
高产乙酸酯酿酒酵母菌种的选育与提高白酒酯香物质含量的研究
2018-07-25 03:09张翠英崔丹瑶马红霞
酿酒科技 2018年7期
张翠英,崔丹瑶,李 维,张 雨,马红霞
(工业发酵微生物教育部重点实验室,天津市工业微生物重点实验室,天津科技大学生物工程学院,天津300457)
酯香物质是白酒中的重要风味成分,是白酒香气的主要来源[1-2]。酯香物质一般具有强烈的果香味或溶剂味,穿透力强,且阈值较低。白酒中的酯类主要有乙酸乙酯、乳酸乙酯、丁酸乙酯、己酸乙酯、乙酸异丁酯、乙酸异戊酯、丙酸乙酯、辛酸乙酯、乙酸苯乙酯、苯乙酸乙酯、丁二酸二乙酯等。我国高档白酒中一般酯含量较高,但中低档白酒中酯含量相对较低。如何适当提高白酒中有积极作用的酯类物质含量,一直是我国白酒行业和相关科研人员研究的热点。
乙酸乙酯、乙酸异丁酯、乙酸异戊酯、乙酸苯乙酯属于乙酸酯类,是白酒中最重要的一类活性香气酯。各种香型白酒中乙酸乙酯占总酯量的比例均很高,尤其是在凤香型、清香型、酱香型白酒中乙酸乙酯占总酯量的比例最高,西凤酒、汾酒、茅台酒中分别达60.14%、53.48%、38.28%。在各种香型高档白酒中乙酸乙酯的含量不同,但都很高,汾酒、茅台酒、习酒中乙酸乙酯含量分别为2627.9 mg/L[3]、1435.54~1769.81 mg/L[4]、1903.28 mg/L[5]。
1 酿酒酵母中乙酸酯的代谢机制
1.1 乙酸酯的合成
酯的合成途径主要有两种,化学合成和生物合成。化学合成速度慢,合成量低,底物浓度要求高。在白酒发酵过程中,乙酸酯主要是由醇酰基转移酶[6]或者其他酯合成酶在酿酒酵母细胞内生物催化合成的(图1)[7]。催化反应需要能量来连接酰基辅酶A和基质。乙酰辅酶A是酿酒酵母中含量最丰富的酰基辅酶A,既可以来源于丙酮酸的氧化脱羧,也可以由乙酸直接活化得到。大部分的乙酰辅酶A是由丙酮酸的氧化脱羧产生的。

图1 酵母菌中乙酸乙酯的生物合成途径[7]
醇乙酰基转移酶(AATase)是酵母合成乙酸酯类(如乙酸乙酯、乙酸异戊酯等)的关键酶,催化醇类和乙酰辅酶A生成乙酸酯。早期研究发现[8-12],有3种不同的醇乙酰基转移酶AATaseⅠ、Lg-AATaseⅠ和AATaseⅡ,分别由ATF1、Lg-ATF1和ATF2编码。ATF1和ATF2在酿酒酵母和啤酒酵母中均存在,而Lg-ATF1只存在于啤酒酵母(巴斯德酵母)中。Malcorps和Dufour[13]通过实验得到了纯化19000倍的AATase I,并估算出其分子量为57 kDa±3 kDa。研究证实,该酶的最适pH8.0,相对于乙酸异戊酯和乙酸乙酯的Km值是25 mmol/L和45 mmol/L,对异戊醇则是25 mmol/L。通过对酿酒酵母基因组进行蛋白印迹分析发现,每个酿酒酵母单倍体基因组都含有一个ATF1基因。研究表明,ATF1的同源基因Lg-ATF1编码的蛋白质Lg-Atf1p与Atf1p一致性高达81%。Atf2p与Atf1p的一致性为36.9%[11]。
酿酒酵母乙酸酯代谢途径[14]见图2。大量研究结果证明,酿酒酵母AATase编码基因ATF1和ATF2的表达水平密切影响乙酸乙酯和乙酸异戊酯的合成。除此之外,ATF1和ATF2基因编码的酯合成酶还与很多微量挥发酯的生成有关,如乙酸丙酯,乙酸异丁酯,乙酸戊酯,乙酸己酯,乙酸庚酯,乙酸辛酯和乙酸苯乙酯。相比而言,ATF1基因的作用要大于ATF2基因的作用。
ATF1基因的表达水平受不饱和脂肪酸和氧气抑制,受蛋白激酶SCh9和蛋白激酶A调控[15-18]。这些激酶通过改变碳、氮和磷酸盐的含量来对基因的转录调节发挥重要调控作用。Sch9主要调控参与细胞生长、糖和海藻糖代谢的基因。不同不饱和脂肪酸的影响与该脂肪酸的解链温度密切相关,越低解链温度的不饱和脂肪酸对ATF1的转录抑制越强烈。除了氧气和不饱和脂肪酸的影响外,碳和氮的缺失也会对ATF1的转录产生抑制。这种情况下参与乙酸酯合成的酶活性下降。研究表明,ATF1基因的表达还受温度和酒度的影响。

图2 酿酒酵母乙酸酯代谢途径[14
最近一篇研究报道,在酵母菌中发现了一个新的醇乙酰基转移酶家族Eat1,可能是酵母菌中乙酸乙酯合成的主要酶[7]。在海洋嗜杀酵母(Wickerhamomyces anomalus)产乙酸乙酯过程中EAT1基因显著上调;敲除乳酸克鲁维酵母(Kluyveromyces lactis)的EAT1基因,乙酸乙酯产生量降低80%。这个酶在所有产乙酸乙酯的酵母菌中都存在编码基因序列[7]。
1.2 乙酸酯的水解
由酿酒酵母酯合成酶催化合成的酯还会被酵母产生的酯水解酶催化水解[19]。与酯合成相比,国内外关于酯水解的研究较少。研究证实,酯合成酶与水解酶的平衡性是酯最终浓度的决定性因素[20]。酯酶由许多不同种类的水解酶组成,它们可以催化酯的分解,有时候也会促进酯之间的缔合。酯酶主要通过以下反应催化酯的分解:RCOOR+H2O→ROH+RCOOH。
对葡萄酒的研究发现,过表达IAH1基因可以导致乙酸乙酯、乙酸异戊酯和己酸乙酯的浓度显著降低[21]。对清酒的研究发现,IAH1基因缺失的清酒酵母菌株所产乙酸异戊酯的浓度远高于野生清酒酵母菌株,乙酸异丁酯的产量较野生清酒酵母菌株也有增加。因此,研究认为Iah1p可以水解乙酸异戊酯。
对酯水解酶的三维结构分析显示,有α-螺旋结构和β-片状结构两种水解酶。催化主要有3个重要元素组成:丝氨酸-天冬氨酸-组氨酸,在活性位点上通常具有这样一种共同序列:甘氨酸-X-丝氨酸-X-甘氨酸。这种三元催化系统还存在于其他一些酶中,如硫酯水解酶、蛋白酶、环氧化物水解酶等。研究证实,丝氨酰-组氨酸以及相似的二肽可以自行切割DNA和蛋白质。另外,很可能还存在其他一些酶与酯的水解有关如丝氨酸蛋白酶。
酶的活性很大程度上取决于乙酸酯主链的长度,而不是它的结构(直链或者支链)。如图3所示,Iah1p对乙酸异丁酯的亲和力和作用力更强[22]。

图3 Iah1p对乙酸酯类的底物特异性(设定对乙酸异丁酯的相对活性为100%)[22]
2 高产乙酸酯酿酒酵母菌种的选育
乙酸酯是酿酒酵母发酵产生的香气成分中最重要的一类,构成了高质量酒的果香成分。乙酸酯的浓度主要是受ATF1和ATF2基因的调控。酿酒酵母细胞内影响ATF1和ATF2基因表达的因素都会影响乙酸酯的终浓度。
通过基因敲除缺失酿酒酵母酯水解酶(Esterase)编码基因IAH1,发酵结果显示,重组菌株IY1与亲本菌株RY1相比,乙酸乙酯含量提高到47.4 mg/L,提高了46%;乙酸异丁酯、乙酸异戊酯以及相关高级醇(异丁醇、异戊醇)的产量均没有明显变化[23](表1)。研究表明酿酒酵母酯水解酶编码基因IAH1的缺失可以提高其乙酸乙酯的产量,而对乙酸异戊酯和乙酸异丁酯没有任何影响。

表1 突变株IY1与亲本菌株RY1酯醇产生量(mg/L)
选用PGK1作为启动子,利用同源重组技术,在过表达酿酒酵母编码醇乙酰基转移酶I的ATF1基因的同时,将酿酒酵母基因组中编码酯水解酶的IAH1基因敲除,获得了酿酒酵母基因工程菌EY1。与酿酒酵母亲本菌株RY1相比,在其他发酵性能不受影响的情况下,模拟半固态发酵后,异戊醇含量降低了约50%,乙酸乙酯含量提高了近20倍,乙酸异戊酯的含量提高到100 mg/L,乙酸异丁酯含量提高到5~7 mg/L(表2);模拟固态发酵后,总酯提高了4倍,其中乙酸乙酯提高了近35倍[24]。

表2 突变株EY1与亲本菌株RY1酯醇产生量(mg/L)
选用PGK1作为启动子,利用同源重组技术,在过表达酿酒酵母编码醇乙酰基转移酶II的ATF2基因的同时,将酿酒酵母基因组中编码酯水解酶的IAH1基因敲除,获得了酿酒酵母基因工程菌EY2。与酿酒酵母亲本菌株RY1相比,在其他发酵性能不受影响的情况下,模拟半固态发酵后,乙酸乙酯含量提高3.9倍,乙酸异戊酯的含量提高到26.68 mg/L,乙酸异丁酯含量提高到7.60 mg/L[25-26](表3)。
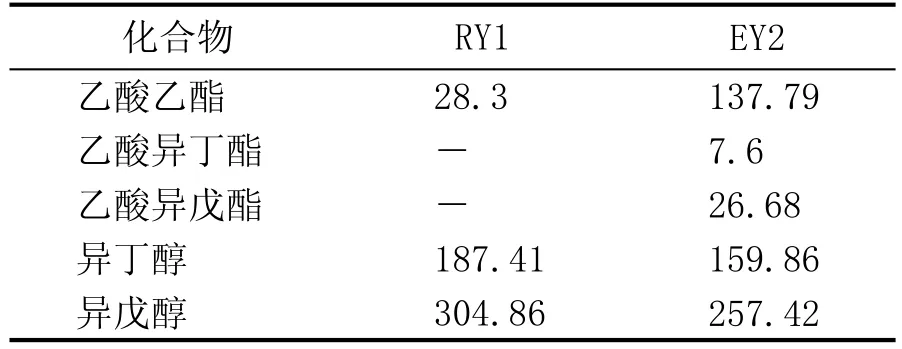
表3 突变株EY2与亲本菌株RY1酯醇产生量(mg/L)
选用强启动子PGK1将酿酒酵母中特有的编码醇乙酰基转移酶Lg-I的Lg-ATF1基因在酿酒酵母中高效表达,得到含有Lg-ATF1基因的酿酒酵母工程菌EY3。该菌在其他发酵性能不受影响的情况下,与亲本菌株RY1相比:模拟半固态发酵后,乙酸乙酯含量提高1.5倍,乙酸异戊酯的含量提高到8.66 mg/L[25-26](表4)。

表4 突变株EY3与亲本菌株RY1酯醇产生量(mg/L)
3 高产乙酸酯酿酒酵母在白酒中的应用
利用改良的高产酯酿酒酵母(1种酵母)取代目前复合生香酵母(1种酿酒酵母+4种生香酵母)生产芝麻香型白酒,可减化芝麻香型白酒生产工艺,提高生产效率。生产试验结果表明,理化指标、原酒色谱数据与原料出酒率与对照组无明显差异,感官品评结果,试验组较对照组芝麻香较成熟、清、带甜香味,较醇和(表5)。说明高产酯酿酒酵母可代替复合酵母生产芝麻香型白酒,且芝麻香基酒的典型性更好,优质酒率可提高30%以上。
利用高产酯低产高级醇酿酒酵母,进行清香型酒丢糟配粮发酵生产高酯调味酒试验[27-28]。发酵周期(6 d)缩短至原来的1/5,车间生产效率提高约8倍;乙酸乙酯和总酯含量明显提高,高级醇含量有所下降,清香更显突出;基酒中总酯含量高于5000 mg/L,乙乳比提高至3.0左右,适合于用来勾调夏季生产时乳酸乙酯含量偏高的基酒(表6)。

表5 原酒感官品评描述

表6 清香型白酒生产试验原酒主要成分比较(mg/L)
4 结论
乙酸酯是白酒中最重要的一类活性香气酯,对白酒产品风味与品质具有重要影响。在白酒发酵过程中,乙酸酯主要是在酵母细胞中由醇酰基转移酶或者其他酯合成酶催化合成的。大量研究结果证明酿酒酵母AATase编码基因ATF1的表达水平是影响乙酸乙酯和乙酸异戊酯合成的关键因素。最近也有研究报道在酵母菌中发现的新的醇乙酰基转移酶家族Eat1可能是酵母菌中乙酸乙酯合成的主要酶。由酿酒酵母酯合成酶催化合成的酯还会被酵母产生的酯水解酶(IAH1基因编码酯水解酶)催化水解。研究证实酯合成酶与水解酶的平衡性是酯最终浓度的决定性因素。
通过敲除编码酯水解酶的IAH1基因,过表达编码醇乙酰基转移酶的ATF1、ATF2、Lg-ATF1基因可以获得高产乙酸酯酿酒酵母菌株,乙酸乙酯含量分别提高了0.46倍、20倍、3.9倍、1.5倍;过表达ATF1、ATF2、Lg-ATF1基因菌株乙酸异戊酯的含量分别提高到100 mg/L、26.68 mg/L、8.66 mg/L。
选育的高产酯酿酒酵母菌株应用到白酒生产中,可显著改善白酒产品的风味和品质。芝麻香型白酒生产应用试验显示,高产酯酿酒酵母菌株的应用可减化芝麻香型白酒生产工艺,提高生产效率,且芝麻香基酒的典型性更好,优质酒率可提高30%以上。清香型酒丢糟配粮发酵生产试验显示,发酵周期大大缩短,车间生产效率提高;乙酸乙酯和总酯含量明显提高,高级醇含量有所下降,清香更显突出。
猜你喜欢
山西化工(2022年4期)2022-09-23
中学生数理化·高一版(2022年4期)2022-05-09
中学化学(2021年11期)2021-12-09
昆明医科大学学报(2021年8期)2021-08-13
江苏农业科学(2019年8期)2019-08-20
科技创新导报(2018年1期)2018-05-07
中成药(2017年9期)2017-12-19
中成药(2017年8期)2017-11-22
科学与财富(2017年17期)2017-06-16
纺织导报(2017年4期)2017-05-05
