
2016~2017年四川地区PRRS病毒ORF5基因的遗传变异分析
2018-08-02 11:37:28丁梦蝶朱佳文梁璐琪张毅陈斌邵靓
四川畜牧兽医 2018年7期
丁梦蝶 ,朱佳文 ,梁璐琪 ,张毅 ,陈斌 ,邵靓
(1.四川省动物疫病预防控制中心,四川 成都 610041;2.成都市农林科学院畜牧研究所,四川 温江 611130)
猪繁殖与呼吸综合征(PRRS,俗称猪蓝耳病)于1987年首次在美国发现,我国于1996年首次分离到PRRSV美洲型经典毒株(CH-1a),2006年我国大面积暴发高致病性PRRS,给养猪业造成了巨大的经济损失[1]。2013年以来,我国部分地区又出现了欧洲型PRRSV毒株[2]以及美洲型NADC30类病毒(NADC30-like)[3-4]。PRRSV 是单股正链 RNA 病毒,基因组的变异使得新毒株不断出现。ORF5编码主要囊膜蛋白GP5,该蛋白是病毒与细胞受体结合区,包含病毒中和位点,在结构蛋白中变异率最高,因此ORF5基因是研究PRRSV遗传进化及分子流行病学的重要靶基因。本研究应用RT-PCR法从2016~2017年采集自四川地区的疑似PRRSV感染病料中扩增ORF5基因,并进行序列分析,以了解近两年来PRRSV在四川地区的流行特点,为有效防控该病提供科学依据。
1 材料与方法
1.1 病料 疑似PRRSV感染猪病料来自于2016年和2017年四川地区的流行病学调查采样,保存于-80℃。
1.2 主要试剂 柱式RNA提取试剂盒,购自北京世纪元亨动物防疫技术有限公司;RT-PCR一步法试剂盒,购自宝生物(大连)工程有限公司。
1.3 样品处理 取病死猪的组织样品,用无菌镊子剪碎,加入灭菌PBS(pH 7.2),然后在样品制备系统中破碎,离心后取上清用于RNA提取,提取步骤按照试剂盒说明书进行。
1.4 ORF5基因的扩增和序列测定 使用Zhou L等[5]设计的引物ORF5-F、ORF5-R扩增ORF5基因,预期扩增片段长度为682bp。RT-PCR扩增反应体系:25μL 2×1step buffer、2μL Enzyme mix、上下游引物各 1μL(20μmoL)、4μL RNA、17μL RNase Free H2O,反应总体积为50μL。RT-PCR反应条件:50℃ 30 min,94℃ 2min;94℃ 30s,56℃ 30s,72℃ 50s,共 32 个循环;72℃ 8min。PCR产物经1%琼脂糖凝胶电泳检测,确认有目的条带后送上海生工进行T载体克隆和测序。
1.5 ORF5基因的序列分析 应用DNAStar软件对所测得的ORF5基因序列,以及CH-1a(AY032626)、JXA1 (EF112445)、VR-2332(AY150564)、NADC30(JN654459)、QYYZ(JQ308798)、LV(M96262)等参考毒株的ORF5基因序列进行同源性分析和序列比对。用MEGA6软件Neighbor-Joining法进行遗传变异分析,设置自展值(bootstrap)为1000,构建系统进化树。
2 结果
2.1 ORF5基因的测序结果 经测序后共得到12个ORF5基因序列。对测序结果进行剪切后,长度均为603bp,编码200个氨基酸。
2.2 序列同源性分析 12个ORF5基因序列之间的核苷酸序列同源性为82.3%~99.8%,推导的氨基酸序列同源性为81.1%~99.5%。与美洲型毒株CH-1a、JXA1、VR-2332、NADC30、QYYZ 的 ORF5 基因的核苷酸序列同源性分别为85.1%~95.4%、83.6%~99.5%、85.1%~89.4%、83.9%~94.2%、82.8%~90.9%,与欧洲型代表株LV的核苷酸序列同源性仅为61.4%~64.6%;推导的氨基酸序列同源性分别为85.6%~93.5%、83.6%~98.5%、83.1%~88.6%、84.1%~93.5%、80.6%~93.0%,与LV氨基酸序列的同源性仅为53.3%~58.9%。
2.3 遗传变异分析 基于ORF5基因序列构建的遗传进化树如图1所示。
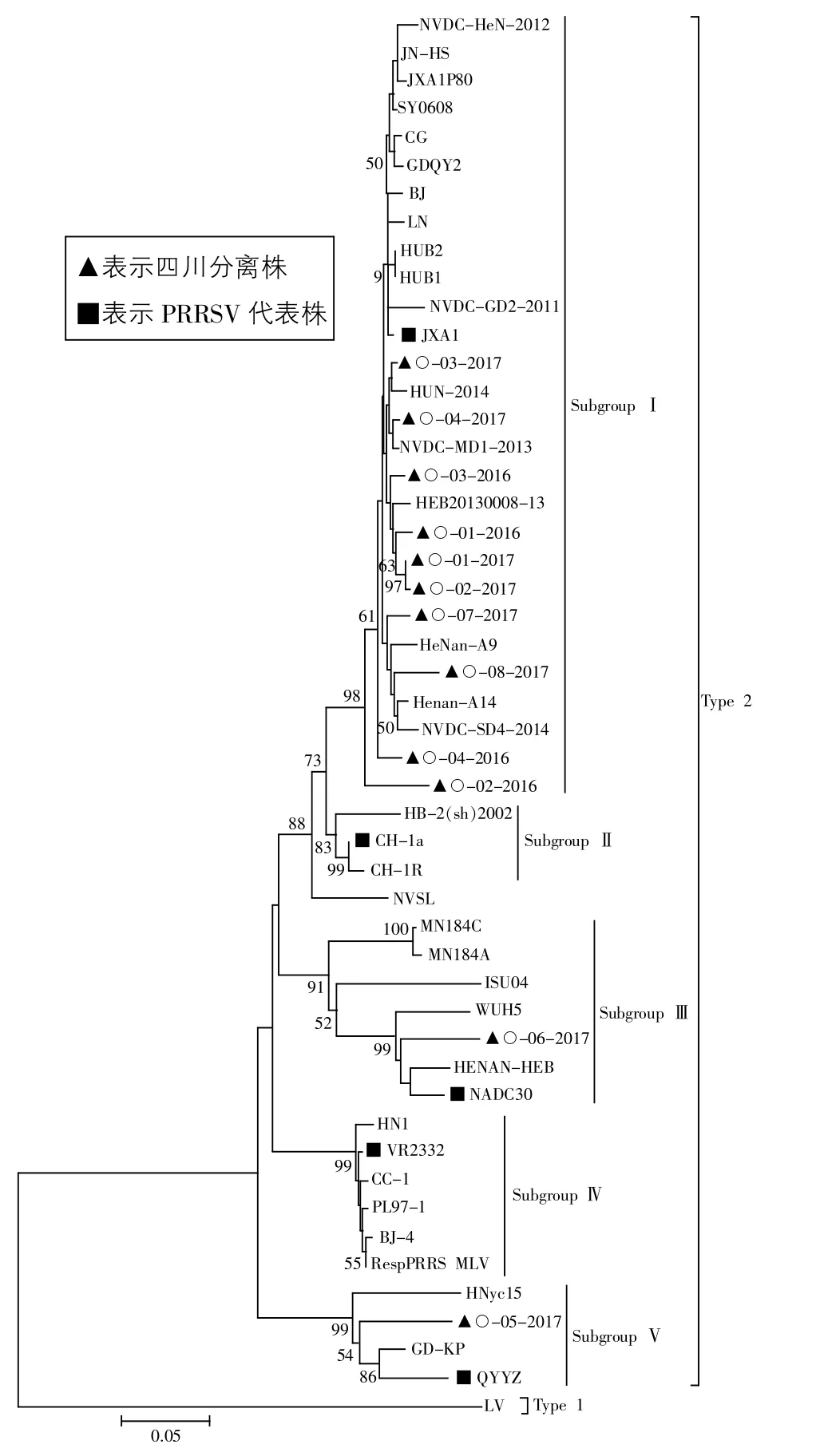
图1 基于ORF5基因序列的遗传进化树
PRRSV毒株分为美洲型(Type 2)和欧洲型(Type 1)两个基因型。美洲型毒株可以分为5个亚群,分别以JXA1、CH-1a、NADC30、VR-2332 和 QYYZ 毒株为代表。本研究中的12个序列均属于美洲型,其中有10个 (O-01-2016、O-02-2016、O-03-2016、O-04-2016、O-01-2017、O-02-2017、O-03-2017、O-04-2017、O-07-2017、O-08-2017)属于以 JXA1 毒株为代表的 I亚群,另 2个(O-06-2017、O-05-2017)分别属于以NADC30株为代表的Ⅲ亚群和以QYYZ毒株为代表的V亚群。
2.4 ORF5基因推导的氨基酸比对分析 抗原表位区域和毒力位点均存在氨基酸变异。与所在亚群的代表株相比,在 B 细胞表位 1(27~30aa)中,O-08-2017株发生 V29→A29突变,O-06-2017株发生A27→V27突变;在 B 细胞表位 2(37~45aa)中,O-02-2016株发生I39→F39突变;在B细胞表位3(180~197aa)中,O-02-2016株发生 L196→Q196突变,O-04-2016株发生 R191→K191突变,O-07-2017、O-08-2017株发生L196→R196突变。在T细胞表位区域,与所在亚群的代表株相比,I亚群中O-02-2016株发生 I121→V121、V124→I124突变,O-07-2017株发生R151→K151突变,O-08-2017株发生I121→L121突变;Ⅲ亚群中O-06-2017株发生120LI121→120FF121、K129→R129、K163→R163 突变;V亚群中O-05-2017株发生F117→L117、I121→T121、I124→V124、Y153→H153 突变。在毒力位点的第 13 位 aa,O-05-2017、O-06-2017 株为 Q13,其余为 R13;第 151 位 aa,O-05-2017、O-06-2017、O-07-2017株为K151,其余为R151。
3 讨论
在PRRSV的结构蛋白中,普遍认为编码GP5蛋白的ORF5基因最容易发生变异,因此可通过分析ORF5基因的变异情况来研究PRRSV的分子流行病学[6-7]。PRRSV主要分为以LV株为代表的欧洲型毒株和以VR-2332株为代表的美洲型毒株,两者之间的核苷酸序列同源性约60%。本研究中12个美洲型ORF5序列与LV株ORF5基因的核苷酸序列同源性仅为61.4%~64.6%,符合这一现象。此外,虽然都属于美洲型,但12个ORF5序列之间的核苷酸序列和推导的氨基酸序列的同源性相互之间有较大差异,也符合ORF5基因变异大的特点。
PRRSV流行毒株的ORF5基因在抗原表位区域和毒力位点均有发生氨基酸突变的报道[8]。在抗原表位区域有6个ORF5序列(O-02-2016、O-04-2016、O-05-2017、O-06-2017、O-07-2017、O-08-2017)发生了以下突变:A27→V27、V29→A29、I39→F39、I121→V121、V124→I124、K163→R163、R191→K19 1、L196→R196、L196→Q196 等。这些突变可能会引起抗原性改变,进而影响疫苗免疫效果。在毒力相关位点,强毒株为R13和R151,弱毒株为Q13和G151。在第 13 位 aa,2 株(O-05-2017、O-06-2017)出现毒力致弱(R13→Q13);在第 151位 aa,3株(O-05-2017、O-06-2017、O-07-2017)可能出现毒力改变(R151→K151)。此外,已知ORF5序列的前31位氨基酸为信号肽序列,是GP5蛋白的高变区,本研究中除 O-02-2016、O-03-2016、O-04-2016 外的其他序列在第10~30aa均有氨基酸突变。
4 结论
本研究12个ORF5序列中,有10株与高致病性毒株JXA1的亲缘关系较近,有1株与NADC30毒株的亲缘关系较近,另1株与广东分离株QYYZ的亲缘关系较近,可见四川地区近两年流行的PRRSV以高致病性的JXA1亚型为主,也有少量NADC30亚型和QYYZ亚型毒株出现,应密切关注。
猜你喜欢
遵义医科大学学报(2023年4期)2023-05-05 05:05:16
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
小哥白尼(野生动物)(2021年12期)2021-03-29 01:15:40
娃娃乐园·综合智能(2019年10期)2019-12-07 05:27:34
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
中成药(2018年2期)2018-05-09 07:19:51
中国免疫学杂志(2017年1期)2017-01-17 04:53:25
畜牧兽医学报(2015年3期)2015-07-05 08:22:53
医学研究杂志(2015年6期)2015-07-01 17:41:11
