
人工老化处理对不同休眠特性水稻种子生理特性的影响
2018-07-10 03:06,,,,
种子 2018年6期
, , , ,
(1.湖州市农业局农作物技术推广站, 浙江 湖州 313000;2.浙江农林大学 农业与食品科学学院,浙江省农产品品质改良技术研究重点实验室, 杭州 311300;3.浙江省农业科学院 作物与核技术利用研究所,浙江农科种业有限公司, 杭州 310021)
粮食储备是粮食安全的重要环节[1],稻谷储藏过程中米质劣变、发芽率下降一直是困扰生产、储运和经营部分而又难以克服的问题[2],由此造成我国每年粮食储藏损失高达350亿kg。种子的耐贮性影响粮食储备,因此研究水稻种子的耐贮性尤为重要。种子耐贮性的研究方法主要有自然老化法和人工加速老化法。种子老化是种子活力在生理成熟期达到最高后开始出现不可逆的自然衰退[3]。种子老化可引起种子体内贮藏物质下降[4],有毒物质积累[5],细胞结构破坏,生理代谢紊乱,从而严重影响种子的耐贮性和种子活力[6]。董国军等认为,自然老化非常缓慢,难以在较短时间内测定种子活力,在品种改良中进行耐储藏种质筛选时难以应用[7]。而Ellis等认为,在-13~80 ℃对种子进行处理,种子的老化规律是一致的[8]。因此,用人工加速老化方法模拟自然老化已成为研究种子耐贮性的通用方法。
在杂交水稻制种过程中,通过选育具有强休眠性的保持系和不育系可以解决杂交稻制种穗发芽的危害,延长种子的储藏时间。侯雅杰认为,休眠种子能通过长时间储藏而保持活力,适应性较强[9]。但也有学者认为,休眠性强的种子可能会影响种子的萌发,并不一定更耐储藏[10]。种子休眠性和耐藏性之间的关系研究较少,仍不明确。本研究采用高温高湿人工加速老化方法,对2个具有休眠特性差异的水稻品种及其杂交F2代种子进行老化处理,研究老化对不同休眠特性差异水稻种子生理特性的影响,以期揭示水稻种子休眠性与耐藏性的关系,为耐贮性水稻品种选育提供理论参考。
1 材料与方法
1.1 材 料
供试材料为籼稻品种4 K 58(Ⅱ-32 B休眠)、4 K 59(Ⅱ-32 B)、C 178和C 179的种子。4 K 58种子具有休眠性;C 178和C 179分别是4 K 58和4 K 59与不育系(Ⅱ-32 A)杂交获得的F2代种子。
采用人工加速老化方法对供试材料进行处理:100%RH,40 ℃,老化3,6,9 d。老化结束后将种子放在室温下晾干24 h,用纸袋包装,4 ℃保存备用。以未老化种子为对照(ck)。
1.2 方 法
1.2.1种子丙二醛含量测定
分别将不同老化天数处理的种子均匀置于湿润的滤纸上吸胀24 h[11],参照Cao等[12]的方法测定丙二醛(MDA)的含量。称取0.15 g的吸胀种子样品,加入4 mL 0.05 mol/L磷酸缓冲液(pH=7.8)进行研磨,然后在10 000 r/min条件下离心15 min。取1.5 mL上清液,加入2.5 mL 5%(w/v)硫代巴比妥酸-三氯乙酸溶液。混合溶液在100 ℃下煮沸15 min,然后迅速冷却。以1 800 r/min离心10 min后,测定上清液在532 nm和600 nm下的吸光度值。采用155 mM-1cm-1消光系数计算MDA的含量。
1.2.2种子抗氧化物酶活性测定
分别将不同老化天数处理的种子均匀置于湿润的滤纸上吸胀24 h,称取0.15 g的吸胀种子样品,加入4 mL 0.05 mol/L磷酸缓冲液(pH=7.8)进行研磨,然后在10 000 r/min条件下离心15 min,取上清液作为测定酶活性的粗酶液[13]。根据Hu等[13]的方法,通过监测H2O2在240 nm下吸光度值的降低来测定过氧化氢酶(CAT)的活性。反应液包括2.8 mL 25 mmol/L磷酸缓冲液(pH=7.0,含2 mmol/L乙二胺四乙酸二钠(EDTANa2)),100μL 300 mmol/L H2O2和100μL粗酶提取液。参照Nakano等[14]的方法,通过监测反应液在290 nm下吸光度值的降低来检测抗坏血酸过氧化物酶(APX)活性,3 mL混合液中含有2.7 mL 25 mmol/L磷酸缓冲液(pH=7.0,含2 mmol/L EDTANa2)、100μL 300 mmol/L H2O2和100μL粗酶提取液。根据Guan等[15]的方法,通过测定1 min内反应液在470 nm下吸光度值的增加来测定过氧化物酶(POD)活性。反应液含有2.7 mL 25 mmol/L磷酸缓冲液(pH=7.0,含2 mmol/L EDTANa2)、100μL 1.5%愈创木酚、100μL 300 mmol/L H2O2和100μL粗酶提取液。根据Xu等[16]的方法测定超氧化物歧化酶(SOD)的活性。在560 nm下,以抑制NBT光化还原反应50%时的酶量作为一个酶活性单位,计算总的SOD活性。
1.3 统计分析
所得数据用SAS进行统计分析,多重比较采用最小显著差法(LSD),α=0.05,百分率数据在分析前进行反正弦转换(y=arcsin[sqrt (x/100)])。
2 结 果
2.1 不同老化时间处理对种子MDA含量的影响
未经老化处理的种子(ck)中,C 178和C 179的MDA含量均显著高于4 K 58和4 K 59;C 178和C 179之间,4 K 58和4 K 59之间均无显著差异。种子老化3 d后,C 178种子的MDA含量显著高于4 K 58和4 K 59,后两者之间无显著差异。C 178和C 179之间也无显著差异。种子老化6 d后,4 K 59种子的MDA含量显著低于其余3个品种,3个品种之间无显著差异。种子老化9 d后,C 178种子的MDA含量显著高于其余3个品种,C 179种子的MDA含量显著高于4 K 58和4 K 59,4 K 58和4 K 59之间无显著差异。
表1不同水稻品种老化处理后种子MDA含量比较
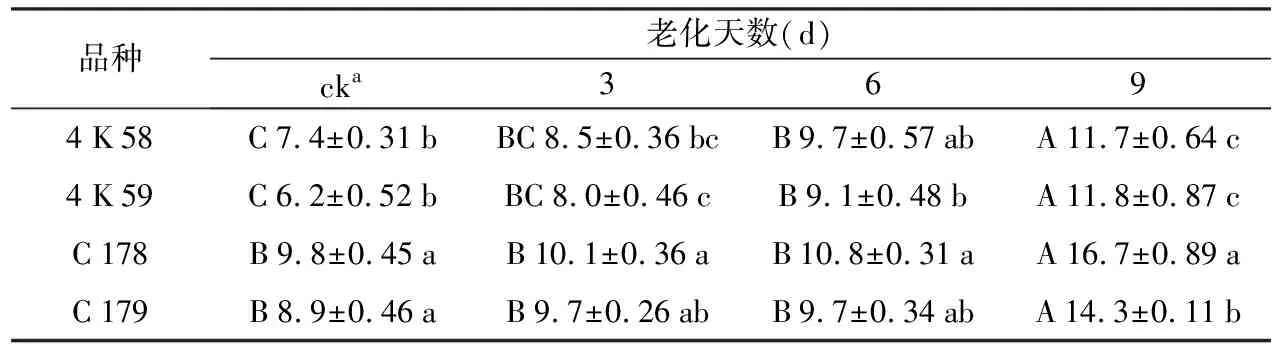
品种老化天数(d)cka3694K58C7.4±0.31bBC8.5±0.36bcB9.7±0.57abA11.7±0.64c4K59C6.2±0.52bBC8.0±0.46cB9.1±0.48bA11.8±0.87cC178B9.8±0.45aB10.1±0.36aB10.8±0.31aA16.7±0.89aC179B8.9±0.46aB9.7±0.26abB9.7±0.34abA14.3±0.11b
注:cka是指未经老化处理的种子。数值后小写字母表示不同水稻品种经相同老化天数处理后的种子MDA含量差异显著性;数值前大写字母表示同一水稻品种经不同老化天数处理后的种子MDA含量差异显著性(LSD,α=0.05)。
随老化处理时间延长,4个品种种子MDA含量均呈逐渐上升趋势。老化9 d后,4个品种种子MDA含量均显著高于老化6 d。老化6 d后,4个品种种子MDA含量与老化3 d无显著差异,但4 K 58和4 K 59的种子MDA含量均显著高于ck。老化3 d后,4个品种中种子MDA含量与ck无显著差异。
2.2 不同老化时间处理对种子CAT活性的影响
2.2.1不同老化时间处理对种子CAT活性的影响
未经老化处理的种子(ck)中,4 K 58的CAT活性与4 K 59无显著差异,二者均显著低于C 178;C 178的CAT活性显著低于C 179。种子老化3 d后,4 K 58种子的CAT活性显著低于4 K 59,4 K 59则显著低于C 178,C 178的CAT活性显著低于C 179。种子老化6 d后,4 K 58种子的CAT活性显著低于4 K 59;4 K 59与C 178之间无显著差异,但二者均显著低于C 179。种子老化9 d后,4个品种种子的CAT活性变与同老化3 d一致。
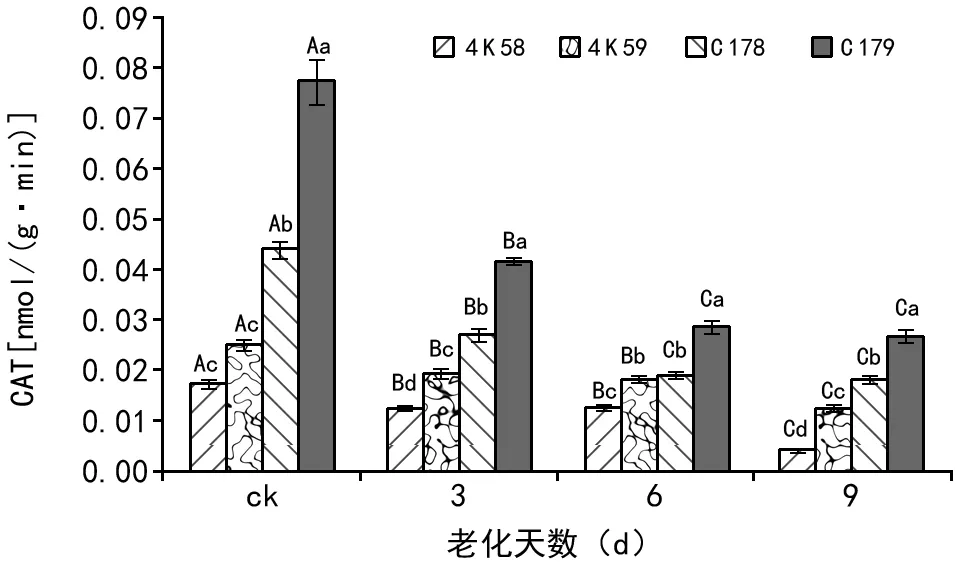
注:ck是指未经老化处理的种子。柱形图上方小写字母表示不同水稻品种经相同老化天数处理后的种子CAT活性差异显著性;大写字母表示同一水稻品种经不同老化天数处理后的种子CAT活性差异显著性(LSD,α=0.05)。图1 不同水稻品种老化处理后种子的CAT活性
随老化时间的延长,4个品种种子的CAT活性均呈下降趋势。老化3 d后,4个品种种子的CAT活性均显著低于ck。老化6 d后,4 K 58和4 K 59种子的CAT活性与老化3 d无显著差异;C 178和C 179的CAT活性均显著低于老化3 d。老化9 d后,4 K 58和4 K 59种子的CAT活性均显著低于老化6 d;C 178和C 179的CAT活性与老化6 d无显著差异。
2.2.2不同老化时间处理对种子APX活性的影响
未老化的4 K 58种子APX活性显著低于4 K 59,4 K 59则显著低于C 178,C 178的APX活性显著低于C 179。种子分别老化3 d和6 d后,4个品种种子的APX活性变化与未老化处理一致。种子老化9 d后,4 K 58种子的APX活性显著低于4 K 59;4 K 59与C 178之间无显著差异,二者均显著低于C 179。

注:ck是指未经老化处理的种子。柱形图上方小写字母表示不同水稻品种经相同老化天数处理后的种子APX活性差异显著性;大写字母表示同一水稻品种经不同老化天数处理后的种子APX活性差异显著性(LSD,α=0.05)。图2 不同水稻品种老化处理后种子的APX活性
随老化时间的延长,4个品种种子的APX活性均呈下降趋势。除C 179之外,未老化处理的其余3个品种种子APX活性均显著高于老化3 d的种子。老化6 d后,4 K 58和4 K 59种子的APX活性与老化3 d无显著差异;C 178和C 179的APX活性均显著低于老化3 d。老化9 d后,除4 K 59之外,其余3个品种种子APX活性均显著低于老化6 d。
2.2.3不同老化时间处理对种子POD活性的影响
未老化处理的种子中,4 K 59的POD活性显著高于其余3个品种;4 K 58显著高于C 178和C 179;后两者之间无显著差异。种子老化3 d后,4 K 59种子的POD活性显著高于其余3个品种,4 K 58显著高于C 179,C 179显著高于C 178。种子老化6 d后,4个品种种子的POD活性变化与老化3 d一致。种子老化9 d后,4个品种种子的POD活性变化与未老化处理(ck)一致。
随老化时间的延长,4个品种种子的POD活性均呈下降趋势。种子老化3 d后,4 K 58和4 K 59的种子POD活性与未老化处理(ck)无显著差异,C 178和C 179种子的POD活性均显著低于ck。种子老化6 d后,4 K 59和C 178的POD活性显著低于老化3 d,4 K 58和C 179与老化3 d无显著差异。种子老化9 d后,除4 K 59的POD活性与老化6 d无显著差异外,其余3个品种均显著低于老化6 d。
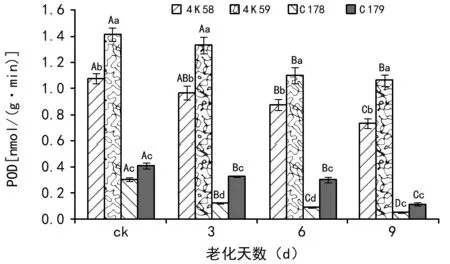
注:ck是指未经老化处理的种子。柱形图上方小写字母表示不同水稻品种经相同老化天数处理后的种子POD活性差异显著性;大写字母表示同一水稻品种经不同老化天数处理后的种子POD活性差异显著性(LSD,α=0.05)。图3 不同水稻品种老化处理后种子的POD活性
2.2.4不同老化时间处理对种子SOD活性的影响
未老化的4 K 58种子SOD活性显著低于4 K 59,4 K 59显著低于C 178和C 179,C 178和C 179之间无显著差异。种子老化3 d后,4个品种种子的SOD活性变化与ck一致。种子老化6 d后,4 K 58的SOD活性显著低于其余3个品种,后三者之间无显著差异。种子老化9 d后,4 K 58的SOD活性显著低于C 178,C 178显著低于4 K 59,4 K 59显著低于C 179。

注:ck是指未经老化处理的种子。柱形图上方小写字母表示不同水稻品种经相同老化天数处理后的种子SOD活性差异显著性;大写字母表示同一水稻品种经不同老化天数处理后的种子SOD活性差异显著性(LSD,α=0.05)。图4 不同水稻品种老化处理后种子的SOD活性
随老化时间的延长,除4 K 59种子的SOD活性无显著变化外,其余3个品种种子的SOD活性均呈下降趋势。种子老化3 d后,3个品种种子的SOD活性与未老化处理无显著差异。种子老化6 d后,C 178的SOD活性显著低于老化3 d,其余2个品种与老化3 d无显著差异。种子老化9 d后,4 K 58和C 178的SOD活性显著低于老化6 d。
3 讨 论
种子老化是作物种子贮藏过程中普遍存在的一种自然现象。在种子老化过程中,其内部会发生一系列的生理生化变化,包括膜完整性的破坏和膜的自动氧化,种子内酶活性变化,有毒物质积累,种子的合成修复能力降低等[17]。本研究采用人工加速老化方法对2个具有休眠特性差异的籼稻品种及其各自与不育系的杂交F2代种子进行老化处理,发现所有种子经老化处理后劣变程度加深,表现为MDA含量显著上升,抗氧化物酶(SOD、POD、CAT和APX)活性均下降等,且不同品种间存在显著差异。
MDA通常作为脂质过氧化指标,其含量在一定程度上可以反映膜脂质过氧化水平[18]。本研究发现,4个水稻品种种子随老化时间的延长,MDA含量均呈现显著上升的趋势,说明老化引起脂质过氧化作用加强,这与小麦[19]、糯玉米[20]和高粱[21]等种子研究结果一致。此外,本研究还发现,在所有处理中,2个自交系种子(4 K 58和4 K 59)的MDA含量均显著低于2个杂交种(C 178和C 179),这可能与杂交水稻种子比常规稻(自交系)种子不耐贮藏有关[22]。杂交稻种子具有的遗传特性使其米粒组织疏松,闭颖较差,保护性能下降,易受外界因素影响,不利于贮藏;同时,杂交稻种子生理代谢旺盛,呼吸强度比常规稻更大,也使得其贮藏稳定性较差[23]。休眠特性差异对老化后种子的MDA含量有一定影响,但不显著,仅在老化9 d后的C 178种子中,发现其MDA含量显著高于C 179。
种子老化的另一个重要表现为种子体内抗氧化物酶类活性的下降。李春雷等对人工老化处理的玉米种子活力指标和生理指标进行了研究,发现种子活力与过氧化物酶、过氧化氢酶、脱氢酶活性的相关系数达到显著或极显著水平,能很好地预测种子活力水平[4]。因此,研究种子老化过程中抗氧化物酶活性的变化对于了解种子活力及劣变程度具有重要参考价值。本研究发现,随着老化时间的延长,4个水稻品种种子的抗氧化物酶活性均呈下降趋势,尤其是CAT、APX和POD 3种酶。这些结果与前人在玉米[4]、桔梗[11]和小麦[24]等种子的研究结果一致。类似的,在所有处理中,2个自交系种子的SOD、CAT和APX活性均显著低于2个杂交种,这进一步验证了杂交稻种子生理代谢比常规稻更旺盛,呼吸强度更大[22-23]。但需注意的是,2个自交系种子的POD活性却显著高于2个杂交种,这可能与水稻品种或抗氧化物酶的类别不同有关。种子的休眠特性差异对抗氧化物酶活性的影响显著:在所有处理中,4 K 58种子的抗氧化物酶(SOD、CAT、APX和POD)活性均显著低于4 K 59;同样,由4 K 58和不育系的杂交F2代种子(C 178),其抗氧化物酶活性也低于C 179。另外,结果还发现,随老化时间的延长,4 K 58和C 178种子的SOD、APX和POD活性下降程度均分别较4 K 59和C 179更为剧烈。由于种子的抗氧化物酶活性与种子活力水平呈显著正相关[4,25],表明具有休眠特性的水稻种子也不耐贮藏。
综上,经人工加速老化处理后,4个水稻品种种子内MDA含量随老化时间延长而显著增加,抗氧化物酶活性逐渐下降;表明老化处理显著抑制了种子体内抗氧化系统的活性,加深了膜脂过氧化程度,引发或加剧了种子的劣变。杂交F2代种子的MDA含量和抗氧化物酶活性均显著高于自交系种子,证明杂交稻种子比常规稻种子更不耐贮藏。此外,由于4 K 58和C 178种子的抗氧化物酶活性均分别低于4 K 59和C 179,推测具有休眠性的水稻种子更不耐贮藏。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年7期)2023-01-05
现代仪器与医疗(2021年4期)2021-11-05
中国人兽共患病学报(2020年11期)2020-12-08
保健医苑(2020年11期)2020-12-04
数学大王·中高年级(2020年5期)2020-05-25
小型微型计算机系统(2019年4期)2019-05-05
电子制作(2019年24期)2019-02-23
电子制作(2018年10期)2018-08-04
小天使·六年级语数英综合(2017年8期)2017-08-04
Coco薇(2016年2期)2016-03-22
