
裸果木种群遗传多样性特点及与地理气候因子关联研究
2018-07-30 03:28:56徐振朋张晓明伊卫东宛诣超
草地学报 2018年1期
徐振朋, 宛 涛*, 蔡 萍, 张晓明, 伊卫东, 宛诣超
(1. 内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019; 2. 锡林郭勒职业学院,内蒙古 锡林浩特 026000;3. 内蒙古农业大学农学院,内蒙古 呼和浩特 010019)
裸果木为石竹科(Caryophyllaceae)裸果木属(Gymnocarpos)多年生小半灌木,国家一级保护植物,起源于第三纪,古地中海残遗成分[1-2]。分布于甘肃河西走廊、内蒙古西部、宁夏中西部和新疆东部[3]。生境较为单一,主要生长在海拔为800~2 500 m荒漠区的干河床、干河道、山前洪积扇及砾石戈壁滩等地。在防风固沙、水土保持方面发挥十分重要的作用,对维护当地生态系统稳定具有着不可替代的作用[1]。分布区年均温为4.1~9.3℃,≥10℃年积温在3 000~4 000℃之间,极端最高温达40.0℃,极端最低温达到-31.6℃;年均日照时数为2 900~3 400 h左右;年均降水量50~170 mm之间,集中在7~9月;年蒸发量2 100~3 300 mm。从植物区系组成及水热特征来看,为典型的戈壁荒漠和荒漠化草原。目前对于裸果木主要集中于种群生态学[4-5]、分类学[6-7]、繁殖生物学[8-10]等方面研究。因群落结构简单、生态系统脆弱、自然地理环境变迁及人类活动干扰等因素,使裸果木种群斑块状分布格局更趋于明显,分布区面积缩小,种群数量锐减。
遗传多样性是生物体携带遗传信息的总和,是生物多样性的重要组成部分,维持生物繁殖活力及与环境变化相互适应的基础。物种在受环境压力的影响,除逃避和表型可塑性外,主要是通过改变遗传基础来适应环境[11]。因此,种群或物种遗传多样性越高,对环境变化的适应能力越强,越容易扩展其分布范围[12]。简单序列重复区间扩增多态性(inter simple sequence repeat,ISSR)分子标记技术以简便快捷、多态性高和重复性好等优点在遗传多样性评价方面得到广泛应用[13-15]。为深入了解裸果木遗传基础、斑块状分布的种群遗传结构、地理气候因素对种群遗传多样性水平的影响,以分布我国甘肃河西走廊及内蒙古西部地区裸果木种群为研究对象,利用ISSR遗传标记技术对不同地理种群进行遗传评价。从分子水平上探讨裸果木种群间的亲缘关系及遗传多样性水平,进而为其种质资源的科学化管理及保育提供依据。
1 材料与方法
1.1 试验材料
2015年5月对分布于内蒙古、宁夏、新疆及甘肃河西走廊区域12个种群进行了考察,每个种群采集20个植株健康叶片,每株间隔距离10 m以上共计采集240株。将采集的新鲜叶片立即放入加变色硅胶自封袋中干燥,带回实验室保存于-80℃冰箱中备用。记录样地经纬度、海拔等地理信息及收集当地气象部门提供近10年离样地最近的气象数据。此外,由于裸果木多生长在偏僻无人的地区,部分气象站点距离样地较远。因此,通过DIVA软件进行当前气候数据提取(精确度为2.5 min),对样地气候数据进一步校准及补充,样地地理及气候信息如表1所示。
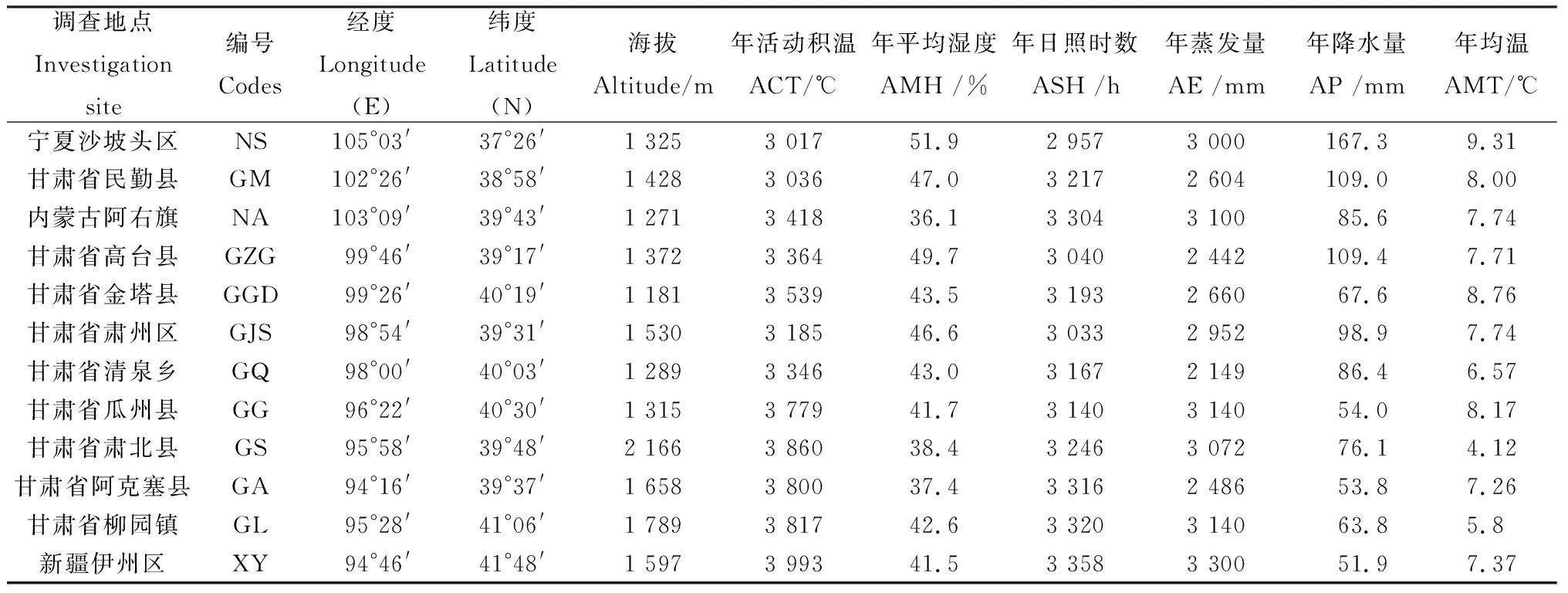
表1 裸果木种群试验区及样地地理及气候信息Table 1 The test site and sample plot’s information of the population of Gymnocarpos przewalskii
1.2 研究方法
1.2.1DNA提取 采用改进CTAB法进行DNA提取[16];利用紫外分光光度法检测DNA浓度,并用1.0%的琼脂糖凝胶电泳检测其完整性,-20℃环境保存备用。
1.2.2引物筛选与PCR扩增 采用UBC(GA)8C引物进行裸果木PCR扩增条件筛选,初步确定ISSR-PCR的最佳扩增程序为:94℃预变性3 min;94℃变性30 s,54℃复性45 s,72℃延伸1.5 min,35个循环;72℃延伸10 min,4℃保存。正交试验优化ISSR-PCR反应体系:在25 μL反应体系中dNTP 浓度为0.30 mmol·L-1,引物0.50 μmol·L-1,Mg2+为2.50 mmol·L-1,Taq DNA酶为1.00 U·μL-1,DNA 浓度为40.00 ng·μL-1效果最佳。依据反应体系对100 个 ISSR引物进行筛选,从中选取扩增条带清晰,重复性较好的引物。
1.3 数据分析
用POPGENG 32软件对种群间和种群内遗传参数分析,利用NTSYS-pc 2.1软件采用UPGMA法构建种群亲缘关系树状聚类图;建立不同种群裸果木的地理(经纬度)的距离矩阵,利用SAS 9.0软件对遗传距离与地理距离进行相关性检测。同时,将遗传参数与气候因子进行相关性分析。
2 结果分析
2.1 裸果木ISSR标记引物筛选
对100条ISSR引物进行筛选,最终筛选出15条扩增清晰,重复性好的引物,用于不同种群裸果木的遗传多样性分析,引物扩增条带及退火温度如表2所示。对12个种群的240个裸果木植株进行扩增,总计扩增出条带数为143,具有多态为条带数127,引物多态位平均比例为88.81%。
2.2 裸果木种群遗传多样性分析
不同种群裸果木遗传多态参数如表3所示,其中多态位点百分率(PPL)变化范围为61.54%~81.12%,平均多态位点百分率(PPL)为69.93%,较高的有甘肃省阿克塞县(GA)种群(81.12%)及甘肃省肃北县(GS)种群(72.73%)等,其次分别为宁夏沙坡头区(NS)、甘肃省柳园镇(GL)、甘肃省清泉乡(GQ)及甘肃省金塔县(GGD)等种群,最低为内蒙古阿右旗(NA)种群(61.54%)。种群中有效等位基因数(Ne)变化范围为1.4035~1.4624,较高的为甘肃省柳园镇(GL)种群,最低为甘肃省金塔县(GGD)种群。种群间He变化范围为0.2356~0.2697,平均值为0.2418;种群间I变化为0.3509~0.4057,平均值为0.3629。甘肃省阿克塞(GA)与甘肃省肃北县(GS)种群的He及I值均高于其他种群,具有丰富的遗传多样性,而内蒙古阿拉善右旗(NA)及甘肃金塔县(GGD)2个种群He及I较低。
注:Na:观测等位基因数;Ne:有效等位基因数;He:种群内遗传多样性;I:Shannon 信息指数;PPL:多态位点;下同
Note:Na: Observed number of alleles;Ne:Effective number of alleles;He: Gene Diversity within Populations;I:Shannon information index;PPL: Percentage of polymorphic loci;the same as below
裸果木种群基因多样性分析如表4所示,其中总的遗传多样性(Ht)为0.3452,而种群内的遗传多样性(Hs)为0.2489,群体间遗传分化系数Gst为0.2790,即种群间的遗传变异占总变异的27.90%,表明裸果木遗传多样性主要存在于种群内,而种群间也存在一定的分化。种群间基因流(Nm)为1.2920,表明种群之间存在着遗传交换。
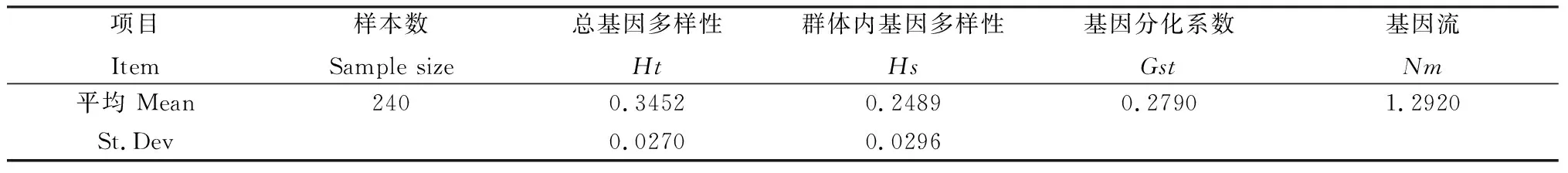
表4 ISSR标记对裸果木种群基因多样性Nei’s分析Table 4 Nei’s analysis of genetic diversity of Gymnocarpos przewalskii population by ISSR
2.3 遗传多样性参数与地理气候因子相关性分析
裸果木地理距离与遗传距离之间的相关性分析如表5所示,亲缘关系最近的甘肃省高台县(GZG)和甘肃省肃州区(GJS)的遗传一致度为0.9929,其地理距离约为79.19 km,而地理距离约为1004.78。宁夏沙坡头区域(NS)和新疆伊州区(XY)种群遗传一致度为0.7622,裸果木种群的遗传距离与地理距离存在一定的相关性(R2=0.2144)。

表5 裸果木种群遗传距离和地理距离Table 5 Nei’s genetic distance and geographic distance of Gymnocarpos przewalskii
注:遗传一致度(对角线下方)和地理距离(对角线上方)
Note: Genetic identity (below diagonal)and geographic distance (above diagonal) /km
依据Nei’s遗传一致度对12个种群裸果木进行UPGMA聚类分析,结果如图1所示,以河西走廊中西部为中心划分为4类群:甘肃省高台县(GZG)、甘肃省清泉乡(GQ)、甘肃省肃州区(GJS)、甘肃省金塔县(GGD)为中心区的东部为一类,与向东和向东南延伸的内蒙古阿右旗(NA)、宁夏沙坡头区域(NS)和甘肃省民勤县(GM)亲缘关系较近;甘肃省瓜州(GG)、甘肃省肃北县(GS)及甘肃省柳园镇(GL)为中心区的西部区种群,与向西南和向西延伸的甘肃省阿克塞县(GA)和新疆伊州区(XY)种群亲缘关系较近。
对裸果木种群ISSR标记的各遗传多样性参数与地理气候因子进行相关分析,结果如表6所示,ISSR标记的裸果木遗传多样性与地理、气候因子存在一定的相关性,遗传参数Ne、He及I与经度呈显著负相关(P<0.05),与海拔呈现极显著正相关(P<0.01),与年均温呈现显著负相关(P<0.05)。
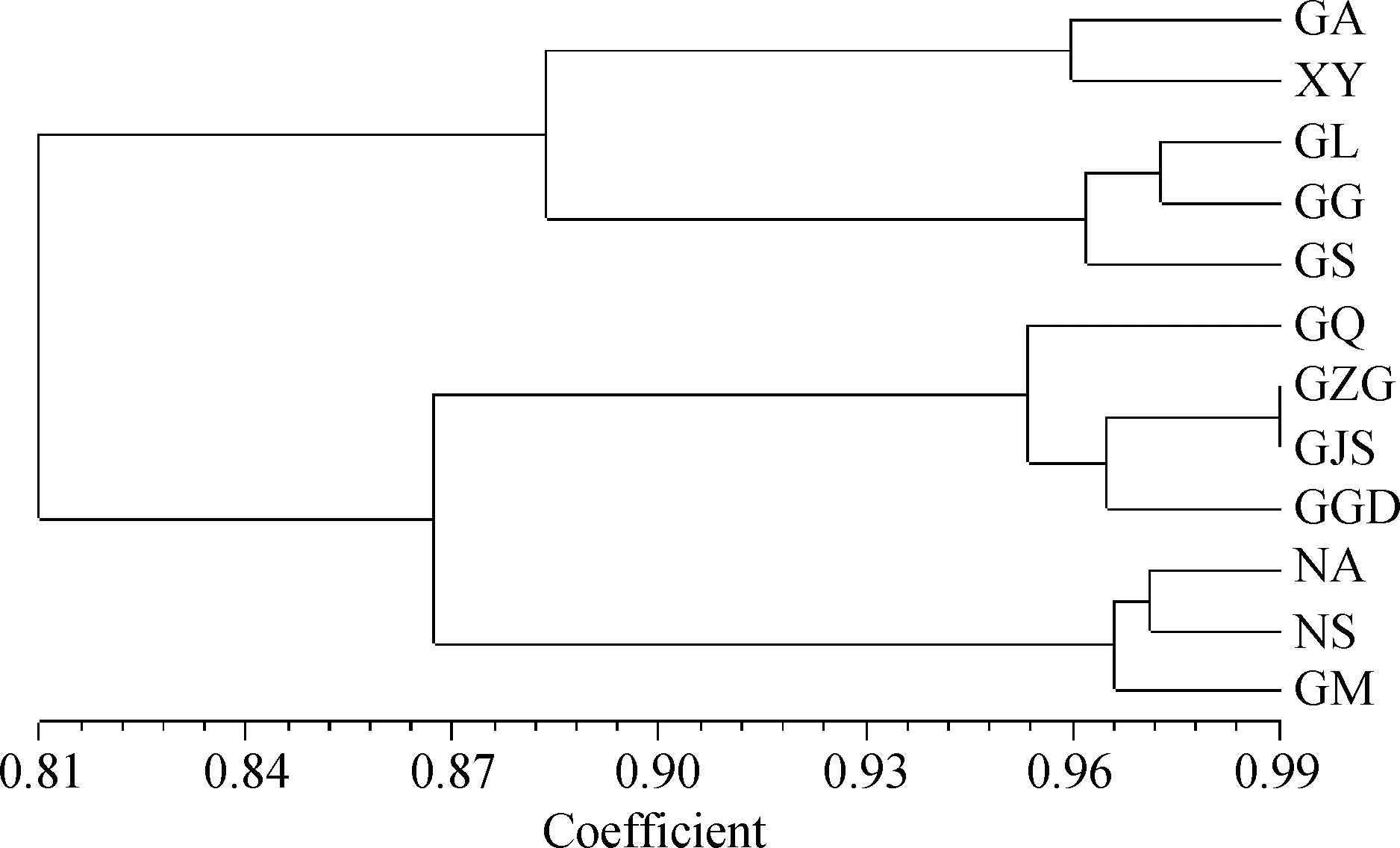
图1 基于ISSR的12个裸果木种群的UPGMA聚类图Fig.1 UPGMA dendrogram of 12 Gymnocarpos przewalskii clusters based on ISSR

表6 裸果木遗传参数与地理气候因子相关性分析Table 6 Correlation analysis between geographic climatic factors and genetic diversity of Gymnocarpos przewalskii
注: *表示P<0.05水平;**表示P<0.01水平
Note: *indicates the diffence at the 0.05 level;** indicates the diffence at the 0.01 level
3 讨论与结论
3.1 裸果木种群遗传多样性特点
遗传多样性水平高低与多因子有关,如分布区、生境的变化、繁育系统、个体细胞突变、奠基者效应、更新世产生的物种遗留区等[17]。荒漠濒危植物因繁殖方式、分布大小及生长环境的限制,如绵刺(Potaniniamongolica)、蒙古扁桃(Amygdalusmongolica)等常常表现为相对较低的遗传多样性水平[18-19]。然而也有研究发现濒危或分布狭窄的物种由于区系的古老性表现出较高的遗传多样性水平,如四合木(Tetraenamongolica)及长叶红砂(Reaumuriatrigyna)等[20-21]。Nybom等认为期望杂合度(He)和Shannon信息指数(I)更为重要,注重物种在种群水平的遗传多样性[22],而Hamrick等认为多态位点百分率(P)和期望杂合度(He)比较重要,注重物种水平的多样性[23]。本研究结果表明不同种群裸果木平均多态位点百分率(PPL)为69.93%。种群间He平均值为0.2418;I平均值为0.3629。裸果木总的遗传多样性(Ht)为0.3452,而种群内的遗传多样性(Hs)为0.2489,综合分析裸果木遗传多样性较高,维持着较高的遗传变异,这与贾舒雯等对中国西北干旱区裸果木遗传多样性分析结果相同[24]。一方面裸果木以有性繁殖为主,世代周期较长,为异花授粉植物,具虫媒及风媒传粉特点[8]。另一方面裸果木为古地中海孑遗种,起源于第三纪,末次间冰期裸果木可能分布范围较广,但受冰期的影响使得当时大部分植物消失[25],然而部分遗留下的裸果木在历史的变迁过程中保存至今,种群为适应环境的变化,积累丰富了遗传变异,从而可能使得裸果木存在较高的遗传多样性水平,推断裸果木存在较高的遗传多样性可能与其生物学特性、演化历史和分布区生态因子的特殊性分不开。
3.2 裸果木种群遗传结构与分化特点
种群的遗传结构是一个物种最基本的特征,基因流被认为是种群遗传结构均质化的主要因素之一。基因流与种群扩散、遗传有效性种群扩散有关,具有较高基因流的物种往往比基因流较低物种遗传分化小[26]。Slatkin认为Nm<1,表明种群间差异显著,基因流不足以抵制种群内因遗传漂变而引起的种群分化,若Nm>1表明种群之间有一定频率的基因流,遗传物质的交换可以降低种群内的遗传漂变,可以在一定程度上减少种群间的强烈分化[27]。研究发现裸果木的基因流为1.2920,种群间存在较高的基因流,种群间的遗传分化小,遗传变异在种群间均衡分布。马松梅采用cpDNA标记法对裸果木的遗传结构进行研究表明,中国西北地区的干旱和沙漠的扩展造成裸果木片段化[28]。本研究发现引发裸果木基因流较大的原因可能有以下几方面:从繁殖生物学角度看,裸果木繁育系统属兼性异交类型,异交为主,自交为辅,异交导致裸果木的基因流增大;从传粉生态学角度看,裸果木兼有风媒和虫媒传粉,盛花期裸果木花期开花量较大[8],传粉昆虫较多,使得基因在较多个体间交流。此外有学者对喜马拉雅红豆杉(Taxuswallichiana)[29]和桃儿七(Sinopodophyllumhexandrum)[30]的遗传特性进行研究,结果表明高原隆升形成的主要山系阻碍植物群体间的基因流,促进种群间植物异域分化。本研究分布区位于河西走廊等地,种群沿着祁连山分布,不存在山系间的隔离,各种群间地理距离较近,种群间的地理隔离因子不强烈,存在较大的基因交流。
3.3 地理气候因子对裸果木遗传多样性的影响
植物种群进化过程中,突变与选择都是重要的进化驱动力,然而一些极端环境或生态异质较大环境中,环境选择压力和变异方向尤为重要。地理和环境因子共同作用可加强植物种群的遗传分化[31]。如Wu等对穗状狐尾藻(Myriophyllumspicatum)遗传格局进行研究发现,区域尺度主要受环境要素制约,地理要素显得更重要[32]。遗传多样性与物种多样性在纬度梯度分布上的耦合暗示着可能存在一些共同的驱动因素[33],导致遗传多样性与物种多样性在纬度梯度上协同变化。对荒漠孑遗植物红砂(Reaumuriasoongarica)的遗传学研究表明,自2.96万年以来,亚洲季风气候促进了生境片断化和物种分化,同时冬季风促进了物种纬向的基因流[34]。本研究发现裸果木种群经度相对与纬度对遗传多样性影响较大,样品采集中经度的变化范围为94°~105°,经度跨越较大,可能引发不同地理间的差异较大,进而影响裸果木种群的遗传多样性水平。裸果木遗传与地理距离关系存在一定的相关性,从裸果木的分布区上可以看出,甘肃省阿克塞、甘肃省肃北县位于裸果木分布的中心地带,其种群西北的分布主要沿着天山山脉延伸分布于新疆部分区域,而向东部分布沿着祁连山分布于河西走廊东部及阴山山脉的内蒙古西部。从中心区向外不断扩展使得种群间的基因流变小,遗传水平随着地理间距离向两边延伸导致遗传水平降低。全球变化下温度成为一个直接的选择因子,这种选择压力也表现在物种的遗传变异上,即和温度相关联位点(适应性基因)的频率发生相关变化。如Jump等对不同年龄级个体欧洲水青冈(Fagussylcatica)的遗传多样性进行研究发现,其中一个位点的频率变化和温度有较高的相关性,表明植物为适应环境而产生变化[35]。Semagn等对Phytolaccadodecandra遗传分化研究,发现种群与温度、海拔以及生态地理因素显著相关,提出遗传变异是适应以上特征的基础[36]。本研究发现裸果木遗传多样性与海拔及温度有关,高海拔地区温度高,降水低,风力较大,在一定程度加速花粉及果实的散布,加强种群间基因交流,导致遗传多样性水平升高。环境与植物作为一个整体系统,各种因子之间相互联系、共同作用,本文仅分析地理及气候因子与遗传多样性的关系,有关多因子的综合作用还有待进一步深入探究。
猜你喜欢
学苑创造·A版(2025年2期)2025-01-14 00:00:00
今日农业(2022年15期)2022-09-20 06:54:16
腹腔镜外科杂志(2022年7期)2022-08-16 04:31:06
机械研究与应用(2022年3期)2022-07-25 05:58:34
甘肃林业(2022年1期)2022-02-19 02:35:08
特区文学(2020年2期)2020-09-10 07:22:44
中小学校长(2020年6期)2020-08-07 10:54:36
药学研究(2019年4期)2019-05-15 02:44:30
广西林业科学(2016年2期)2016-03-20 05:53:37
当代畜禽养殖业(2014年10期)2014-02-27 07:59:49
