
高通量测序技术在瘤胃产甲烷菌群研究中的应用
2018-06-25 07:56:28赵圣国王加启
家畜生态学报 2018年5期
周 旭,闵 力,赵圣国,郑 楠,2,王加启
(1.中国农业科学院 北京畜牧兽医研究所,动物营养学国家重点实验室,北京 100193;2.中国农业科学院 北京畜牧兽医研究所,农业部奶产品质量安全风险评估实验室(北京),北京 100193;3.新疆农业大学 动物科学学院,新疆 乌鲁木齐 830052)
甲烷是全球温室气体排放量仅次于二氧化碳的温室气体,但是其在地球的升温潜能值(Global warming potential,GWP)却远高于CO2,约为CO2的25倍[1]。有研究表明,农牧业排放甲烷占总甲烷排放量的40%,其中反刍动物释放的甲烷比例最大,占据农牧业甲烷排放量的25%,是甲烷排放的最主要来源[2]。另有研究表明,反刍动物瘤胃排放甲烷不仅会造成2%~12%的消化能损失,降低饲料利用率,而且导致动物生产性能降低,养殖成本增加[3]。瘤胃甲烷减排已经成为缓解环境温室效应和提高能量利用效率的重要途径。因此,对反刍动物瘤胃产甲烷菌群的研究显得尤为重要。
产甲烷菌(methanogen)在生物降解过程的最后一个环节发挥着关键作用,影响甲烷的合成[4]。近年来,高通量测序技术已经被广泛应用于瘤胃产甲烷菌群的研究,它不仅能够分析瘤胃产甲烷菌群落结构和功能,还能揭示瘤胃产甲烷菌群与环境之间的关系。因此,高通量测序技术的应用对研究反刍动物瘤胃产甲烷菌群的多样性与功能,寻找甲烷调控的靶标菌群具有重要的指导意义。
1 瘤胃产甲烷菌的种类
产甲烷菌是一类独特的微生物群体[5]。在产生甲烷的过程中,产甲烷菌含有一种参与甲烷代谢途径的特殊低电位电子载体——F420辅酶因子,它能使瘤胃中约2.8%~4.0%瘤胃产甲烷菌氧化态时吸收420 nm左右的紫外光,激发出约470 nm的蓝绿色荧光[6]。利用产甲烷菌具有的自发荧光的光谱学特征,可在紫外荧光显微镜下鉴定产甲烷菌的菌落或细胞[7]。此外,与其他细菌相比产甲烷菌在细胞膜的组成上也有所不同,产甲烷菌的细胞壁没有胞壁酸,细胞膜的脂质通过乙醚、类异戊二烯与甘油或者碳水化合物相结合[7]。通过16S rRNA分析其序列发现,反刍动物瘤胃中存在的古菌基本为严格厌氧的产甲烷菌,能将无机或有机化合物厌氧发酵转变成CH4和CO2[8]。一般而言,瘤胃产甲烷菌主要有5种,即:反刍兽甲烷短杆菌(Methanobrevibacterruminantium)、甲酸甲烷杆菌(Methanobacteriumformicicum)、巴氏甲烷八叠球菌(Methanosarcinabareri)、马氏甲烷八叠球菌(Methanosarcinamazei)和活动甲烷微菌(Methanomicrobiummobile)[9]。近年来,还发现了未分类的热原体古菌C簇甲烷菌(Rumen Cluster C/RCC)[10]。
2 高通量测序技术的特点
高通量测序技术能够一次测定数百万条DNA分子序列,有利于对一个物种的转录组和基因组进行细致全面的研究分析[11]。高通量测序技术主要包括Roche公司的454焦磷酸测序技术、ABI公司的SOLID技术、Life Technologies的Solid技术、Illlumina公司的HiSeq和MiSeq技术Pacific Biosciences公司的SMRT技术、Helicos Biosciences公司的单分子测序技术和Oxford Nanopore Technologies公司的纳米孔单分子技术[12-13]。表1对已实现商品化的三代测序平台各自的特点进行了比较。
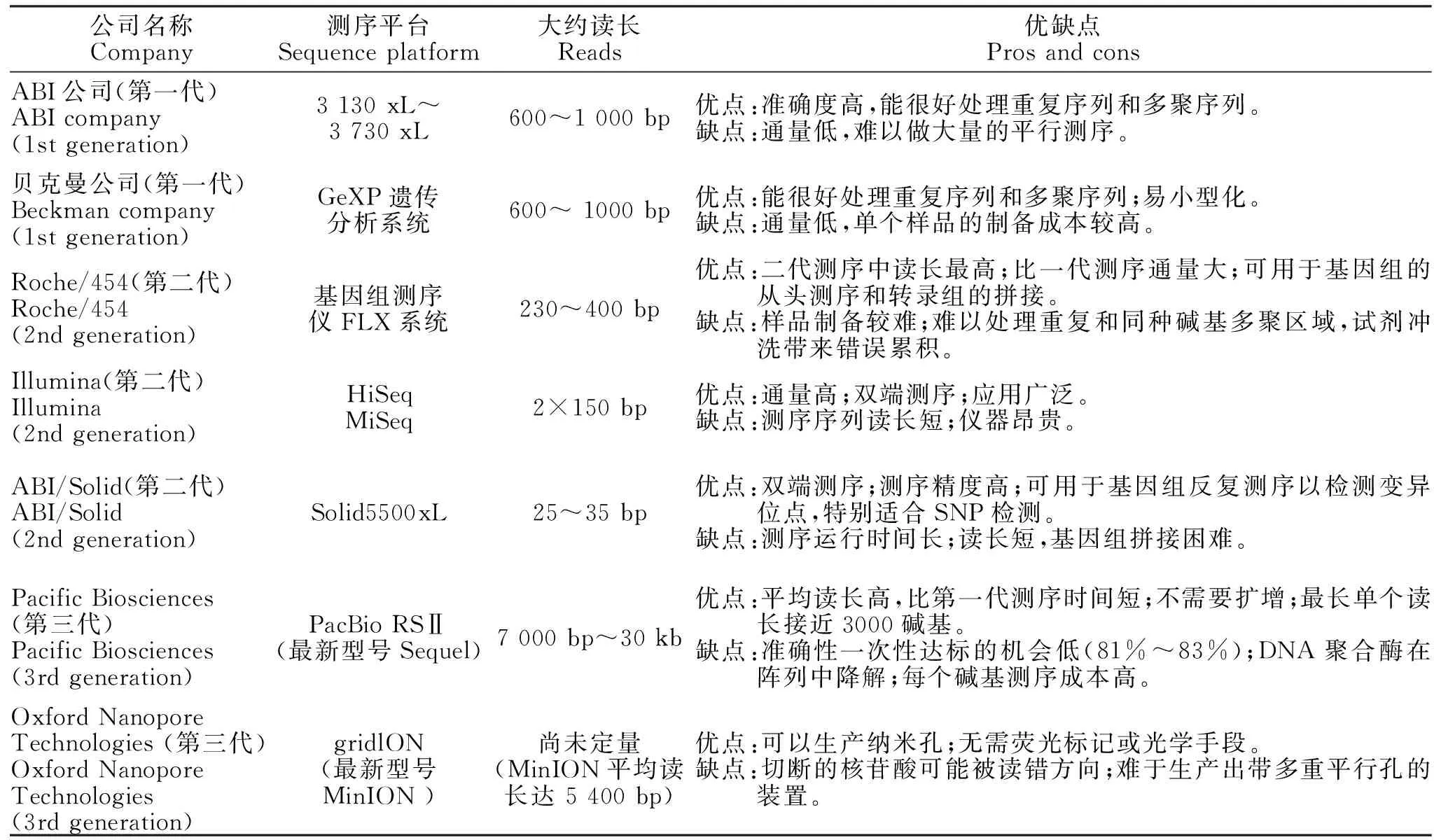
表1 已实现商品化的三代测序平台特点比较Table 1 Comparison of the characteristics on commercialized sequencing platform
目前,以Illumina测序平台为代表的第二代测序平台在通量、准确度上都有较大的提高,同时测序成本也大幅度降低,成为商用测序的主流平台。但在序列读长方面,第二代测序技术比第一代测序技术要短很多。第三代测序技术通过现代光学、高分子、纳米技术等手段来区分碱基信号间的差异,达到直接读取序列信息的目的,在DNA序列片段读长上优于第二代,但在准确度上比第二代稍差,价格也偏贵。未来随着技术的改善,第三代测序将更为稳定和成熟。
3 高通量测序在瘤胃产甲烷菌群基因组研究中的应用
基因组数据对一个物种基因的组成结构和不同物种间基因进化关系的研究起着决定性的作用。国内外的学者对瘤胃产甲烷菌群基因组进行了大量的研究,目前已完成MethanobrevibacterruminantiumM1、Methanobrevibactersp. AbM4、MethanobacteriumformiciumBRM9、Methanosarcinasp. CM1、MethanobrevibactermilleraeSM9、Methanosphareasp. ISO3-F5等大部分瘤胃甲烷菌基因组测序项目[14-15],并且研究者们还新发现了反刍动物瘤胃第二大产甲烷菌群甲烷菌第七目Methanomassiliicoccales[16-17]。通过对产甲烷菌基因组测序和注释分析等组学的研究,可以发现其基因组中参与产甲烷途径酶的编码基因,推测产甲烷菌基本生理代谢途径,分析出产甲烷菌的氧代谢、氢利用、一氧化碳代谢途径和固氮的过程,全面了解未知产甲烷菌个体和群落的基本代谢信息,从而抑制甲烷菌的产生以及甲烷的排放。
4 高通量测序在瘤胃产甲烷菌群多样性研究中的应用
以往对瘤胃产甲烷菌群多样性的检测是通过mcrA或SSU rRNA数据库设计引物,qPCR扩增检测特定类菌群微生物数量,化学标记物检测产甲烷菌代谢附属物等方法来分析瘤胃产甲烷菌的代谢情况[18],但由于引物特异性低或代谢附属物容易受到瘤胃中其他条件影响等因素,导致检测精度低、获得的信息量少[19]。而高通量测序技术能更高效用于检测瘤胃产甲烷菌多样性的研究,对瘤胃复杂微生物环境中产甲烷菌的生物学信息进行更加全面的检测。能够更加准确、深入地分析瘤胃产甲烷菌群落的结构组成、基因功能、代谢途径。
表2比较了影响瘤胃产甲烷菌多样性的因素。由表2可知,动物的品种、饲粮配方等因素对瘤胃产甲烷菌的多样性产生很大差异,存在这种差异的原因首先可能在于测序使用的引物不同[20-21]。Tajima等[22]用0025e/1492和D30/D33两对引物对奶牛瘤胃产甲烷菌种类进行研究,结果表明由于引物扩增效率的不同,使用引物0025e/1492所得到的产甲烷菌种类多于D30/D33。我们认为,由于在不同模板中引物结合部位、周围结构、G+C含量等的不同,都会使部分模板优先扩增,而不同引物优先扩增的种类差异,就会引起菌群多样性差异。因此,引物的差异、PCR扩增的偏差、扩增错误、测序错误等都可能降低菌群多样性估计的准确性。

表2 影响瘤胃产甲烷菌多样性的因素[19]Table 2 The effect of factors on rumen methanogens diversity
饲粮的差异是引起反刍动物瘤胃菌群数量变化和改变优势菌类型的决定性因素。Mccabe等[23]利用高通量测序技术研究了限制饲喂的牛体内微生物的组成和变化,发现与自由采食组奶牛相比,琥珀酸弧菌属在限制饲喂组奶牛瘤胃食糜液相和固相的丰度显著降低,而产甲烷古菌在限制饲喂组奶牛瘤胃食糜中的丰度却非常高,进而发现琥珀酸弧菌属与产甲烷古菌之间存在一种争夺底物的竞争关系。因此,我们推测不同饲粮可能通过影响瘤胃pH值和瘤胃发酵类型等因素,对瘤胃产甲烷菌种类多样性和数量产生不同影响。
宿主动物和瘤胃微生物之间的选择与进化历程的差异也可能引起瘤胃菌群多样性的变化。Wallace等[24]为对比高产甲烷牛与低产甲烷牛瘤胃中产甲烷菌(和酶)的含量,采用Illumina Hiseq测序,将OUT代表序列与RIM-DB数据库进行BLAST比对,获取每个OUT所对应的分类学信息,通过计算相对丰度,得到细菌和古菌的比例,发现高产和低产甲烷牛在细菌、原生动物含量等方面大体相同,只有古菌含量方面存在显著性差异(P<0.01),说明极高产甲烷肉牛的古菌含量显著高于极低产甲烷的肉牛的古菌含量。同时Wallace等[24]将Hiseq 序列与KEGG基因数据库比对,通过功能丰度证明了高产甲烷牛的甲烷合成酶丰度高于低产甲烷牛。由此可知,反刍动物瘤胃菌群多样性是宿主动物与瘤胃微生物强烈选择与协同进化的结果。
5 高通量测序在瘤胃产甲烷菌群功能代谢研究中的应用
产甲烷菌在瘤胃中的主要代谢方式是利用H2和CO2生成CH4。有研究发现,Methanobrevibacter和Methanosphaera主要利用H2,同时Methanosphaera也能够利用甲基营养途径产甲烷[25]。该途径包含两种代谢模式,第一种为严格的甲基营养型模式,一部分甲基化合物被氧化产生CO2以及还原当量,用于还原甲基化合物中的甲基基团产甲烷[26-27]。这也是大多数甲基营养型产甲烷菌的代谢途径,它们仅以甲基化合物为底物产甲烷,第二种为H2依赖型代谢模式,以利用甲醇作为生成CH4的底物,以H2作为电子供体,还原甲基化合物中的甲基基团产甲烷[28],瘤胃未知甲烷菌群C簇[29-30]也是通过该途径产甲烷。具体反应过程见图1。

图1 产甲烷菌的代谢途径

Fig.1 Metabolic pathways of methanogenesis
随着基因改造、基因组测序、转录组测序和生物信息学等新技术的发展,国内外学者对产甲烷菌的功能代谢和调控过程的认识已越来越深入全面。Rubino等[31]对14头奶牛的瘤胃微生物宏基因组进行Illumina GAIIx平台测序,发现甲烷短杆菌属中的基因要比梭菌属中的基因更丰富,在涉及某些特定代谢通路方面,普氏菌属比甲烷短杆菌属拥有更加多样的功能异构体,如丙酮酸氧化、羟基丙酸—羟基丁酸循环;而梭菌属在半胱氨酸生物合成、甲醛同化/丝氨酸代谢、PRPP生物合成方面要比甲烷短杆菌属拥有更加多样的功能异构体。表明普氏菌属会优先降解半纤维素,而当木聚糖化合物生成后,纤维表面集群逐渐被梭菌所取代。Kamke等[32]以23只羊作为研究对象,通过Illumina HiSeq测序平台对低甲烷产量羊瘤胃微生物特征进行研究,KEGG注释发现,与高甲烷产量样品相比,调控氨基酸生物合成、磷酸转移酶系统、半乳糖代谢以及短链脂肪酸代谢的基因在低甲烷产量羊瘤胃微生物中相对丰度更高,暗示这些基因可能与微生物产甲烷量有关,研究证明动物瘤胃容积越小、转换率高、Sharpeaspp.和Megasphaeraspp.两种微生物富集是最终导致低甲烷产量形成的原因。Hoedt等[29]对澳大利亚袋鼠前胃样品微生物多样性进行分析,根据结果进行产甲烷古菌纯化与分离,然后对分离得到的微生物(WGK6)进行鉴定和功能分析,发现WGK6与Methanosphaerastadtmanae(DSMZ 3091T)具有较近的系统发育关系,WGK6能够利用乙醇产生甲烷,同时可生成乙酸盐,推测该菌在H2或乙醇存在时参与甲醇还原过程。因此,高通量测序技术不但可以清楚地了解目的产甲烷菌的代谢过程和系统发育的位置,而且利用高通量测序技术与蛋白质组、基因组、代谢组等其他分子生物学研究手段的结合,能够更加深入的了解复杂瘤胃微生物产甲烷菌群功能基因表达与调控的方式,为鉴定甲烷代谢调控靶点提供更多的信息,进而了解甲烷的产生机制和抑制甲烷的产生。
6 小 结
瘤胃产甲烷菌群落是反刍动物特有的微生物群落,需要不断探索和研究。近年来,在瘤胃产甲烷菌群落的研究中高通量测序技术已经被广泛应用,可以检测未知序列的特征,为部分不可培养的产甲烷菌提供了很好的平台,具有寻找新基因的作用。但在实际的操作中,一方面应注意这些技术本身存在的不足和研究过程中不稳定因素对试验结果的影响,另一方面要注意瘤胃中菌群样品本身的特殊性。因此,如何应用高通量测序技术,从复杂环境中定向检测生长条件苛刻、但是具有特殊生态学功能的产甲烷古菌,了解反刍动物瘤胃中产甲烷菌的群落结构和生理状态,将是一个必须面对和解决的关键问题。
参考文献:
[1] 方晓瑜,李家宝,芮俊鹏,等. 产甲烷生化代谢途径研究进展[J]. 应用与环境生物学报, 2015,21(1):1-14.
[2] KUMAR S, CHOUDHURY P K, CARRO M D, et al. New aspects and strategies for methane mitigation from ruminants[J]. Applied Microbiology and Biotechnology, 2014,98(1):31-44.
[3] ST-PIERRE B, WRIGHT A D. Diversity of gut methanogens in herbivorous animals[J]. Animal,2013,7(s1):49.
[4] DEMIREL B, SCHERER P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: a review[J]. Reviews in Environmental Science and Bio/Technology, 2008,7(2):173-190.
[5] YANAGITA K, KAMAGATA Y, KAWAHARASAKI M, et al. Phylogenetic analysis of methanogens in sheep rumen ecosystem and detection of Methanomicrobium mobile by fluorescence in situ hybridization[J]. Journal of the Agricultural Chemical Society of Japan, 2000, 64(8):1 737-1 742.
[6] CHEESEMAN P, TOMS-WOOD A, WOLFE R S. Isolation and properties of a fluorescent compound, factor, 420, from Methanobacterium strain M.o.H[J]. Journal of Bacteriology, 1972, 112(1):527-531.
[7] 孙凯佳,朱建营,梅洋, 等. 降低反刍动物胃肠道甲烷排放的措施[J]. 动物营养学报, 2015,27(10):2 994-3 005.
[8] 承磊,郑珍珍,王聪, 等. 产甲烷古菌研究进展[J]. 微生物学通报, 2016, 43(5):1 143-1 164.
[9] 高民,胡红莲,杜瑞平. 反刍动物甲烷排放及减排策略[J]. 中国畜牧杂志,2013,49(18):35-41.
[10] 金巍,成艳芬,毛胜勇,等. 厌氧真菌培养液中特异RCC甲烷菌的发现[C]//中国畜牧兽医学会动物营养学分会中国饲料营养学术研讨会,2014.
[11] 陈蕾,小理,周一鸣,等. 高通量测序技术在肠道菌群研究中的应用[J]. 食品工业, 2016(3):269-273.
[12] GLENN T C. Field guide to next-generation DNA sequencers[J]. Molecular Ecology Resources, 2011,11(5):759-769.
[13] 沈圣,屈彦纯,张军. 下一代测序技术在表观遗传学研究中的重要应用及进展[J]. 遗传, 2014,36(3):256-275.
[14] 吴鹏,陈忠法,王佳堃. 宏基因组学揭示瘤胃微生物多样性及功能[J].动物营养学报,2017,29( 5):1 506-1 514
[15] MORGAVI D P, KELLY W J, JANSSEN P H, et al. Rumen microbial (meta)genomics and its application to ruminant production.[J]. Animal, 2013,7(s1):184-201.
[16] JIN W, CHENG Y, ZHU W. The community structure of Methanomassiliicoccales in the rumen of Chinese goats and its response to a high-grain diet:[J]. Journal of Animal Science & Biotechnology, 2017,8(1):47.
[17] SEEDORF H, KITTELMANN S, HENDERSON G, et al. RIM-DB: a taxonomic framework for community structure analysis of methanogenic archaea from the rumen and other intestinal environments[J]. Peerj, 2014,2(11):e494.
[18] THORPE A. Enteric fermentation and ruminant eructation: the role (and control) of methane in the climate change debate[J]. Climatic Change, 2009, 93(3/4):407-431.
[19] 裴彩霞, 毛胜勇, 朱伟云. 山羊瘤胃产甲烷古菌多样性及与其他动物瘤胃的比较[J]. 畜牧兽医学报, 2012, 43(6):909-914.
[20] TYMENSEN L D, MCALLISTER T A. Community structure analysis of methanogens associated with rumen protozoa reveals bias in universal archaeal primers[J]. Applied and Environmental Microbiology, 2012,78(78):4 051-4 056.
[21] SNELLING T J, GENC B, MCKAIN N, et al. Diversity and community composition of methanogenic archaea in the rumen of Scottish upland sheep assessed by different methods.[J]. Plos One, 2014, 9(9):e106491.
[22] TAJIMA F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism.[J]. Genetics, 1989,123(3):585-595.
[23] MCCABE M S, CORMICAN P, KEOGH K, et al. Illumina miseq phylogenetic amplicon sequencing shows a large reduction of an uncharacterised succinivibrionaceae and an increase of the Methanobrevibacter gottschalkii clade in Feed Restricted Cattle.[J]. Plos One, 2015,10(7):e0133234.
[24] WALLACE R J, ROOKE J A, MCKAIN N, et al. The rumen microbial metagenome associated with high methane production in cattle[J]. BMC Genomics, 2015,16(1):839.
[25] ABECIA L, RAMOSMORALES E, MMARTINEZFERNANDEZ G, et al. Feeding management in early life influences microbial colonisation and fermentation in the rumen of newborn goat kids[J]. Animal Production Science, 2014,54(9):1 449-1 454.
[26] BIAVATI B, VASTA M, FERRY J G. Isolation and characterization of “Methanosphaera cuniculi” sp. nov. [J]. Applied and Environmental Microbiology, 1988,54(3): 768-771.
[27] FRICKE W F, SEEDORF H, HENNE A, et al. The genome sequence of Methanosphaera stadtmanae reveals why this human intestinal archaeon is restricted to methanol and H2 for methane formation and ATP synthesis [J]. Journal of Bacteriology, 2006,188(2): 642-658.
[28] LIU Y, WHITMAN W B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea [J]. Annals of the Aew York Academy of Sciences, 2008,1125(1): 171-189.
[29] HOEDT E C, CUIV P, EVANS P N, et al. Differences down-under: alcohol-fueled methanogenesis by archaea present in Australian macropodids[J]. Isme Journal, 2016,10:2 376-2 388.
[30] POULSEN M, SCHWAB C, JENSEN B B, et al. Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen[J/OL]. Nature Communications, 2013,4:1 428. doi:10.1038/ncomms2432.
[31] RUBINO F, CARBERRY C, WATERS S M, et al. Divergent functional isoforms drive niche specialisation for nutrient acquisition and use in rumen microbiome[J]. Isme Journal, 2017,11(4):932-944.
[32] KAMKE J, KITTELIMAN S, SONI P, et al. Rumen metagenome and metatranscriptome analyses of low methane yield sheep reveals a Sharpea-enriched microbiome characterised by lactic acid formation and utilisation[J]. Microbiome, 2016,4(1):56.
猜你喜欢
土壤学报(2022年3期)2022-08-26 12:15:26
大自然探索(2022年5期)2022-07-11 03:10:33
知识就是力量(2022年6期)2022-06-16 20:19:36
智富时代(2019年5期)2019-07-05 18:43:32
黑龙江八一农垦大学学报(2017年5期)2017-10-31 06:17:12
湖南农业科学(2017年7期)2017-03-04 02:01:03
河北建筑工程学院学报(2015年2期)2015-04-29 12:23:52
河北建筑工程学院学报(2015年4期)2015-03-24 09:48:11
应用海洋学学报(2014年4期)2014-11-22 07:43:56
环境工程技术学报(2014年5期)2014-07-19 06:44:22
