薯块茎形成期进行。设置土壤相对含水量分别为40%(为田间持水率的40%,下同,T1)、55%(T2)、70%(T3)、85%(T4)和100%(T5)5个处理,随机区组设计,每个处理4次重复,为防止各小区间水分互渗,影响控水效果,各小区均间隔0.5 m。播种后各处理土壤相对含水量统一控制在75%左右,从马铃薯块茎形成期(7月1日)开始控水,连续处理21 d,水分胁迫处理结束后,进行复水处理,保持各处理的土壤含水量均在80%以上直至块茎膨大期结束,淀粉积累期土壤含水量保持在65%以上,之后按常规管理。其中T5处理在水分胁迫处理结束后停止供水,待土壤相对含水量自然降至80%后开始与其他处理一样控水。土壤相对含水量用TDR水分测定仪测定,每隔3 d测定1次,当土壤相对含水量低于设定值时及时进行灌水,灌水量由水表控制。
1.2.2 室内试验设计 试验采用盆栽法,选择高50 cm、最大口径30 cm的塑料桶,内装过筛沙土4 kg,土壤取自内蒙古农业大学农场。选取大小均一的种薯播种,每盆播种3个马铃薯种薯,每个种薯质量5~7 g,播种后保持土壤相对含水量70%左右。试验设干旱处理(土壤相对含水量为40%,DT)和对照(保持土壤相对含水量为70%,CK),每处理设10个重复。室内温度保持在(23±2) ℃,平均相对湿度为50%,出苗后株高约5 cm时开始进行水分胁迫,每天09:00-10:00固定时间采用称重法控水,以确保各处理的土壤含水量。
1.3 测定项目及方法
水分胁迫7 d后取样,每隔7 d取1次样,共取样3次;从复水7 d后取样,每隔7 d取1次样,共取样3次。
1.3.1 SPAD值 采用日产SPAD-502型手持便携式叶绿素仪,于从上往下数第2片完全展开叶最宽处测定SPAD值,每小区选取4株进行测量,每片叶子重复测量3次,取其平均值表示该叶片叶绿素相对含量。
1.3.2 光合特性 每个小区选择位于小区中间有代表性且生长健康、长势一致的植株3株,使用Li-6400型光合仪在晴天的09:00-11:00,测定从上往下数第2片完全展开叶的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)。测定叶室面积为6 cm2,光照强度(PAR)为1 500 μmol/(m2·s),叶室CO2含量为500 μL/L,温度为(25±2) ℃。
1.3.3 产 量 田间试验期间预留完整2行用来测产,其占小区面积的20%。首先统计株数,然后全部挖出并分级,分别记录其数量后称质量,计算单株平均产量,根据其密度再计算出每公顷产量。
1.3.4 基因表达 水分胁迫14 d后,分别在干旱处理和对照的30个单株上,采集马铃薯幼嫩叶片并均匀混合,用液氮速冻,置于-80 ℃冰箱中储存备用。委托北京诺禾致源生物信息科技有限公司转录组测定,并利用KOBAS(2.0)软件对差异表达基因显著富集的KEGG(kyoto encyclopedia of genes and genomes)通路进行分析。
1.4 数据处理与分析
采用Excel 2010进行试验数据处理,方差分析和多重比较采用SPSS 16.0最小显著性差异(LSD)法,线性模拟斜率采用Rv 3.2.2软件进行协方差分析[15]。
2 结果与分析
2.1 水分胁迫及复水对马铃薯叶片叶绿素相对含量的影响
由图1可以看出,块茎形成期进行水分胁迫后,各处理叶片SPAD值均呈下降趋势。水分胁迫第7天时,T4处理叶片SPAD值显著高于其他处理,而T1、T2、T3处理和T3、T5处理之间无显著差异;水分胁迫第14天时,T1处理叶片SPAD值显著低于其他处理;水分胁迫第21天时,除了T2和T5处理外,其余处理之间差异均显著,表明水分胁迫初期对SPAD抑制不明显,但是随着水分胁迫时间的延长,抑制作用逐渐明显。复水处理后,T1和T2处理叶片SPAD仍呈下降趋势,但下降幅度变缓,T3处理叶片SPAD则快速恢复到T4处理水平,在复水21 d时,T1、T2、T3、T5处理的叶片SPAD值分别达到T4处理的33.5%,40.5%,97.5%,97.0%,T3、T4和T5处理之间差异不显著,但显著高于T1、T2处理,T1与T2处理之间有显著差异。
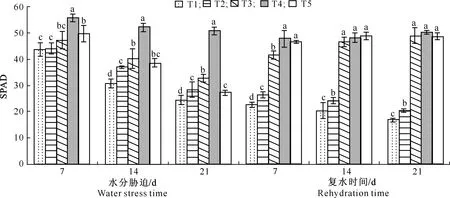
图柱上标不同小写字母表示在P<0.05水平上差异显著。下图同 Different small letters indicate significant difference at P<0.05.The same below图1 水分胁迫及复水对马铃薯叶片叶绿素相对含量的影响Fig.1 Effects of water stress and rehydration on SPAD of potato leaves
2.2 水分胁迫及复水对马铃薯光合特性的影响
水分胁迫及复水对马铃薯光合特性的影响如图2~5所示。
2.2.1 净光合速率 由图2可以看出,随着水分胁迫时间的延长,马铃薯叶片净光合速率(Pn)呈下降趋势;随着水分胁迫强度的增加,Pn呈先增后减的趋势。
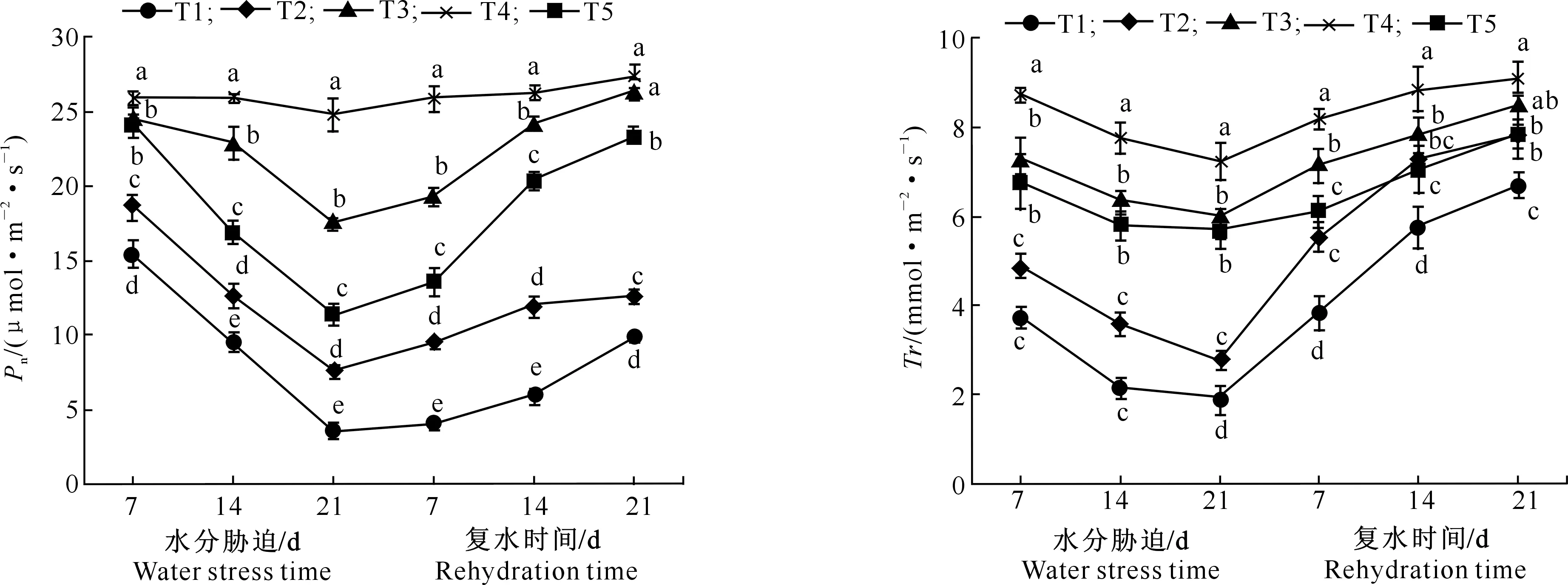
图2 水分胁迫及复水对马铃薯叶片Pn的影响 图3 水分胁迫及复水对马铃薯叶片Tr的影响Fig.2 Effects of water stress and rehydration on net photosynthetic rate of potato leaves Fig.3 Effects of water stress and rehydration on transpiration rate of potato leaves
图2显示,水分胁迫7 d 时,除T3与T5处理间Pn无显著差异外,其他处理间差异均达到显著水平;水分胁迫14 d时,各处理之间Pn差异均已达到显著水平,具体表现为T4>T3>T5>T2>T1,说明水分胁迫初期对Pn的影响较大,且水分胁迫时间越长影响越明显。复水后,各处理马铃薯叶片Pn逐步恢复,其中T3处理Pn的恢复程度较大,T1、T2处理Pn虽然有所恢复,但仍显著低于T3、T4处理,T5处理Pn恢复程度也显著低于T3、T4处理,但显著高于T1、T2处理,表明当土壤相对含水量低于55%时,连续水分胁迫或过度饱和对马铃薯造成了一定的损伤,复水后虽然恢复了一部分功能,但无法完全恢复。
2.2.2 蒸腾速率 从图3可以看出,随着水分胁迫时间的延长,T3、T4、T5处理马铃薯叶片蒸腾速率(Tr)下降趋势较缓慢,T1、T2处理下降幅度较大。水分胁迫期间,T4处理的叶片Tr显著高于其他处理,T3与T5处理之间无显著差异,水分胁迫第7天和14天时,T1与T2处理之间无显著差异;至水分胁迫第21天时,T1与T2处理之间差异显著,各处理Tr表现为T4>T3>T5>T2>T1。复水后第21天,T3与T4处理之间Tr无显著差异,说明T3、 T4处理Tr未受到明显影响,而T1处理的Tr上升幅度较大,但仍显著低于其他处理,表明当土壤相对含水量低于40%时,其对马铃薯叶片结构造成一定的损伤,且无法恢复。
2.2.3 气孔导度 由图4可以看出,随着水分胁迫时间的延长,气孔导度(Gs)呈下降趋势,T1、T2处理Gs下降幅度较大,T3、T4和T5处理下降幅度较小。水分胁迫7 d时,各处理Gs差异显著,说明在遭受水分胁迫时,马铃薯通过迅速降低叶片Gs、减少水分蒸发来适应水分胁迫。水分胁迫21 d时,各处理Gs表现为T4>T3>T5>T2>T1,T2与T1处理之间差异不显著,其他处理之间差异显著。复水后,T4处理的Gs无明显变化,T1、T2、T3、T5处理的Gs均呈现增长趋势,其中T3处理Gs在复水14 d时几乎恢复到与T4处理相当的水平,二者之间无显著差异,而T1、T2处理显著低于T3、T4处理,表明T1、T2处理的叶片气孔结构受到损伤,无法恢复。
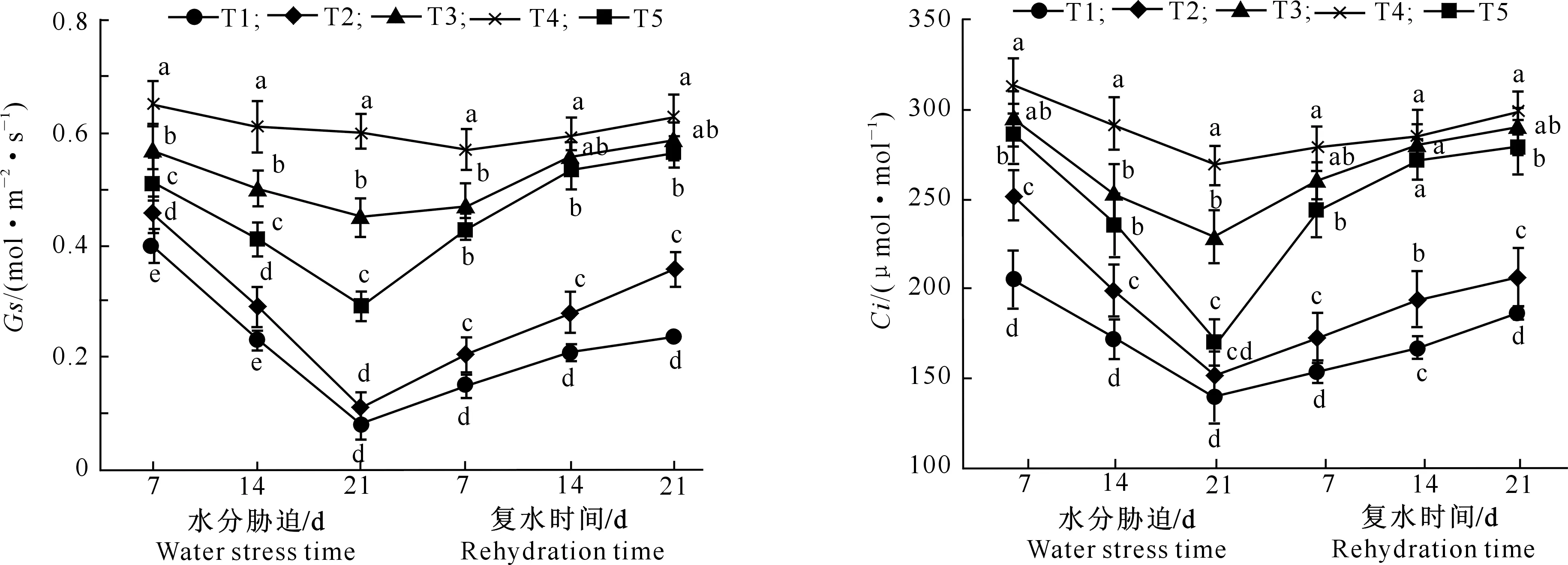
图4 水分胁迫及复水对马铃薯叶片Gs的影响 图5 水分胁迫及复水对马铃薯叶片Ci的影响Fig.4 Effects of water stress and rehydration on stomatal conductance of potato leaves Fig.5 Effects of water stress and rehydration on intercellular CO2 concentration of potato leaves
2.2.4 胞间CO2浓度 由图5可以看出,水分胁迫和复水期间,各处理马铃薯叶片胞间CO2浓度(Ci)均呈“V”型变化,即水分胁迫期间各处理Ci下降,复水后呈增长趋势。水分胁迫第21天时,除T1与T2、T2与T5处理之间Ci无显著差异外,其余处理之间差异显著。复水后,T3处理Ci几乎恢复到T4处理水平,T1、T2处理Ci虽然有所回升,但是仍显著低于T3、T4、T5处理,这可能是因为受到水分胁迫对Gs的影响,T1、T2处理Ci也无法恢复。
2.3 水分胁迫对马铃薯产量的影响
由图6可以看出,随着水分胁迫程度的加深,马铃薯产量先增加后下降,其中T4处理马铃薯产量最高;其次是T3和T5处理,且二者差异不显著;产量最低的是T1处理,表明水分胁迫对马铃薯产量有显著影响。

图6 水分胁迫对马铃薯产量的影响Fig.6 Effects of water stress on potato yield
2.4 水分胁迫时间与马铃薯叶绿素含量、光合特性线性关系的模拟
从胁迫时间对叶绿素含量、光合特性的线性模拟结果来看,除T4、T5处理下Tr和T4处理下Gs以外,胁迫时间与SPAD值、光合特性之间均有显著线性关系,并随着水分胁迫时间的延长,各指标均呈下降趋势(表1),直线模拟后,不同处理各指标斜率的差异显著性不同(图7~9)。

表1 水分胁迫时间与马铃薯各项生理指标的线性模拟结果Table 1 Linear regressions between stress time and each index under different treatments
注:y代表各项指标,x代表水分胁迫时间,R代表相关系数,*代表在P<0.05水平上显著相关,**代表在P<0.01水平上显著相关。
Notes:“y” indicates each index,“x” indicate water stress time,“R” indicates correlation coefficient,“*” indicates significant atP<0.05 level,and “**” indicates significant atP<0.01 level.
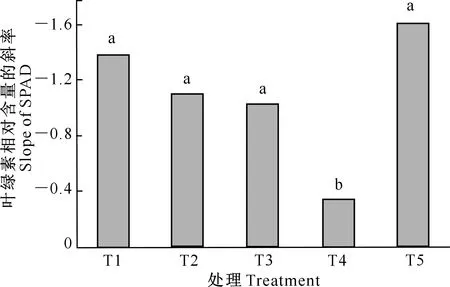
图7 不同处理水分胁迫时间与马铃薯叶片叶绿素相对含量线性模拟斜率的比较Fig.7 Linear regressions between water stress time and SPAD of photo leaves under different treatments
不同处理水分胁迫时间与马铃薯叶片叶绿素相对含量线性模拟斜率的比较结果如图7所示。从图7可以看出,T1、T2 、T3和T5处理之间的SPAD斜率无显著差异,但均显著高于T4处理。
从图8和图9可以看出,T1、T2和T5处理之间的Pn斜率无显著差异,但均显著高于T3处理,T3处理的Pn斜率又显著高于T4处理。T1与T2、T3与T4处理之间的Gs斜率无显著差异,T1、T2处理的Gs斜率显著高于T5处理, T5处理又显著高于T3和T4处理。T2和T5处理的Ci斜率显著高于T1、T3、T4处理。各处理之间Tr斜率无显著差异,说明时间梯度上Tr变化幅度不大,而SPAD值、Pn、Gs和Ci的变化幅度较明显。

图9 不同处理水分胁迫时间与马铃薯叶片气孔导度和胞间CO2浓度线性模拟斜率的比较Fig.9 Linear regressions between water stress time and potato stomatal conductance and intercellular CO2 concentration under different treatments
2.5 水分胁迫对马铃薯KEGG代谢通路和叶片光合特性基因表达的影响
2.5.1 马铃薯KEGG代谢通路 遭受水分胁迫后,马铃薯很多代谢途径中酶的合成由于基因表达的变化而受到影响。由表2可以看出,与对照相比,水分胁迫后,马铃薯KEGG代谢通路差异显著的途径有6条,分别为类黄酮合成、二芳基庚和姜辣素的生物合成、甘油酯代谢、糖脂生物合成、光合作用-天线蛋白和维他命B6代谢,表明干旱胁迫对光合作用的影响较为显著。

表2 水分胁迫下马铃薯代谢差异显著的KEGG通路Table 2 KEGG pathway with significant difference under water stress
2.5.2 叶片光合特性基因表达 本研究发现,马铃薯苗期水分胁迫14 d后,KEGG通路中光合作用-天线蛋白通路显著性富集(P<0.05),其中通路中捕光复合体Ⅰ的叶绿素a/b结合蛋白1(light-harvesting complex Ⅰ chlorophyll a/b binding protein 1,Lhca 1)、叶绿素a/b结合蛋白4(Lhca 4)的PGS-C0003DMG400023344、PGSC0003DMG400023461、PGSC0003DMG400019508基因表达显著上调,捕光复合体Ⅱ的叶绿素a/b结合蛋白1(light-harvesting complex Ⅱ chlorophylla/ b binding protein 1,Lhcb 1)、叶绿素a/b结合蛋白3(Lhcb 3)的PGSC0003DMG400013460、PGSC0003DMG400008564基因表达显著上调(表3)。可知当马铃薯遭受水分胁迫后,捕光复合体Ⅰ和Ⅱ蛋白相关的基因表达显著上调,说明马铃薯通过吸光天线蛋白系统来调节缺乏水分的应急情况,即天线蛋白系统向相连的反应中心有效地输送能量,从而适应缺水环境。
KEEG通路中,植物病原体相互作用(plant-pathogen interaction)通路虽然没有显著性富集(P=0.64>0.05),但是相关基因中,环核苷酸门控离子通道(cyclic nucleotide-gated ion channels,CNGCs)相关的PGSC0003DMG402017989基因和钙调素(calmodulin,CaM)、钙调素类蛋白(calcium-binding protein,CML)相关的PGSC0003DMG400-019527基因表达显著下调(表3)。当马铃薯遇到水分胁迫后,通过环核苷酸门控离子通道,使调节Ca2+稳态的基因表达量显著下调,减少细胞质中Ca2+含量,与CaM、CML相关的基因表达量也显著下调,从而使一氧化氮合酶活性受影响,最终导致气孔关闭。
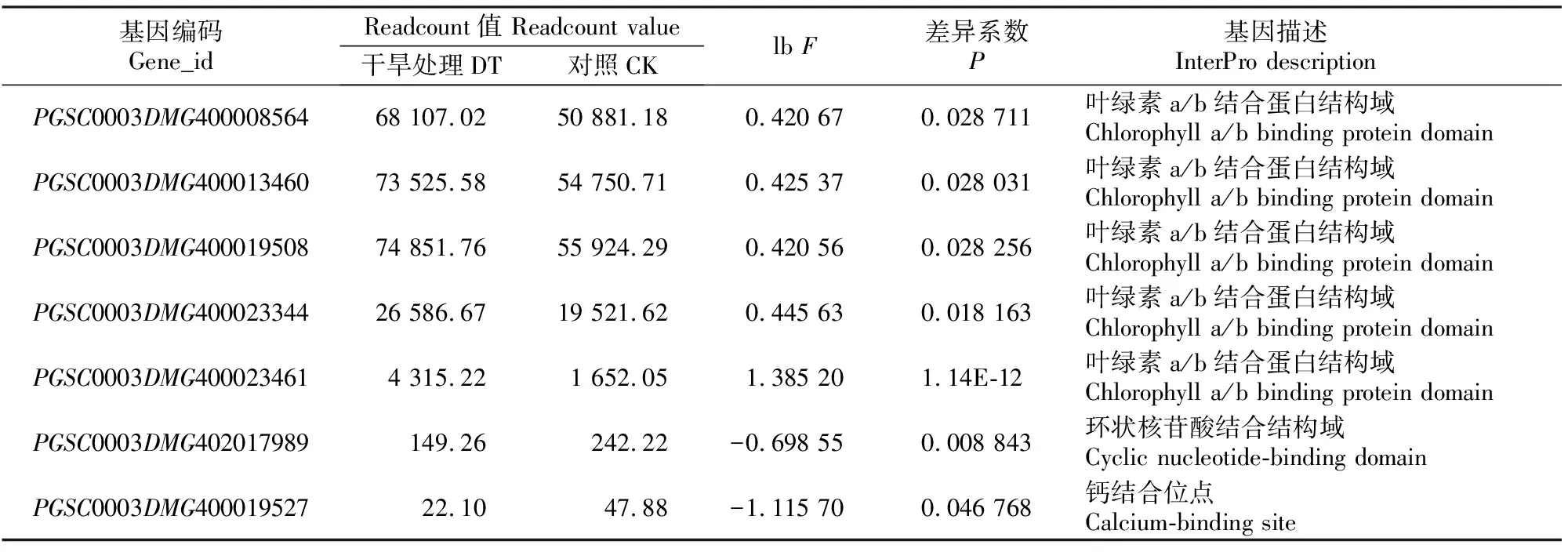
表3 水分胁迫下马铃薯光合作用-天线蛋白及植物病原体相互作用通路中的差异表达基因Table 3 Differential gene list in photosynthesis-antenna proteins plant-pathogen interaction pathways of potato under water stress
注:lbF=lb (干旱处理与对照 Readcount值的比值)。lbF>0表明基因呈上调表达,lbF<0表明基因呈下调表达。
Note:lbF=lb (DT/CK Readcount value). lbF>0 means genes were up-regulated,while lbF<0 means genes were down-regulated.
3 讨 论
3.1 水分胁迫下马铃薯各项指标的变化趋势
本研究中,随着水分胁迫时间的延长,马铃薯叶片SPAD值、Pn、Tr、Gs和Ci均下降;随着水分胁迫程度的加剧,马铃薯叶片SPAD值、Pn、Tr、Gs、Ci和产量先增加后下降,这与冯晓敏等[8]、王婷等[12]的研究结果一致。整个水分胁迫期间,T4处理的SPAD值和光合特性表现最稳定,产量最高,说明马铃薯块茎形成期适宜的土壤相对含水量为85%;其他处理马铃薯均受到不同程度的水分胁迫,导致马铃薯光合生理特性产生抑制作用,从而影响了产量。
3.2 复水后马铃薯各项指标的变化趋势
本研究中,随着复水时间的延长,马铃薯叶片SPAD值、Pn、Tr、Gs、Ci增加,但不同处理各指标的恢复速度和恢复程度不同。复水21 d后,T3与T4处理上述指标之间无显著差异,说明T3处理SPAD值、Pn、Tr、Gs、Ci能恢复到与适宜含水量处理(T4)相当的水平,说明土壤相对含水量为70%的水分胁迫处理复水后各项光合特性功能在一定程度上能恢复。复水后,T1和T2处理的SPAD值仍呈下降趋势,表明马铃薯叶片在遭受中度水分胁迫(土壤相对含水量为55%)或重度水分胁迫(土壤相对含水量为40%)时导致马铃薯叶片损伤较重,叶绿体超微结构受损,在试验期间即使复水也无法修复,这与赵天宏等[16]对玉米的研究结论一致。与T3处理相比,T1、T2处理SPAD值和光合特性恢复较慢,而且显著低于T3处理,表明土壤相对含水量为40%~55%的连续水分胁迫对马铃薯光合系统产生了危害,复水后虽然恢复了一部分功能,但无法完全恢复。T3处理SPAD值和光合指标值均高于T5处理,但二者只有Pn差异显著,SPAD值、Tr、Gs、Ci无显著差异,最终二者产量之间也无显著差异,说明T5处理Pn恢复需要更长时间。
3.3 水分胁迫时间与马铃薯各项指标的线性模拟
通过胁迫时间与光合特性的线性模拟,用回归直线斜率的显著性差异来反映光合特性在时间梯度上的变化差异,能反映光合指标对水分胁迫的敏感程度。从本研究的线性模拟结果来看,随着胁迫时间的延长,SPAD与光合特性均呈下降趋势,但线性模拟各指标斜率的显著性差异各不相同。T3和T4处理SPAD值、Pn的斜率差异显著,这与Ramírez等[17]的研究结果一致,说明马铃薯遭受轻度水分胁迫(土壤相对含水量为70%)时,SPAD值、Pn对水分较敏感,主要通过调节叶绿素含量,从而影响叶片Pn;T2与T3、T3与T5处理之间的Pn、Gs和Ci斜率差异显著,说明马铃薯遭受中度水分胁迫(土壤相对含水量为55%)或水分过度饱和时,Gs、Ci受影响,从而影响叶片Pn;T1与T2处理之间的Ci斜率差异显著,说明马铃薯遭受重度水分胁迫(土壤相对含水量为40%)时,通过迅速调节Ci来适应水分胁迫。因此,在今后研究中可以将SPAD值、Pn作为首选抗旱指标。
3.4 水分胁迫对马铃薯光合特性基因表达的影响
本研究通过转录组测定和差异基因表达分析认为,马铃薯受到水分胁迫后,KEEG通路中光合作用-天线蛋白通路显著富集,捕光复合体Ⅰ和Ⅱ叶绿素a/b结合蛋白相关基因表达显著上调,说明马铃薯通过加强天线蛋白系统向相连的反应中心有效输送能量,从而响应水分胁迫环境。植物中的环核苷酸门控离子通道(CNGCs)和气孔在抵御非生物胁迫中发挥了重要作用。马铃薯受到水分胁迫后,植物病原体相互作用通路中与CNGCs和CaM、CML相关的基因表达显著下调。马铃薯通过CNGCs调控细胞质内Ca2+含量,使与Ca2+稳态的相关基因表达量显著下调,使得细胞质中Ca2+含量减少,从而关闭气孔来适应水分胁迫环境[18]。本试验在分子水平上揭示马铃薯苗期光合作用相关的基因,这为马铃薯块茎形成期、膨大期、淀粉积累期研究奠定了理论基础,对解释水分胁迫下马铃薯块茎形成期光合特性变化规律提供了分子支撑。综合田间和室内试验可知,水分胁迫对马铃薯的光合特性产生影响,但是在分子水平上,时间梯度的光合作用变化还需要进一步研究。
综上所述,马铃薯产量与土壤相对含水量密切相关,水分胁迫会对马铃薯产量产生影响,其主要原因是通过调整SPAD值、Gs、Ci变化来间接影响叶片Pn和Tr,从而影响了光合作用机制,最终的影响体现在产量上。
猜你喜欢
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
物理之友(2020年12期)2020-07-16 05:39:16
森林工程(2018年4期)2018-08-04 03:23:16
时代农机(2018年11期)2018-03-17 06:05:31
福建中学数学(2016年7期)2016-12-03 07:10:28
陕西画报(2016年1期)2016-12-01 05:35:30
光学精密工程(2016年1期)2016-11-07 09:01:53
电测与仪表(2016年6期)2016-04-11 12:05:54
创新作文(小学版)(2016年31期)2016-03-11 19:08:09
电源技术(2016年9期)2016-02-27 09:05:28
