
不同倍性小麦光合日变化的研究
2018-07-26 02:42赵鹏涛赵小光张耀文翟周平关周博罗红炼
江西农业学报 2018年7期
赵鹏涛,赵小光,张耀文,翟周平,关周博,罗红炼
(陕西省杂交油菜研究中心 小麦研究室,陕西 杨凌 712100)
异源多倍体化是高等植物重要的进化过程,在天然物种和农作物中广泛存在[1];自然界中真正的二倍体物种极少,在外界选择压力下,逐渐向着多倍体进化[2]。近年来通过对芸苔属、小麦、拟南芥、棉花等植物的研究发现,在异源多倍体形成早期经常伴随着与孟德尔法则不一致的基因组变化,表现在遗传、转录、表达水平等上[3-5],它们有助于多倍体新物种的形成,同时也为植物的长期进化提供了可能的原材料。多倍体小麦是通过异源多倍体化方式进化和培育的典型,特别是六倍体普通小麦是世界范围种植最广泛的、最重要的粮食作物之一。研究表明,在小麦从二倍体到八倍体的演化中,小麦产量显著增加。作为大多数栽培小麦的起源,古老的小麦基因型是在小麦改良中获得新的遗传多样性最有价值、最易得到的来源[6],从而使异源多倍体小麦对复杂多样的环境条件常表现出更广泛的适应性和其它有利的农艺性状。
光合作用是植物进行物质积累与生理代谢的基础,其效率的高低与产量潜力发挥以及品质优劣密切相关,因此研究作物的光合作用对农业育种工作具有重要的意义[7]。光合作用是一个复杂的生理过程,既与作物自身因素如光合反应酶的活性、叶片叶绿素含量、植株发育时期密切相关,同时又受光照强度、空气温度、大气CO2浓度等外界因素的影响。因此,我们依靠仪器测定的作物瞬时净光合速率处在一个动态的变化中,并不能真正衡量作物光合能力的强弱,而光合参数的日变化可以更直观和综合地反映出作物自身的光合能力及其环境因素的变化[8-9]。近年来国内外不少学者对油菜、谷子、水稻、玉米、大豆、花生、番茄[10-16]等作物的光合日变化进行了大量的研究,发现作物的净光合速率日变化有很强的规律性,其变化曲线多为双峰型(C3植物)或单峰型(C4植物)。尽管国内外对小麦的光合日变化也进行了较多的研究[17-20],但是主要围绕比较单一的实验材料进行的,而对小麦不同倍性进化材料的光合日变化研究较少,仅有关于八倍体与六倍体普通小麦光合日变化的比较研究[21]。本实验利用便携式Li-6400光合作用测定仪在小麦灌浆期分别测定了二倍体、四倍体、六倍体、八倍体小麦旗叶的净光合速率、蒸腾速率等光合生理参数,以及光合有效辐射、大气温度等环境因子的日变化,以期能够对小麦产量形成关键期的光合性状有全面的了解,从而为小麦的高光效育种提供进一步的理论支持。
1 材料与方法
1.1 试验材料
试验材料二倍体节节麦(Aegilopstauschii)、四倍体硬粒小麦(T.durum)、六倍体普通小麦(T.aestivum)及八倍体小黑麦(Triticale)均由陕西省杂交油菜研究中心小麦研究室提供。试验于2016~2017年在陕西省杨凌示范区小麦试验田进行,田间试验采用随机区组设计,每小区种植5行小麦,行长2.0 m,行距0.25 m;每个倍体材料重复3次,田间种植与管理同国家小麦区域试验[22]。
1.2 试验方法
1.2.1 测定方法 在小麦的灌浆期,选择生长健壮、均匀一致的单株,用Li-6400便携式光合作用测定仪对受光方向一致的旗叶进行活体测定,每个倍体材料都测定10株。测定时选用透明叶室,空气流速设为500 mol/s,温度设置为大气温度。从早上8:00开始测定,直到18:00结束,每小时重复测定1次。叶片夹样部位选择旗叶的中段;如果叶片宽度小于2 cm,不能充满整个叶室,则光合速率等的重计算方法参见文献[23]。
1.2.2 测定指标 测定的指标有:净光合速率Pn[μmol/(m2·s)]、气孔导度Gs[mmol/(m2·s)]、胞间CO2浓度Ci(μmol/mol)、蒸腾速率Tr[mmol/(m2·s)]、光合有效辐射PAR[μmol/(m2·s)]、空气温度Ta(℃)、大气CO2浓度Ca(μmol/mol)、空气相对湿度RH(%)等;水分利用效率WUE的计算公式为:WUE=Pn/Tr[24]。
1.2.3 数据的整理与计算 利用Excel 2003对试验数据进行初步整理,用SPSS 19.0数据统计软件进行统计分析。
2 结果与分析
2.1 环境因子的日变化特征
作物的光合作用日变化在很大程度上受外界环境因子的影响,其中起主要作用的有光合有效辐射(PAR)、空气温度(Ta)、大气CO2浓度(Ca)和空气相对湿度(RH)[23]。从图1-A中可以看出:PAR在一日内的变化表现为明显的单峰曲线,从早晨开始逐渐上升,到12:00时达到最高值,然后逐渐下降;Ta的日变化趋势与PAR一致,也表现为单峰曲线,不过变化的幅度较PAR小,且峰值出现的时间比PAR延迟,出现在14:00。在图1-B中,Ca的日变化规律与前两者明显不同,从早晨开始急剧下降,在10:00以后下降趋势缓慢,且在一日内始终处于下降中。这主要是由于整个生态系统中绿色植物的光合作用对大气中CO2的消耗,从而造成Ca的持续下降。而RH的日变化则与PAR和Ta相反,表现为倒抛物线,在14:00出现最低值,它的变化规律主要受Ta的影响,即温度越高,空气湿度越低。

图1 不同环境因子的日变化
2.2 不同倍体小麦光合参数的日变化比较
2.2.1 不同多倍体小麦净光合速率的日变化 净光合速率(Pn)是作物的一系列生理和生态因子综合作用的瞬时表现形式,Pn日变化可以更直观地反映出作物利用光能的强弱,常被用来作为分析作物生长及产量形成的主要参考标准。由图2可以看出,4种倍体的小麦均存在光午休现象,且上午的光合效率高于下午。在上午,二倍体和八倍体小麦的Pn高于四倍体和六倍体的;在下午,二倍体小麦的Pn下降比较明显,低于其他3种材料的,而八倍体的Pn仍然高于其他几个多倍体材料的。另外,二倍体、四倍体、八倍体的Pn日变化第一个峰值均出现在10:00,而六倍体的峰值则出现在11:00;二倍体、四倍体的光午休点出现在14:00,而六倍体和八倍体则出现在15:00。因此,随着小麦倍体的不断进化,高倍体小麦材料对光午休的抵抗能力明显增强,其下午的光合效率比低倍体材料也有明显的提高。
2.2.2 不同多倍体小麦气孔导度和蒸腾速率的日变化 作物的气孔导度(Gs)和蒸腾速率(Tr)都与气孔的开放程度密切相关,而气孔的开闭又受到一日内温度和湿度等外界因子的调节。从图3-A可以看出,4种倍体小麦的Gs日变化规律与Pn的日变化规律基本一致,都表现为双峰曲线。从早上开始,随着光照的刺激和气温的升高,小麦叶片的Gs逐渐升高,二倍体和四倍体在10:00达到峰值,而六倍体和八倍体在11:00达到峰值。在中午时刻,由于温度过高,植物为了避免过量失水,气孔部分关闭,所以气孔导度在中午出现明显的下降,二倍体和四倍体在13:00出现最低值,而六倍体和八倍体在14:00出现最低值。

图2 不同多倍体小麦净光合速率的日变化
从图3-B可以看出: Tr的日变化趋势与Gs相似,4种多倍体小麦的Tr日变化均表现为双峰曲线,即在11:00时Tr达到最大值;随着气孔的关闭,Tr在13:00均达到最低值;然后又逐渐升高,在15:00达到第二峰值。与Pn日变化不同的是,Gs和Tr的日变化在上午和下午差异不是很大,而4种倍体小麦的Pn在上午的平均值都明显高于下午的。
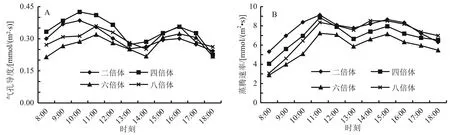
图3 不同多倍体小麦气孔导度和蒸腾速率的日变化
2.2.3 不同多倍体小麦胞间CO2浓度的日变化 胞间CO2浓度(Ci)作为植物进行光合作用的直接碳源,其数值高低直接影响着作物光合效率的高低。从图4可以看出,不同多倍体小麦的Ci日变化趋势基本一致,均表现为倒抛物线。从早上开始,随着小麦光合作用的增强,Ci一直处于下降的趋势。到了中午时刻,尽管小麦的Pn由于受光午休影响开始下降,光合作用对Ci的消耗降低,然而叶片的气孔大部分关闭,从外界交换来的CO2也处于减少中,在两者的共同作用下,Ci继续下降,一直到14:00达到最低值;此后随着光午休的减弱,胞间CO2浓度逐渐升高。此外,六倍体和八倍体在早上各时间点的Ci低于二倍体和四倍体的,表明前两者对碳源的利用效率要高于后两者的。
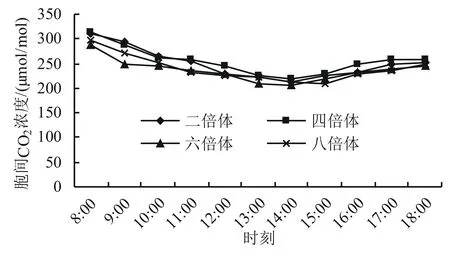
图4 不同多倍体小麦胞间CO2浓度的日变化
2.2.4 不同多倍体小麦水分利用效率的日变化 从图5可以看出,不同多倍体小麦的水分利用效率(WUE)日变化趋势基本一致,从早上开始一直处于下降中,在中午过后下降趋势基本平缓。二倍体和四倍体小麦的WUE在一日中一直低于六倍体和八倍体的,六倍体小麦的WUE在早上低于八倍体,而在下午高于八倍体,表明随着小麦倍体的进化,多倍体材料对水分的利用效率得到了提高,从而更能适应高温和干旱的环境。
2.3 小麦净光合速率与影响因子的相关性分析
对不同多倍体小麦一日内各时间点的净光合速率Pn与相关气体交换参数以及环境因子进行了相关性分析,结果(表1)表明:4种多倍体小麦的Pn在一日内与Gs呈极显著正相关(P<0.01),因此Pn的日变化主要受到Gs的调节。另外二倍体和四倍体的Pn还与RH和WUE显著相关(P<0.05),说明这两种倍体小麦Pn的日变化在很大程度上受到外界和自身水分的调节。而六倍体和八倍体小麦的Pn与环境因子均无显著性相关,说明两者对环境的抗逆性较强,光合能力主要与自身的生理代谢相关。
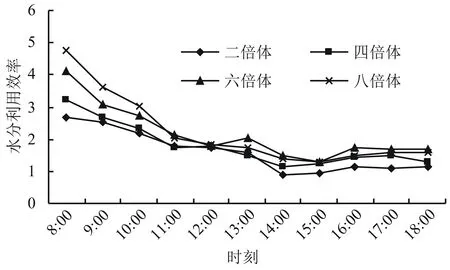
图5 不同多倍体小麦水分利用效率的日变化

表1 不同倍体小麦净光合速率与其影响因子间的相关系数
注:“*”表示在0.05水平上显著相关;“**”表示在0.01水平上极显著相关。
3 讨论
异源多倍体化是形成新物种的一种有效手段,已经在育种上得到了广泛的应用,形成了丰富的种质资源和新品种。在小麦上,六倍体小麦已经大面积种植,八倍体小麦在某些农艺和品质性状上也表现出独有的优势,逐渐被小麦育种家重视。本试验发现:小麦从二倍体进化到八倍体,其Pn得到了逐步提高,对CO2和水分的利用率也得到了提高,对环境的适应性明显增强;六倍体和八倍体小麦的光合效率比二倍体和四倍体小麦有了明显的改良。这与魏爱丽等的报道结果[21]一致。另外八倍体小麦材料的光午休时间较其他3种多倍体材料延迟且缩短,因此我们可以借鉴该方法减弱小麦的光午休现象,进一步提高小麦的总体光合效率。由中国科学院遗传与发育生物学研究所选育的杂交种小麦“小偃101”在中午强光下的光合效率超过其双亲,从而有效地提高了该作物的最终产量,在多点预先试验中比对照增产10%左右[25]。
目前,我国的小麦单产量已经取得了巨大的突破,在理想的条件下可以达到7500 kg/hm2,然而距离国外的最高产量纪录还有不小的差距,所以我国的小麦产量还有很大的提升潜力。因此,在传统的抗逆、品质、早熟等育种的基础上,应辅助高光效育种手段,提高亲本资源的光合能力,通过杂交改良提高后代对光的截获和利用能力,降低光午休和光抑制等负效应,最大程度地将光能转化为小麦的化学能贮存起来,从而最终提高小麦的产量。
猜你喜欢
西北农业学报(2022年5期)2022-06-07
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
山西农业科学(2021年3期)2021-12-06
东北林业大学学报(2021年12期)2021-02-10
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
江苏农业科学(2014年9期)2014-11-15
