
盐胁迫条件下杨树bZIP转录因子全基因组分析
2018-08-02 12:22王升级孙赫党慧
山西农业大学学报(自然科学版) 2018年8期
王升级,孙赫,党慧
(1.山西农业大学 林学院,山西 太谷 030801;2.国家林业和草原局,北京 100714)
植物在生长发育过程中常受到干旱、低温、盐碱、虫害等不同生物及非生物因子的影响。为适应多变的环境,植物体内大量胁迫相关的基因被诱导表达,同时抗逆相关蛋白得到积累[1]。转录因子(transcription factor)又称为反式作用因子,在植物应答逆境胁迫生物学过程中起到开关的作用,能够通过调控目的功能基因的表达提高植物的抗逆性[2]。bZIP转录因子家族是植物中最大的转录因子家族之一。bZIP转录因子一般包含一个高度保守的bZIP结构域。在bZIP保守结构域外往往还含有起转录激活作用的脯氨酸富集区和谷氨酸富集区[3]。该转录因子家族成员在植物种子成熟萌发、花器官发育、激素和光信号转导、ABA胁迫诱导等生物及非生物胁迫应答过程中发挥着重要的作用。尤其在植物的应激反应和激素信号转导途径中,bZIP转录因子通过与ABRE作用元件(ABA responsive element)结合调控目的靶基因的表达[4]。根据其结构和功能的特点,bZIP转录因子家族常被分为A、B、C、D、E、F、G、H、I、S等不同的亚族[5]。拟南芥AtbZIP39转录因子基因参与ABA胁迫信号转导途径,并且能够通过调控LEA基因的表达参与种子萌发过程[6,7];玉米bZIP转录因子基因Opaque2与PBF蛋白互作调控种子贮藏蛋白的合成[8]。转录因子基因AtbZIP57/OBF4/TGA4通过参与水杨酸或者乙烯合成途径应答植物病原侵染反应。在应对病原体攻击反应中,植物体内的水杨酸诱导抗病相关基因的表达,TGA元件能够与抗病相关基因启动子中的as-1顺式作用元件结合进而提高植物的抗病能力[9,10]。近年来,关于草本植物及农作物中bZIP转录因子的研究报道较多,而涉及木本植物杨树的研究较为少见。自2004年杨树基因组测序完成以后,伴随分子生物学及生物信息学的发展,杨树已经被广泛用作研究林木分子育种及逆境抗性育种的模式植物[11]。
本研究为明确杨树bZIP转录因子家族的结构特征和在应答盐胁迫过程中的作用机制,通过生物信息学分析获得杨树bZIP转录因子214个,分析其序列及蛋白质二级结构特征;盐胁迫条件下,利用转录组测序分析其盐胁迫条件下表达模式,鉴定出应答盐胁迫bZIP转录因子基因19个,为杨树耐盐关键基因筛选及bZIP转录因子功能研究提供参考依据。
1 材料和方法
1.1 试验材料
将4周大小的无菌组织培养84 K杨树(Populusalba×P.glandulosa)幼苗置于温室内水培,温室温度控制在24 ℃左右,光照时间为14 h·d-1。约4周后,选取长势相似且健壮的植株分为对照组(W1,W2,W3,W4)与试验组(S1,S2,S3,S4)二组,每组设置生物学重复4次。对照组与实验组分别用水和150 mmol·L-1NaCl 溶液处理24 h。处理后分别采集对照与试验组的杨树幼苗叶片组织,液氮中迅速冷冻后于-80 ℃超低温冰箱中保存以备总RNA提取及转录组测序使用。
1.2 试验方法
利用生物信息学分析方法从PlantTFDB(http://planttfdb.cbi.pku.edu.cn/index.php)数据库检索到杨树bZIP转录因子214条,命名为PtbZIP1- PtbZIP214。分别利用软件ClustalX 1.83和MEGA 7.0 进行保守域氨基酸序列比对分析及进化树的构建(Bootstrap:1000)。利用软件DNAMAN 6.0预测杨树bZIP转录因子家族二级结构。
RNA-seq由金唯智(GENEWIZ)测序服务公司完成。其测序文库构建过程包括:RNA质量检测;文库构建;文库纯化;文库检测;文库定量分析;cBOT 自动成簇。检测合格后,按照有效浓度及测序需求混样后进行Illumina Hiseq2500测序。
RNA-seq 数据分析过程包括:利用软件Fastqc检测测序数据质量;移除多余接头以获得高纯度质量序列片段;利用软件Tophat将序列片段与杨树参照基因组进行比对;利用软讲Cufflinks将比对成功的序列片段进行拼接并量化基因表达量。基因表达量以每百万条成功比对的序列中每1 000 bp转录组含有的序列片段数量(Fragments per kilo-base Transcript per million mapped reads, FPKM)表示。
1.3 统计分析
本文利用单因素变量分析方法t检测比较试验处理前后各基因表达水平的差异,其中P≤0.05的定义为差异表达基因;利用无监督聚类方法(unsupervised clustering analysis)对表达模式相似的差异表达基因进行聚类分析。数据统计分析处理均由R软件(http://cran.r-project.org/)完成。
2 结果与分析
2.1 杨树bZIP转录因子家族比对分析
利用生物信息学分析方法获得214个杨树bZIP转录因子(表1),参照拟南芥bZIP转录因子分类方法,根据杨树bZIP转录因子保守结构域氨基酸序列比对结果,将其分为11个亚族:A(PtbZIP122、PtbZIP163、PtbZIP164)、B(PtbZIP115、PtbZIP201、PtbZIP202)、C(PtbZIP28、PtbZIP29、PtbZIP30、PtbZIP92、PtbZIP109、PtbZIP127、PtbZIP128、PtbZIP129、PtbZIP130、PtbZIP131、PtbZIP132、PtbZIP133、PtbZIP134、PtbZIP176、PtbZIP177、PtbZIP178、PtbZIP179、)、D(PtbZIP4、PtbZIP5、PtbZIP6、PtbZIP7、PtbZIP8、PtbZIP9、PtbZIP10、PtbZIP11、PtbZIP12、PtbZIP13、PtbZIP34、PtbZIP35、PtbZIP36、PtbZIP37、PtbZIP53、PtbZIP54、PtbZIP55、PtbZIP56、PtbZIP57、PtbZIP58、PtbZIP59、PtbZIP60、PtbZIP61、PtbZIP86、PtbZIP87、PtbZIP88、PtbZIP89、PtbZIP93、PtbZIP94、PtbZIP95、PtbZIP96、PtbZIP97、PtbZIP102、PtbZIP103、PtbZIP104、PtbZIP116、PtbZIP117、PtbZIP118、PtbZIP119、PtbZIP120、PtbZIP121、PtbZIP140、PtbZIP141、PtbZIP142、PtbZIP150、PtbZIP151、PtbZIP161、PtbZIP162、PtbZIP170、PtbZIP203、PtbZIP204)、E(PtbZIP22、PtbZIP23、PtbZIP24、PtbZIP25、PtbZIP181、PtbZIP211、PtbZIP212)、F(PtbZIP2、PtbZIP3、PtbZIP62、PtbZIP63、PtbZIP78、PtbZIP79、PtbZIP80、PtbZIP81、PtbZIP82、PtbZIP83、PtbZIP84、PtbZIP85、PtbZIP157、PtbZIP158、PtbZIP159、PtbZIP160)、G(PtbZIP16、PtbZIP17、PtbZIP44、PtbZIP50、PtbZIP51、PtbZIP52、PtbZIP99、PtbZIP100、PtbZIP101、PtbZIP137、PtbZIP138、PtbZIP139、PtbZIP192、PtbZIP193、PtbZIP194、PtbZIP195、PtbZIP196)、H(PtbZIP165、PtbZIP166、PtbZIP167、PtbZIP168、PtbZIP169、 PtbZIP207、PtbZIP208、PtbZIP209)、I(PtbZIP32、PtbZIP33、PtbZIP76、PtbZIP77、PtbZIP105、PtbZIP106、PtbZIP107、PtbZIP143、PtbZIP144、PtbZIP156、PtbZIP180、PtbZIP182、PtbZIP183、PtbZIP184、PtbZIP206、PtbZIP213、PtbZIP214)、S(PtbZIP27、PtbZIP75、PtbZIP98、PtbZIP110、PtbZIP136、PtbZIP155)及其它(图1)。

表1 杨树bZIP转录因子Table 1 Populus bZIP transcription factors
续表

转录因子号TF_id编号PtbZIPno.转录因子号TF_id编号PtbZIPno.转录因子号TF_id编号PtbZIPno.转录因子号TF_id编号PtbZIPno.Potri.001G374200.2PtbZIP23Potri.004G163800.2PtbZIP77Potri.006G277800.5PtbZIP131Potri.014G007100.1PtbZIP185Potri.001G374200.3PtbZIP24Potri.004G175200.1PtbZIP78Potri.006G277800.6PtbZIP132Potri.014G013400.1PtbZIP186Potri.001G374200.4PtbZIP25Potri.004G175200.2PtbZIP79Potri.006G277800.7PtbZIP133Potri.014G028200.1PtbZIP187Potri.002G018400.1PtbZIP26Potri.004G175200.3PtbZIP80Potri.006G277800.8PtbZIP134Potri.014G028200.2PtbZIP188Potri.002G031900.1PtbZIP27Potri.004G175200.4PtbZIP81Potri.007G006900.1PtbZIP135Potri.014G028200.3PtbZIP189Potri.002G045800.1PtbZIP28Potri.004G175200.5PtbZIP82Potri.007G019900.1PtbZIP136Potri.014G028200.4PtbZIP190Potri.002G045800.3PtbZIP29Potri.004G175200.6PtbZIP83Potri.007G029400.1PtbZIP137Potri.014G028200.5PtbZIP191Potri.002G045800.4PtbZIP30Potri.004G175200.7PtbZIP84Potri.007G029400.2PtbZIP138Potri.014G062400.1PtbZIP192Potri.002G067400.1PtbZIP31Potri.004G175200.8PtbZIP85Potri.007G029400.3PtbZIP139Potri.014G062400.2PtbZIP193Potri.002G069500.1PtbZIP32Potri.004G203400.1PtbZIP86Potri.007G085700.1PtbZIP140Potri.014G094200.1PtbZIP194Potri.002G069500.2PtbZIP33Potri.004G203400.2PtbZIP87Potri.007G085700.2PtbZIP141Potri.014G094200.2PtbZIP195Potri.002G090700.1PtbZIP34Potri.004G203400.3PtbZIP88Potri.007G085700.3PtbZIP142Potri.014G094200.3PtbZIP196Potri.002G090700.2PtbZIP35Potri.004G203400.4PtbZIP89Potri.007G130800.1PtbZIP143Potri.014G120800.1PtbZIP197Potri.002G090700.4PtbZIP36Potri.004G219100.1PtbZIP90Potri.007G130800.2PtbZIP144Potri.015G047700.1PtbZIP198Potri.002G090700.5PtbZIP37Potri.004G219100.2PtbZIP91Potri.008G010800.1PtbZIP145Potri.016G024000.1PtbZIP199Potri.002G090800.1PtbZIP38Potri.005G053200.1PtbZIP92Potri.008G018400.1PtbZIP146Potri.016G024000.2PtbZIP200Potri.002G115900.1PtbZIP39Potri.005G082000.1PtbZIP93Potri.008G018400.2PtbZIP147Potri.016G032400.1PtbZIP201Potri.002G125400.1PtbZIP40Potri.005G082000.2PtbZIP94Potri.008G106700.1PtbZIP148Potri.016G032400.2PtbZIP202Potri.002G125400.2PtbZIP41Potri.005G082000.3PtbZIP95Potri.008G113400.1PtbZIP149Potri.016G036500.1PtbZIP203Potri.002G125400.3PtbZIP42Potri.005G082000.4PtbZIP96Potri.008G118300.1PtbZIP150Potri.016G049200.1PtbZIP204Potri.002G125400.4PtbZIP43Potri.005G082000.5PtbZIP97Potri.008G118300.2PtbZIP151Potri.016G121700.1PtbZIP205Potri.002G167100.1PtbZIP44Potri.005G119300.1PtbZIP98Potri.009G018500.1PtbZIP152Potri.017G027400.1PtbZIP206Potri.002G196200.1PtbZIP45Potri.005G126000.1PtbZIP99Potri.009G075000.1PtbZIP153Potri.017G106700.1PtbZIP207Potri.002G196200.2PtbZIP46Potri.005G126000.2PtbZIP100Potri.009G101200.1PtbZIP154Potri.018G029500.1PtbZIP208Potri.003G014500.1PtbZIP47Potri.005G126000.3PtbZIP101Potri.009G119700.1PtbZIP155Potri.018G029500.2PtbZIP209Potri.003G014800.2PtbZIP48Potri.005G170500.1PtbZIP102Potri.009G125400.1PtbZIP156Potri.018G029500.3PtbZIP210Potri.003G014800.3PtbZIP49Potri.005G170500.2PtbZIP103Potri.009G134900.1PtbZIP157Potri.019G091900.1PtbZIP211Potri.003G097600.1PtbZIP50Potri.005G170500.4PtbZIP104Potri.009G134900.2PtbZIP158Potri.019G091900.2PtbZIP212Potri.003G097600.2PtbZIP51Potri.005G190700.1PtbZIP105Potri.009G134900.3PtbZIP159Potri.019G130000.1PtbZIP213Potri.003G097600.3PtbZIP52Potri.005G190700.2PtbZIP106Potri.009G134900.4PtbZIP160Potri.019G130000.2PtbZIP214Potri.003G194600.1PtbZIP53Potri.005G190700.3PtbZIP107Potri.009G164300.1PtbZIP161Potri.003G194600.2PtbZIP54Potri.005G192900.1PtbZIP108Potri.009G164300.2PtbZIP162
2.2 杨树bZIP转录因子二级结构预测
蛋白质二级结构是指其多肽链有规则的重复构象,主要包括α-螺旋、β-折叠、β-转角及无规则卷曲4种构象。利用软件DNAMAN6.0预测杨树bZIP家族转录因子二级结构显示,bZIP转录因子序列两端主要以卷曲为主,而序列中间部分以α-螺旋构象为主(图2)。
2.3 杨树应答盐胁迫基因鉴定
利用软件Bioconductor的DESeq(V1.12.1)包分析RNA-seq测序数据,获得65 536条基因在不同样品中(W1 vs S1, W2 vs S2, W3 vs S3, W4 vs S4)差异表达结果,对其按照显著性标准(基因表达差异2倍以上且P≤0.05)进行筛选,统计显著上调或下调差异表达基因(图3)。结果表明,W1 vs S1与W4 vs S4两组下调表达基因较多;W2 vs S2与W3 vs S3两组上调表达基因较多。但每组差异表达基因均占基因总数的0.9%左右,不同组间差异可能是由试验材料差异造成的生物学重复误差。
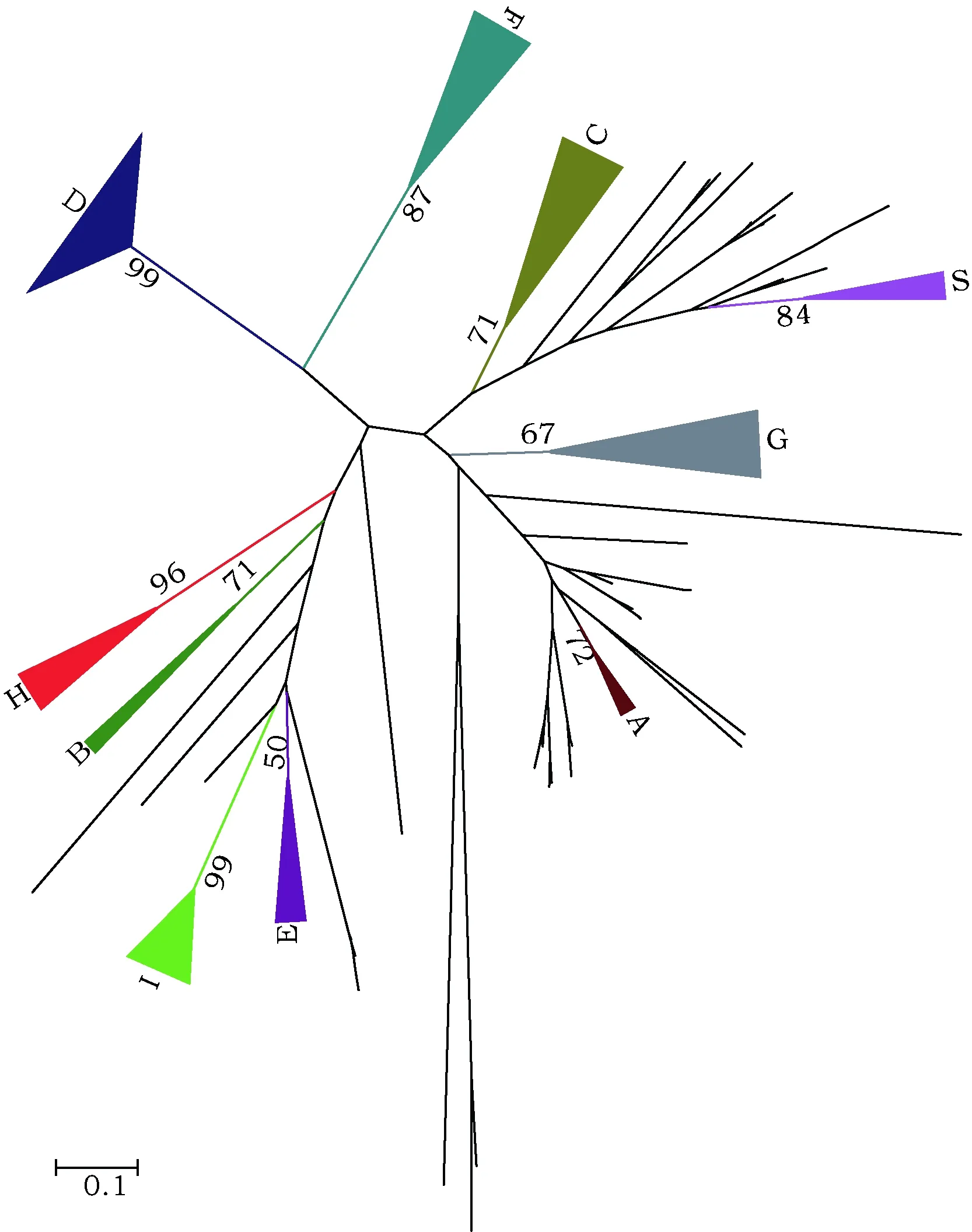
图1 杨树bZIP转录因子家族保守域序列比对及无根系统发生树构建Fig.1 Multiple alignment and unrooted phylogenetic tree construction of poplar bZIP transcription factors
2.4 杨树应答盐胁迫bZIP转录因子基因表达模式分析
利用生物信息学分析方法结合转录组测序数据,从214条杨树bZIP转录因子基因中筛选出应答盐胁迫基因19条(图4)。其中盐胁迫处理后,上调表达基因14条,下调表达基因5条。大部分差异表达基因位于1号及2号染色体上。PtbZIP36和PtbZIP195盐胁迫处理后基因表达量分别达到了200和100左右,其他基因表达水平相对较低。
2.5 杨树应答盐胁迫bZIP转录因子基因聚类分析
根据基因表达量(FPKM),对杨树应答盐胁迫bZIP转录因子基因进行聚类分析,结果表明,19个应答盐胁迫差异表达的基因大致可为分为表达量高、中、低3类(图5)。其中高表达量的基因只有PtbZIP36一个;中表达量的基因有PtbZIP40及PtbZIP195两个;其余16个基因在盐胁迫处理前后的表达量均相对较低。

图2 杨树bZIP转录因子二级结构预测Fig.2 Secondary structure prediction of Populus bZIP transcription factors
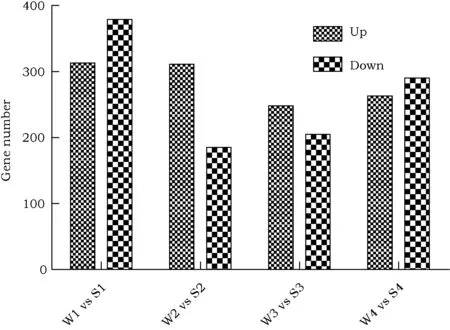
图3 盐胁迫条件下差异表达基因统计Fig.3 Differential expression of genes in response to salt stress
3 讨论与结论
植物体内bZIP转录因子一般由60~80氨基酸残基组成,包含2个位于连续α-螺旋序列中的高度保守的结构域:一个是含有约16个氨基酸的碱性结构域,该结构域能够通过N-x7-R/K元件与DNA结合,起到核定位信号的作用;另一个是由位于氨基酸序列C端第9位的亮氨酸或者其它疏水性氨基酸残基形成的亮氨酸拉链区(亮氨酸每隔7个氨基酸残基重复出现一次),该区域组成的水脂双亲性螺旋结构通过参与蛋白质二聚体化进而参与bZIP转录因子与DNA的互作[5]。
本研究参考拟南芥bZIP转录因子分类方法,将杨树bZIP转录因子分为11个亚族(图1)。研究表明,除I亚族外,A、B、C、D、E、F、G、H及S亚族均含有相对保守一致的bZIP结构域[5]。A 亚族bZIP转录因子大多与植物体内ABA信号及胁迫信号转导相关,如拟南芥bZIP转录因子基因AtbZIP39/ABI5,bZIP36/ABF2/AREB1,AtbZIP38/ABF4/AREB2,AtbZIP66/AREB3,AtbZIP40/GBF4,AtbZIP35/ABF1 及AtbZIP37/ABF3等[12, 13]。C亚族转录因子具有显著的结构特点,在几乎所有植物内均含有一个延伸至9次重复的亮氨酸拉链结构,其潜在的调控核转运与DAN结合特性的磷酸化修饰位点序列亦高度保守。C亚族转录因子主要参与种子发育及生物胁迫与非生物胁迫应答过程[8, 14]。D亚族转录因子通过与TGA因子互作在植物抵御逆境胁迫过程中发挥重要作用,还有少数D亚族转录因子能够参与调控植物的发育过程[10 ]。关于E亚族转录因子功能的研究报道较少,其与I亚族在亮氨酸拉链区具有高度的相似性,但是由于第10位赖氨酸的缺失,将其定义为不同的亚族[5]。G亚族转录因子主要参与紫外线及蓝光信号转导等光信号应答过程,在种子成熟过程中也起到调控作用[15]。H亚族转录因子主要参与植物的光合作用过程。I亚族转录因子主要与导管发育相关,调控赤霉素生物合成和韧皮部形成过程。S亚族转录因子不仅参与植物糖类代谢过程,在逆境胁迫应答及花器官发育过程中也发挥重要作用[5]。

图4 杨树bZIP转录因子基因应答盐胁迫表达模式分析Fig.4 Expression pattern analysis of Poplulus bZIP transcription factor genes in response to salt stress
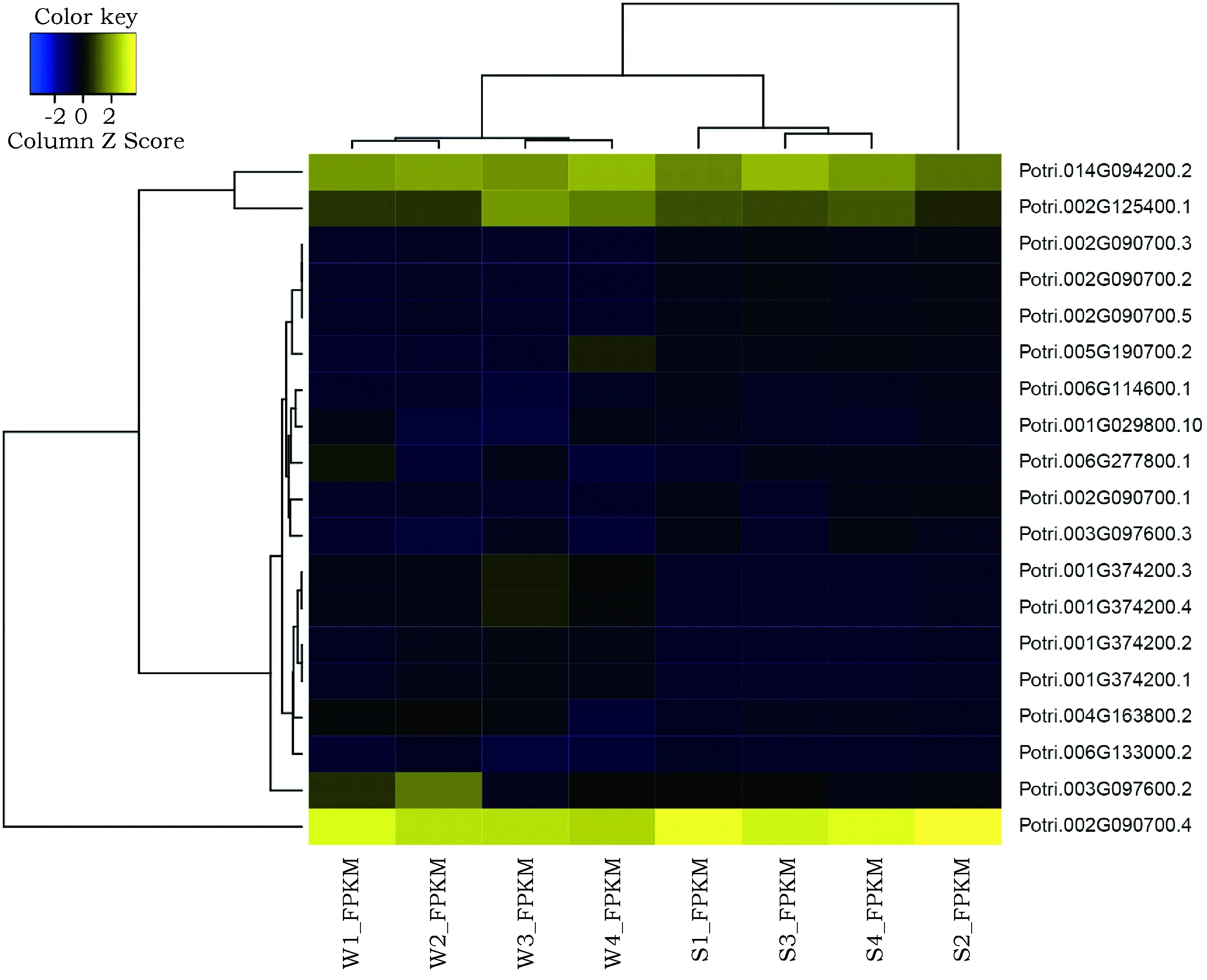
图5 杨树应答盐胁迫bZIP转录因子基因聚类分析Fig.5 Gene clustering analysis of Poplulus bZIP transcription factor genes in response to salt stress
通过RNA-seq数据分析筛选出杨树应答盐胁迫bZIP转录因子19个,约占bZIP家族成员的10%。进一步分析表明,19个bZIP差异表达基因中有6(约1/3)个属于D亚族,其他bZIP差异表达基因主要属于C亚族与G亚族,结果与植物C、D、G亚族bZIP转录因子在植物生长发育及抵御逆境胁迫中发挥重要作用的结论基本一致[5],说明bZIP转录因子家族在杨树盐胁迫逆境应答过程中发挥着重要的作用,为杨树bZIP转录因子功能的进一步研究奠定理论基础。
猜你喜欢
江苏农业科学(2022年17期)2022-09-24
江苏农业科学(2022年16期)2022-08-26
湖北农业科学(2022年11期)2022-07-18
四川大学学报(自然科学版)(2021年4期)2021-07-15
实用肿瘤学杂志(2020年4期)2020-12-08
中国农业科学(2020年5期)2020-03-30
时代英语·高一(2019年1期)2019-03-13
喜剧世界(2017年10期)2017-06-01
中国老年学杂志(2015年9期)2015-01-31
医学综述(2011年12期)2011-12-09
