
草鱼MyoD基因SNP和Indel标记的筛选及其与生长性状的关联分析
2018-07-06 08:55何吉祥樊佳佳吴本丽宋光同
江苏农业学报 2018年3期
陈 静, 何吉祥, 樊佳佳, 黄 龙, 吴本丽, 宋光同, 汪 翔, 武 松
(1.安徽省农业科学院水产研究所,安徽 合肥 230031; 2.农业部热带亚热带水产资源利用与养殖重点实验室,广东 广州 510380)
生肌决定因子基因MyoD是生肌调节因子基因MRFs家族(MyoD、MyoG、Myf5和MRF4)的主要成员之一,生肌调节因子具有一个保守的碱性螺旋-环-螺旋结构(bHLH)[1-2]。MyoD在成肌细胞分化为肌纤维的过程中起关键的正向调控作用,控制着很多关键基因的表达[3-5]。因为时空表达模式不同,4个MRFs家族成员在动物肌肉发育过程中所发挥的作用不同[6],其中MyoD基因是脊椎动物胚胎期肌肉发育的主要调控基因,主要作用是促进骨骼肌的形成和分化,缺少MyoD基因可导致成肌细胞无法正常增殖和分化[7-8]。MyoD与肌肉基因启动子的E-BOX区域结合后,能激活肌肉特异性转录,进而促进肌前体细胞的增殖和分化,另外MyoD基因还可使成纤维细胞及脂肪细胞等转化为成肌细胞,最终分化为成熟肌纤维[9]。
单核苷酸多态性(Single nucleotide polymorphism, SNP)和插入/缺失(Insertion/deletion, Indel)标记作为新型分子标记,广泛分布于整个基因组,是动植物中普遍存在的一种遗传标记[10-12]。草鱼(Ctenopharyngodonidella)是中国淡水养殖鱼类中产量最大的养殖品种,至今仍没有通过国家良种审定委员会鉴定的人工选育的良种。基于MyoD基因对动物肌肉发育和生长的重要作用,本研究以MyoD基因作为草鱼生长性状的候选基因,通过直接测序法检测SNP和Indel标记,并与相关生长性状进行关联分析,以期获得与草鱼生长性状相关联的SNP和Indel标记作为草鱼分子标记辅助选育的候选标记,缩短育种周期,提高选择效率。
1 材料与方法
1.1 材料
收集尽量多来源的草鱼共20尾作为筛选突变位点的样本,取鱼的尾鳍,-20 ℃保存备用。生长性状关联性分析的草鱼样品采自长丰县盛源养殖合作社,随机选择同期繁殖、同塘养殖的3龄草鱼共297尾,测量体质量、全长、体长、体宽、体高、眼间距、尾柄长和尾柄高8项生长指标,同时剪取尾鳍-20 ℃保存。
1.2 基因组DNA的提取与检测
使用QIAamp DNA Mini Kit (北京Qiagen公司产品)提取基因组DNA,用1.5%琼脂糖凝胶电泳检测核酸的质量,-20 ℃保存待用。
1.3 MyoD基因的PCR扩增及突变位点基因分型
根据本实验室克隆得到的草鱼MyoD基因序列(GenBank登录号:MG544985)与草鱼全基因组测序序列(http://www.ncgr.ac.cn/grasscarp/)[13]进行比较,在5′侧翼区、外显子和内含子上筛选多态位点。根据突变位点采用Primer Premier 5.0 软件进行引物设计。
以不同来源的20尾草鱼为模板进行PCR扩增。PCR反应体系包括10.0 μl 2×TaqPCR master mix、上下游引物(20 μmol/L)各0.5 μl、模板DNA 20 ng,加ddH2O至20.0 μl。PCR反应程序:94 ℃ 10 min;94 ℃ 10 s,57 ℃ 15 s,72 ℃ 30 s,共35个循环,72 ℃延伸10 min,保存于4 ℃。PCR产物用1.2%琼脂糖凝胶电泳检测。PCR产物纯化后送至上海捷瑞生物工程有限公司测序。测序结果用软件BioEdit进行比对和筛选,获得多态性高、分型稳定的突变位点。采用直接测序法对297尾草鱼MyoD基因的突变位点进行分型。
1.4 数据统计分析
采用PopGen 32软件计算有效等位基因数(Ne)、观测杂合度(Ho)和期望杂合度(He),利用Picalc程序包计算各位点的多态信息含量(PIC)。采用SPSS 20软件一般线性模型(General linear model,GLM)分析突变位点各基因型与生长性状的关联程度,因变量为草鱼8项生长指标,自变量为筛选到的突变位点的不同基因型。其生物统计模型为:Yij=μ+Bi+eij,其中Yij表示某性状第i个标记在第j个个体上的观测值,μ表示试验观测的所有个体平均值,Bi表示第i个标记的效应值,eij表示对应个体观测值的随机残差效应。
2 结果与分析
2.1 MyoD基因突变位点筛选
通过序列比对,在MyoD基因5′侧翼区、外显子和内含子上共筛选得到6个突变位点,分别命名为M1(940后T碱基的插入)、M2(A1601G)、M3(1630后重复单元AATAGCCT的缺失)、M4(G1669A)、M5(A1681T)、M6(T2322G)。其中M1位于5′侧翼区,M2、M3、M4和M5位于内含子1,M6位于外显子3。M6位点(T2322G)的突变没有导致氨基酸的变化,属于同义改变。
共设计3对引物在20个草鱼样本中对这6个突变位点进行验证(引物序列和参数见表1),3对引物PCR 扩增分别获得409 bp、354 bp和245 bp片段。PCR产物经测序和比对后获得多态性高、分型稳定的4个位点,即M1、M3、M4和M5。

表1 草鱼MyoD基因扩增多态位点检测引物
2.2 MyoD基因突变位点在草鱼群体中的遗传结构分析
采用直接测序法对297尾草鱼MyoD基因的4个多态位点进行基因分型,统计4个突变位点的遗传参数。结果(表2)表明,有效等位基因数(Ne)为 1.422 0~1.978 6,观测杂合度(Ho)为 0.268 5~0.466 4, 期望杂合度(He)为 0.297 2~0.495 4,多态信息含量(PIC)为 0.252 7~0.372 3,He和PIC均大于0.25小于0.50,说明4个突变位点具有中度多态性[14]。经χ2检验,Hardy-Weinberg平衡常数分别为0.690 3、0.175 6、0.093 2、0.311 8,表明试验群体在该4个突变位点均处于哈温平衡状态(P>0.05)。

表2 草鱼MyoD基因SNP和Indel遗传多样性参数
2.3 MyoD基因突变位点与草鱼生长性状的关联分析
采用一般线性模型分析草鱼MyoD基因4个多态位点不同基因型与生长性状的相关性(表3)。M1位点不同基因型个体间的体长、全长、尾柄长和尾柄高存在显著差异(P<0.05),纯合缺失基因型(DD)显著大于纯合插入基因型(TT),为优势基因型;M3和M4位点不同基因型个体间的生长性状差异均不显著(P>0.05);M5位点不同基因型个体间的体质量、体宽、体高和眼间距存在显著差异(P<0.05),优势基因型为TT。
3 讨 论
本研究中,在草鱼MyoD基因中共发现4个多态性高、分型稳定的突变位点,即M1、M3、M4和M5,其中M1位于5′侧翼区,M3、M4和M5位于内含子1。位于5′侧翼区的M1位点虽然不编码氨基酸,但由于5′UTR在基因表达调控中有重要作用,因而其序列突变可能会对基因表达调控产生变化进而影响机体的多种性状[15-16],在本研究中就发现该突变与草鱼全长、体长等生长性状有显著相关。有研究结果表明,基因组中的突变位点绝大多数位于内含子区域,而外显子相对较保守[8,17]。本研究在草鱼MyoD基因中发现的4个突变位点中就有3个位于内含子,可能是因为内含子不参与氨基酸编码,与外显子相比其受到的选择压力较小,变异更容易积累[18-19]。
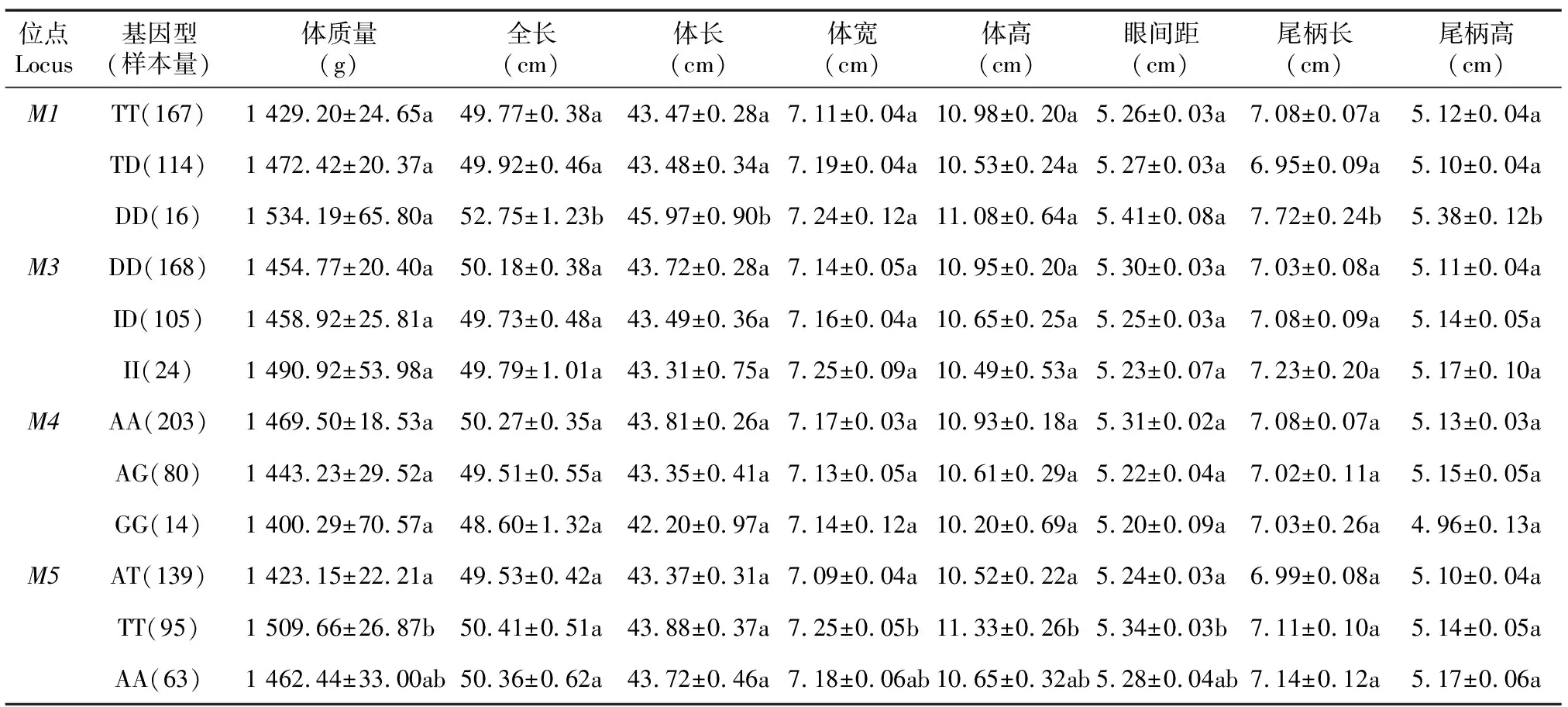
表3 草鱼MyoD基因SNP和Indel位点基因型与生长性状的相关性
相同位点同列数值后不同小写字母表示差异显著(P<0.05)。
MyoD基因是生肌调节因子基因MRFs家族的主要成员之一,是脊椎动物胚胎期肌肉发育的主导调控基因之一,对骨骼肌的形成和分化起主要作用,因此,MyoD基因的核苷酸变异可能会影响一些与肌肉肉质和生长性能相关的生产性状[20]。MyoD基因多态性与生产性能的关联研究在畜牧领域中已经有较多报道。Han等[21]研究发现大白猪MyoD基因的SNP位点g.257A>C与肌肉pH值呈显著关联性。田璐等[22]在肉牛上发现MyoD基因内含子2中不同基因型对肉牛的宰前活质量、胴体质量、净肉质量、高档肉质量、眼肌面积等性状影响极显著或显著。褚敏等[20]在大通耗牛MyoD1基因3′UTR的1 976 bp处发现1个突变位点,对体质量、胸围、体斜长方面影响显著。李万贵等[23]在樱桃谷鸭MyoD基因外显子4发现2个SNP位点,突变基因型TT鸭群体显著小于野生基因型AA群体。Chen等[24]研究发现MyoD1a和MyoD1b对虹鳟肉质有重要影响。陈松波等[25]研究发现牙鲆MyoD基因内含子1上的SNPs对牙鲆的生长性状影响显著。在本研究中,MyoD基因多态性位点与草鱼生长性状的相关分析结果表明,5′侧翼区M1位点不同基因型个体间的体长、全长、尾柄长和尾柄高存在显著差异(P<0.05),纯合缺失基因型(DD)显著大于纯合插入基因型(TT);内含子1中M5位点不同基因型个体间的体质量、体宽、体高和眼间距存在显著差异(P<0.05),与这些经济性状连锁的优势基因型为TT。因此可以将草鱼MyoD基因作为分子辅助草鱼选育的候选基因,位点M1(940后T碱基的插入)和M5(A1681T)作为草鱼分子标记辅助育种的候选分子标记。
参考文献:
[1] BUCKINGHAM M. Making muscle in mammals[J]. Trends Genet, 1992, 8(4):144-149.
[2] 蒋运良,李 宁,吴常信.肌肉生成的分子生物学研究进展(综述)[J]. 农业生物技术学报, 1999, 7(2):201-204.
[3] FRANCETIC T, LI Q. Skeletalmyogenesis andMyf5 activation[J]. Transcription, 2011, 2(3): 109-114.
[4] RESCAN P Y. Regulation and functions of myogenic regulatory factors in lower vertebrates[J]. Comp Biochem Physiol B Biochem Mol Biol, 2001, 130(1):1-12.
[5] NAIDU P S, LUDOLPH D C, TO R Q, et al. Myogenin and MEF2 function synergistically to activate the MRF4 promoter during myogenesis[J]. Molecular and Cellular Biology, 1995, 15(5): 2707-2718.
[6] CARVAJAL J J, RIGBY P W. Regulation of gene expression in vertebrate skeletal muscle[J]. Experimental Cell Research, 2010, 316(18):3014-3018.
[7] WYZYKOWSKI J C, WINATA T I, MITIN N, et al. Identification of novelMyoDgene targets in proliferating myogenic stem cells[J]. Molecular & Cellular Biology, 2002, 22(17): 6199-6208.
[8] 于凌云,白俊杰,叶 星,等. 大口黑鲈MyoD基因结构和单核苷酸多态性位点的筛选[J]. 水产学报, 2009, 33(1):1-8.
[9] 李永平,梁炳生.MyoD肌形成作用机制研究进展[J]. 国际骨科学杂志, 2007, 28(1):37-40.
[10] 赵广珍,贾 青,张翠翠,等. 猪MyoD1基因多态性与肉质性状的相关分析[J].河北农业大学学报, 2012, 35(4):90-94.
[11] 程金花,赵文明,包文斌,等. 皖西白鹅Pit-1基因插入/缺失多态性及其对早期体重的影响[J]. 农业生物技术学报, 2008, 16(3):426-429.
[12] 邱峰芳,聂庆华,金卫根,等. 鸡PIT-1基因57bp插入/缺失多态与生长和屠体性状的相关研究[J]. 江西农业大学学报, 2006, 28(2):284-288.
[13] WANG Y P, LU Y, ZHANG Y, et al. The draft genome of the grass carp (Ctenopharyngodonidellus) provides insights into its evolution and vegetarian adaptation[J]. Nature Genetics, 2015, 47(6):1-8.
[14] BOSTEIN D, WLLITE R L, SKOLNICK M, et al . Construction of a genetic linkage map in man using restriction fragment length polymorphism [J]. The American Journal of Human Genetics, 1980, 32:314-331.
[15] WANG L, LI K, XU Q, et al. Potential synergy between SNP and CpG-A or IL-1 beta in regulating transcriptional activity of IL-20 promoter[J]. The Journal of Investigative Dermatology, 2014, 134(2): 389-395.
[16] YANG Z H, ZHOU C L, ZHU H, et al. A functional SNP in the MDM2 promoter mediates E2F1 affinity to modulate cyclin D1 expression in tumor cell proliferation[J]. Asian Pacific Journal of Cancer Prevention, 2014,15(8):3817-3823.
[17] NIE Q, LEI M, QU Y, et al. Identification and characterization of single nucleotide polymorphisms in 12 chicken growth-correlated genes by denaturing high performance liquid chromatography [J]. Genetics Selection Evolution, 2005, 37 (3): 339-360.
[18] 张 猛,陈 勇,沈玉帮,等. 草鱼MSTN-1基因多态性及与早期生长性状和肌肉成分关联分析[J]. 水产学报, 2016, 40(4):618-625.
[19] ZHAO Z M, FU Y X, HEWETT E D, et al. Investigating single nucleotide polymorphism (SNP) density in the human genome and its implications for molecular evolution [J]. Gene, 2003, 312:207-213.
[20] 褚 敏,阎 萍,粱春年,等. 大通牦牛MyoD1基因3′UTR SNPs多态性及其与生长性状相关性的研究[J]. 中国畜牧兽医, 2012, 39(2):111-113.
[21] HAN X, JIANG T, YANG H, et al. Investigation of four porcine candidate genes (H-FABP,MYOD1, UCP3 and MASTR) for meat quality traits in large white pigs[J]. Mol Biol Rep, 2012, 39(6): 6599-6605.
[22] 田 璐,许尚忠,岳文斌,等.MyoD基因对肉牛胴体性状影响的分析[J]. 遗传, 2007, 29(3): 313-318.
[23] 李万贵,张依裕,王单单,等. 樱桃谷鸭MyoD1基因外显子4多态与体尺的关联效应研究[J]. 基因组学与应用生物学, 2015, 34(5):950-954.
[24] CHEN W X, MA Y, LIU K H. Association ofMYOD1aandMYOD1bgene polymorphisms and meat quality traits in rainbow trout[J]. Genetics and Molecular Research, 2015, 14(3): 9034-9044.
[25] 陈松波,龚 丽,范兆廷,等. 牙鲆MyoD基因单核苷酸多态性位点的筛选[J]. 水生生物学报, 2013, 37(5):863-868.
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年3期)2022-04-26
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
新世纪智能(高一语文)(2020年12期)2020-06-01
当代水产(2019年9期)2019-10-08
生物工程学报(2019年6期)2019-07-10
生物学通报(2019年1期)2019-02-15
广东第二课堂·小学(2018年9期)2018-10-24
中国棉花(2018年6期)2018-07-06
科技视界(2016年27期)2017-03-14
