
不同倍性葡萄种子萌芽力研究
2018-07-25 03:43李伟东程杰山高营营范培格
中外葡萄与葡萄酒 2018年4期
李伟东,程杰山,高营营*,范培格*
(1. 中国科学院植物研究所,北京 100093;2. 鲁东大学农学院,山东烟台 264000)
葡萄是我国五大果树之一,据国家统计局数据,2015年我国葡萄栽培面积79.9万 hm2,产量1366.9万 t,栽培面积和产量分居世界第二位和第一位,葡萄产业在我国农业经济发展中占据重要地位。但我国葡萄实际栽培中还存在旱寒、盐碱等不利因素[1],所以抗逆、加工及鲜食新品种的选育是我国葡萄育种的重要方向。目前有性杂交育种仍是国内外葡萄新品种选育的最主要方法[2],获得较多的杂种苗是新品种选育的首要条件,而提高杂交种子的萌发率对于有性杂交至关重要。在多种植物中的研究发现,变温处理可以打破种子休眠并提高种子萌发率[3-7]。
果树多倍体育种已成为果树新品种选育的重要技术手段并具有广阔的前景[8]。为获得高抗逆、大粒优质新种质,本实验室前期利用高抗逆品种‘北红’‘北玫’染色体加倍获得其同源四倍体植株,为充分利用高抗逆资源,特开展其二倍体与同源四倍体种子萌发试验,为今后葡萄多倍体育种提供理论依据。
1 材料与方法
1.1 材料
于2018年3~4月在中国科学院植物研究所进行,所用种子于2017年10上旬在中国科学院植物研究所北区葡萄品种圃内采收。选用‘北红’‘北玫’及‘四倍体北红’‘四倍体北玫’种子各200粒。
1.2 方法
1.2.1 种子采集
采集各品种充分成熟的葡萄果实,冰箱4 ℃保存,在播种前人工剥皮去除果肉取出种子,挑选饱满种子洗净,做好标签。
1.2.2 种子处理与催芽
将种子先用洗衣粉清洗干净,无菌水冲洗,然后75%乙醇浸泡10 min,无菌水冲洗,再用5%次氯酸钠溶液浸泡15 min,用无菌水冲洗干净。
将各品种种子平均分成两组(甲组和乙组),每组100粒。每组种子平均分到3个置有三层湿滤纸的培养皿中(A,B,C),用封口膜封住,贴好标签,作为3个生物学重复。
将甲组种子放入25 ℃、24 h恒温培养箱中用锡纸遮盖进行暗催芽,乙组种子同样用锡纸遮盖进行暗催芽,放入恒温库中进行30 ℃,12 h/20 ℃,12 h的变温处理。每天定时观察记录发芽种子数,并适时补充水分和换气。以种子开裂露白或长根为萌芽标准[9],待种子发芽数不再增加时终止调查统计。
1.3 统计数据
利用Excel软件进行数据处理,使用SPSS 21.0中的Dancun's法进行差异显著性分析(P<0.05)。
发芽率计算公式如下:
发芽率/%=(萌发种子总数/供试种子总数)×100
日发芽率/%=(每日发芽种子数/供试种子总数)×100
2 结果分析
2.1 不同萌发条件下‘北红’‘北玫’及其四倍体实生种子发芽率

表1 ‘北红’及‘四倍体北红’发芽率情况Table 1 The germination rate of 'Beihong' and 'Tetraploid Beihong'
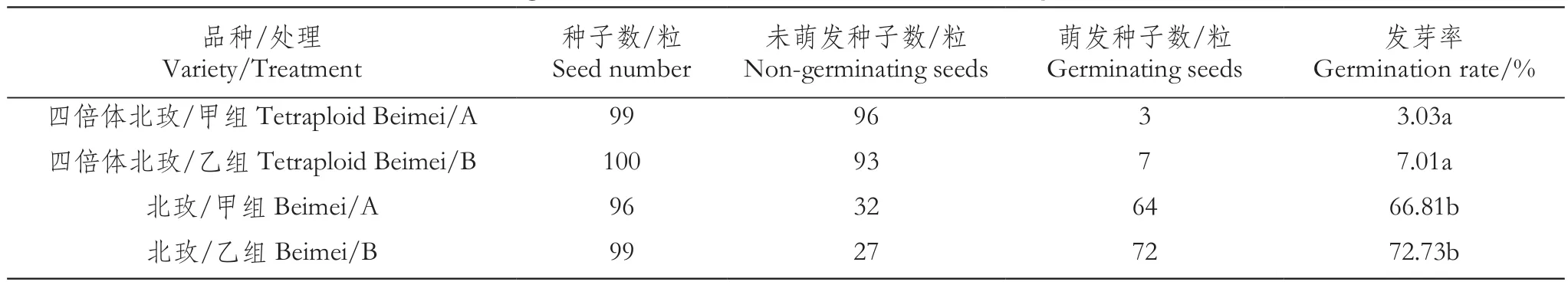
表2 ‘北玫’及‘四倍体北玫’发芽率情况Table 2 The germination rate of 'Beimei' and 'Tetraploid Beimei'
由表1与表2可看出,两种萌发环境处理下的二倍体‘北红’‘北玫’种子的萌芽率均明显高于相应同源四倍体种子,‘北红’两种处理条件下种子萌发率均在90%以上,而其同源四倍体种子的萌发率在两种处理条件下均很低,分别为23%和37%。‘北玫’与其同源四倍体种子的萌发情况与‘北红’类似,只不过‘北玫’种子的萌发率低于‘北红’,‘四倍体北玫’种子的萌发率更低,最高仅为7%。
除‘北红’外,‘北玫’及‘四倍体北红’‘四倍体北玫’在变温处理下种子的萌发率数值与恒温处理下相比,虽未达到显著差异水平,但数值略有增加。
2.2 不同萌发条件下日发芽率

图1 ‘北红’及‘四倍体北红’日发芽率Figure 1 The daily germination rate of 'Beihong' and'Tetraploid Beihong'

图2 ‘北玫’及‘四倍体北玫’日发芽率Figure 2 The daily germination rate of 'Beimei' and 'Tetraploid Beimei'
各参试品种的种子日萌发率达到峰值所需的催芽时间用催芽后天数[10]表征。由图1和图2可知,各参试品种在两种处理下日萌发率达到峰值所用时间分别为:甲组‘北红’7 d,‘四倍体北红’13 d;乙组‘北红’12 d,‘四倍体北红’23 d;甲组‘北玫’9 d,‘四倍体北玫’17 d;乙组‘北玫’12 d,‘四倍体北玫’21 d。相同环境条件下,二倍体种子达到日萌发率峰值的速度均快于其同源四倍体,‘北红’比相应同源四倍体至少提前6 d,‘北玫’至少提前8 d。而变温处理使二倍体和四倍体的日萌发率峰值均推迟,变温使‘北红’种子日萌发峰值推迟了5 d,‘北玫’种子推迟了3 d,‘四倍体北红’推迟了10 d,‘四倍体北玫’推迟了5 d。
本试验中各参试品种的种子从开始发芽到结束发芽所用时间(间隔天数):甲组‘北红’14 d,‘四倍体北红’25 d;乙组‘北红’16 d,‘四倍体北红’21 d;甲组‘北玫’15 d,‘四倍体北玫’18 d;乙组‘北玫’14 d,‘四倍体北玫’12 d。总体来看,不同条件下二倍体品种发芽更为集中,间隔天数为14~16 d,‘四倍体北玫’比‘四倍体北红’萌发时间较短。除‘北红’外,其余品种在变温条件下的间隔天数短于恒温条件下的天数。

图3 ‘北红’及‘四倍体北红’累计发芽率Figure 3 The accumulated germination rate of 'Beihong' and'Tetraploid Beihong'

图4 ‘北玫’及‘四倍体北玫’累计发芽率Figure 4 The accumulated germination rate of 'Beimei' and'Tetraploid Beimei'
2.3 葡萄种子累计发芽率
由图3与图4可知,无论是恒温处理还是变温处理,‘北红’和‘北玫’二倍体种子发芽快,且更为集中,34 d的试验中,二倍体‘北红’(甲组)在催芽开始后18 d即达到最大值94.95%,其四倍体种子(乙组)试验期间陆续萌芽,到34 d仅达到37.56%。相同倍性种子在不同条件下表现出相似的萌发趋势,变温处理下各参试品种在早期的萌芽率均低于恒温处理的,但在后期除‘北红’外,‘北玫’‘四倍体北玫’和‘四倍体北红’的萌芽率均有所上升,超过了恒温处理。
3 讨论与结论
多倍体材料因为染色体的加倍效应,在表型上与二倍体差异较大,如花粉粒大并且形状不规则,胚珠异常,发芽率低,种子较大、较少且部分发育不良,果实成熟期提前等[11]。通过本试验也得出结论,不同染色体倍性葡萄种子的萌发率存在较大差异,二倍体种子的萌发率明显高于同源四倍体种子。这与李丽霞等[11]对葡萄多倍体研究和张巍巍等[12]对四倍体黑芥种子萌发率远低于二倍体种子研究一致。种子的萌发受诸多因素影响,如种子的本身特性、外界环境条件、成熟度、内源激素、层积处理条件等[13]。有研究报道,二倍体葡萄种子较小,种皮相对较薄,水分容易进入,相对容易萌发,四倍体种子相对较大,种皮厚且坚硬,不易萌发[11]。另外相较于二倍体,生产中四倍体种子胚败育的情况比较常见[14]。此外,种子内源激素影响种子萌发[15-17],前人研究表明,四倍体植株内源激素含量不同于其二倍体[18-19],我们推测本研究中可能是四倍体种子内源激素含量的不同导致了种子发芽率较低。对于导致二倍体与同源四倍体种子萌发率差异的原因有待进一步深入研究,以便寻找提高四倍体萌芽率的技术措施。据报道,变温调节可以促进多种植物种子的萌芽[3-7],在本试验中,虽然变温处理下‘北玫’‘四倍体北玫’和‘四倍体北红’的萌芽率高于恒温处理,但不存在显著性差异。变温处理虽然推迟了种子开始萌发的时间和达到日萌发峰值所需的天数,但缩短了从萌发开始到结束的天数,发芽更为集中。所以我们建议如果以发芽速度为目的催芽,可以考虑使用恒温条件,如果以最终发芽率或发芽集中为目的催芽,可以考虑使用变温条件。
猜你喜欢
西北农业学报(2022年5期)2022-06-07
沈阳农业大学学报(2022年2期)2022-06-04
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年12期)2022-01-14
东北林业大学学报(2021年12期)2021-02-10
发电技术(2020年3期)2020-06-29
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
江苏农业科学(2016年1期)2017-05-17
江苏农业科学(2014年9期)2014-11-15
