
NQO1在tBHQ干预急性CO中毒后迟发性脑病大鼠海马区的表达及作用
2018-06-21 09:14张亦雯张益梅李经伦
中风与神经疾病杂志 2018年5期
吴 沙,吕 霞,何 林,张亦雯,张益梅,李经伦
含碳物质的不完全燃烧生成过量一氧化碳(Carbon monoxide,CO),人体吸入过量CO后引起的中毒称急性一氧化碳中毒(Acute carbon monoxide poisoning,ACOP),是临床工作中常见的急危重症病,致死致残率极高。在生活生产中,通气不良的环境下以煤气中毒、炭火燃烧、工业泄漏等形式威胁着人们的生活。急性一氧化碳中毒迟发性脑病(delayed encephalophathy after acute carbon monoxide poisoning,DEACMP)即神经精神后发症,是指约10%~30%急性CO中毒患者早期即使经成功救治意识障碍恢复后,仍表现出在一定时间的“伪恢复期”(2~60 d)后出现以精神心理行为异常、痴呆症状,以及锥体系、锥体外系症状为主的神经系统症状和体征[1]。因其高致死率、高致残率无疑已成为ACOP患者最为严重的并发症,其发病几率在重度CO中毒中可达10%~40%,预后不佳,给社会和家庭带来严重的经济负担,同时多年来该病的预防及治疗也成为摆在国内外无数临床医师面前的极大挑战。目前对于该病发病机制的研究芸芸众多,十分复杂,但仍无统一定论,且任意一种均不能齐全地解释其具体发病特点。近几年来核因子2-相关因子2/抗氧化反应元件(nuclear factor erythroid 2 p45-related factor 2/anti-oxidant response element,Nrf2/ARE)通路在抗氧化应激中发挥着举足轻重的作用,而ACOP中毒的本质为缺氧引发的一系列氧化应激反应,近年来研究表明该通路在DEACMP发病中发挥着重要作用,醌氧化还原酶1(NAD(P)H:quinone oxidoreductase 1,NQO1)作为其下游经典靶基因之一,是调节细胞内物质处于氧化还原状态的黄素酶[2],因其催化醌类物质无单电子中间产物及自由基产生而发挥保护细胞作用已为人们所熟知,但随着研究的深入其弊端也逐渐浮现。该研究通过在成功建立ACOP中毒后迟发性脑病(DEACMP)大鼠模型,运用Nrf2激动剂叔丁基对苯二酚(tertiary buty lhydroquinone,tBHQ)干预大鼠观察其下游靶基因NQO1的表达进一步探讨NQO1在该病发病中发挥的作用,为进一步研究DEACMP 发病机制提供实验参考,致力于减少临床上DEACMP 的发生,为探究其靶向治疗奠定基石、提供指导。
1 材料与方法
1.1 主要的实验材料
1.1.1 实验动物 在成都达硕实验动物有限公司购买健康的清洁级SD雄性大鼠180只,饲养于西南医科大学城北动物实验中心,体重约300 g,生产许可证号:SCXK(川)2016~15。在温度20~25 ℃、湿度约40%~70%、光暗12 h交替环境、给予充足的鼠粮及高压消毒纯净水饲养;
1.1.2 主要实验试剂 在泸州市西南化工研究院购买CO纯品气体(中国),南京华立明化工购买tBHQ试剂(中国),圣克鲁斯生物技术公司的兔抗鼠NQO1多克隆抗体(中国),Roche公司的Western荧光试剂、Tunel试剂盒(德国),Dako公司Envision试剂盒(丹麦)。
1.1.3 主要实验仪器 购自中国成都泰盟软件有限公司的Morris水迷宫实验仪、美国雅培公司的血气分析仪、英国BRIGHT公司的石蜡切片机、美国徕博公司的倒置显微镜、美国ProteinSimple公司的凝胶成像仪。
1.2 实验方法
1.2.1 实验分组 模型建立、干预处理:将饲养并训练1 w的雄性SD大鼠进行Morris水迷宫实验,去掉有学习记忆功能缺失的大鼠,从180只可取的大鼠中随机抽出60只作为空气对照组(AC组),剩余大鼠用于建立迟发性脑病模型,按染毒后1 d、3 d、7 d、14 d、21 d、28 d分为6个亚组,染毒方式如下[3]:首剂CO以100 ml/kg体重腹腔注射,每隔4 h以50 ml/kg体重剂量追加注射,连续追加3次。AC组予以同等方式注射空气。腹腔注射CO的大鼠随机分成TC组60只、CO组60只,按50 mg/kg体重每日一次相同时间点腹腔注射tBHQ溶剂于TC组大鼠,CO组大鼠不做其他处理。
1.2.2 模型判定 动脉血碳氧血红蛋白(HbCO)浓度测定:从TC、CO组每个亚组中随机抽取1只大鼠,AC组中随机抽取1只,按染毒前及注射首剂CO后的不同时间点(15 min,2 h,4 h,6 h,8 h,10 h,12 h,14 h,16 h,20 h)采集0.5 ml股动脉血,血气分析仪测HbCO浓度。采用Morris水迷宫实验方法验证大鼠学习记忆能力的变化。
1.2.3 取材、标本处理 从各组中按每个时间点随机选出一部分大鼠,适量生理盐水进行左心室心脏灌洗,4%多聚甲醛灌注固定后断头取脑,4%多聚甲醛中保存,48 h内冠状位切取含海马、齿状回的脑组织进行石蜡包埋、切片。
1.2.4 免疫组织化学法检测NQO1的表达 烤片脱蜡、梯度酒精水化,PBS冲洗,内源性过氧化物酶失活,枸橼酸纳缓冲液抗原修复,加一抗室温1 h,4 ℃过夜,37 ℃复温45 min,PBS冲洗后加二抗,辣根过氧化物酶酶标,DAB显色3~10 min,显微镜下调控染色水平,苏木精复染,脱水透明封片,镜下观察,采用人工半定量计数法对阳性表达细胞进行统计分析[4]。
1.2.5 Western Blot检测NQO1 蛋白 提取蛋白,配胶,加样,变性离心,放入4 ℃冰箱备用,上样、电泳、转膜、封闭后进行免疫反应、显色,经凝胶成像仪成像,以B-actin为参考值,得出NQO1条带密度相对值,与内参值比较得出目标条带灰度值,进行统计分析。
1.2.6 TUNEL染色 梯度酒精脱蜡,滴加试剂酶标、DAB 显色,复染后脱水透明、封片,40×10倍光镜下观察海马CA1区细胞凋亡情况,算出凋亡指数进行统计分析。
2 结 果
2.1 染毒后行为学表现 与AC组比较,TC组、CO组大鼠染毒后不久(<10 min)即有躁动不安、过度呼吸表现,CO追加次数增多后发生精神萎靡、四肢乏力、呼吸困难、末梢循环缺氧表现,部分大鼠甚至发生肢体强直抽搐、大小便失禁、丧失意识、死亡(8只)。保持通风良好的空气环境中完成操作,及时补充相应数目大鼠的同时将死亡的大鼠剖腹解剖,其内脏不同程度肿胀、颜色暗红、表面散在出血点,剥开颅骨发现其脑组织弥漫性充血水肿。其余存活大鼠摄食减少、精神萎靡、易受惊吓。Morris水迷宫实验结果显示中毒大鼠随染毒时间延长其活动路径以随机、自由、散漫形式多见,毫无规律。染毒后14 d始平均潜伏期较AC组明显延长、平台象限活动时间、穿越平台频次明显减低(P<0.05),直至28 d,TC组上述变化明显较CO组显著(P<0.05)。
2.2 动脉血HbCO浓度变化 染毒前各组大鼠动脉血HbCO浓度均<1%,首剂注射CO 15 min后,HbCO浓度攀升达65%左右,2 h时仍高于60%,4 h时降至55%左右,此时进行追加半剂CO后测HbCO浓度继续攀升至70%左右,而后下降走势,整个染毒过程大鼠体内HbCO浓度>50%持续超过16 h。
2.3 海马区NQO1的表达
2.3.1 免疫组织化学结果 AC组大鼠海马CA1区NQO1蛋白表达极少,各时间点无明显差异。TC组、CO组大鼠海马CA1区NQO1蛋白的阳性细胞在40×10倍显微镜下胞核和胞质出现深浅不一的棕褐色或棕黄色,呈上升-峰点(3 d)-下降的走势,TC组峰点较CO组峰点更高更明显,TC组、CO组组内染毒后3 d与其他时间点比较及各组间各相同时间点比较差异具有统计学意义(见图1,见表1)。

图1 A、B、C分别为染毒后3 d,AC组、CO组、TC组大鼠海马CA1区NQO1蛋白表达情况,阳性细胞为棕褐色或棕黄色
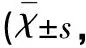
表1 3组大鼠海马CA1区不同时间点NQO1免疫组化阳性结果比较分)
注:与AC组比较各相同时间点*P<0.05;与CO组比较△P<0.05;组内:TC组内▽P<0.05;CO组内▲P<0.05
2.3.2 Western Blot结果 AC组大鼠海马NQO1蛋白表达不显著,CO组、TC组大鼠海马中NQO1的表达(见图2)。
2.4 Tunel染色 AC组大鼠凋亡细胞少,各时间点无明显变化。TC组、CO组大鼠染毒后均出现不同程度的细胞凋亡现象,从1 d开始逐步攀升,7 d达峰点,而后逐渐下降,且组内峰期凋亡情况与其他时间点比较差异具有统计学意义。达峰点(7 d)前TC组较CO组细胞凋亡指数下降,达峰点之后TC组较CO组细胞凋亡指数升高,除14 d外其他各时间点组间比较差异具有统计学意义(见图3,表2)。

图2 Western Blot结果显示NQO1的表达条带
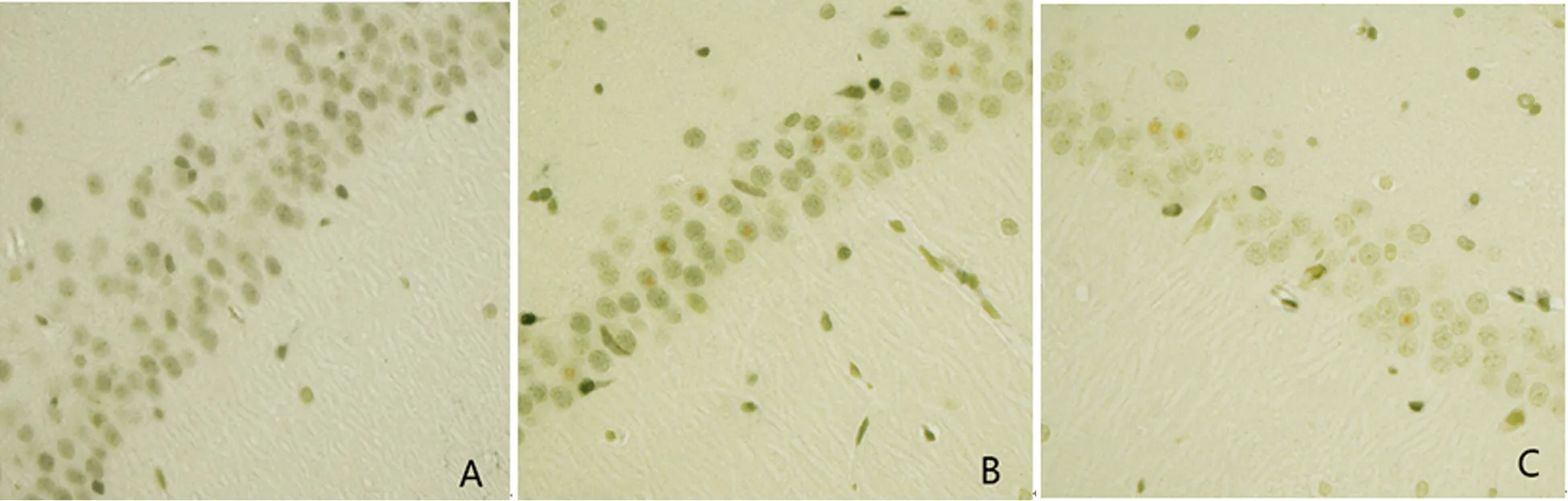
图3 A、B、C分别为染毒后7 d的AC组、CO组、TC组大鼠海马CA1区细胞凋亡情况,胞内含棕色颗粒的细胞为凋亡细胞

表2 3组大鼠海马CA1区不同时间点细胞凋亡指数的比较
注:与AC组比较各相同时间点*P<0.05;与CO组比较△P<0.05;TC组内▽P<0.05,CO组内▲P<0.05
3 讨 论
NQO1(醌氧化还原酶1、DT-diaphorase)是真核生物细胞内广泛存在的一种可诱导的黄素蛋白酶,最早由Ernster 和Navazio于1958年报道[5]。胞质中多见,由NADH或NADPH提供电子催化醌类及其衍生物发生双电子还原反应,催化反应过程中无单电子还原产物或自由基生成故能保护细胞免受氧化应激损伤[6],可被缺氧环境、抗氧化剂、多环芳烃、偶氮染料等诱导表达,这种可诱导性使得NQO1在肿瘤预防及治疗上发挥举足轻重的作用,缺氧和高氧化应激是肿瘤微环境的标志性特征之一,研究发现NQO1在消化、呼吸、生殖、皮肤系统等多种肿瘤性疾病中高表达提示预后不良,患者生存率降低[7,8]。DEACMP实质是缺氧诱发氧化应激反应后导致的一系列脑功能障碍疾病,抗氧化剂还可通过生物异源性物质反应元件(XRE)激活 NQO1基因的表达,NQO1主要存在于细胞质中,以NADH或NADPH作为电子供体,催化包括醌、醌亚胺、亚甲蓝、二氯靛酚、谷胱甘肽取代的蔡醌、偶氮和硝基化合物等等在内的物质发生还原反应,不生成亲电子还原产物及自由基,可解毒醌为毒性较低的氢醌类化合物而清除,避免对细胞的损伤,是公认的解毒防癌酶[9]。NQO1参与维生素E介导的抗氧化,充当辅酶Q的还原酶,维持P53蛋白稳定从而起到保护神经元细胞免受氧化应激以及一系列其他内源性毒素损伤的效果[10];在慢性神经系统变性疾病如:帕金森病和肌萎缩侧索硬化中诱导NQO1的表达上调能够起到很好的神经保护作用[11],阿尔茨海默病(AD)患者脑组织中大量神经细胞丢失、凋亡,神经细胞外老年斑沉积,神经纤维缠结,其中就富含NQO1[12]。且有动物实验结果发现[13]NQO1基因敲除小鼠脑出血发生率及神经细胞损伤严重程度显著高于野生型小鼠。
而近年来有研究发现,经NQO1还原而生成的产物因其结构及稳定性差异而可能对细胞产生相应的毒副作用[14]。经NQO1催化生成的产物氢醌对细胞凋亡影响存在量效、时效关系,当氢醌在细胞内达一定浓度或持续作用一定时间后不仅诱导细胞凋亡还引起细胞继发性坏死[15]。其机制可能如下:(1)低剂量氢醌可能诱导HSP、peroxiredoxin等保护性蛋白表达以使细胞发生适应性反应保护细胞,但细胞氧化应激反应激烈时,cys发生不可逆的活化,上述保护性蛋白发生过氧化而失活,细胞内氧化还原失衡最终致细胞死亡[16]。(2)氢醌通过激活内源性细胞凋亡途径,促发Caspase级联反应,尤其是Caspase-3、Caspase-7活化,诱导细胞程序化死亡[17,18]。(3)氢醌可激活蛋白激酶B,使其定位于胞质中调控下游信号分子FasL、P27、cyclinE、cyclinB等表达影响细胞周期及发生凋亡作用[19,20]。(4)氢醌可经多途径代谢,代谢过程可产生活性氧(ROS),ROS通过直接参与细胞信号通路,使线粒体膜电位变化促发线粒体细胞凋亡程序[21]。(5)氢醌可激活DNA-PKcs/Akt信号通路,活化Akt(蛋白激酶B)进一步推动氧化应激反应,氧化应激正反馈促进Akt活化,增强细胞对ROS诱发细胞凋亡的作用[22]。
tBHQ为广泛应用于人们生活生产中的抗氧化剂,同时是受Nrf2/ARE调节的二相解毒酶诱导剂,能调节该通路下游靶基因NQO1、HO-1、GSH等的表达[23],有实验发现20 μmol/LtBHQ处理细胞,胞核中Nrf2在加药后8 h表达最佳,其下游靶蛋白NQO1在胞质中加药后16 h表达最佳,撤去tBHQ后其诱导表达持续4 h[24]。故本实验采用染毒后每日一次相同时间点腹腔注射经乙醇溶解的浓度为5 mg/mL的tBHQ,达到其持续诱导NQO1表达的效果。另外本实验采用成熟的改良式腹腔注射CO法建立ACOP中毒迟发性脑病大鼠模型,使大鼠体内动脉血HbCO浓度>50%至少持续达16 h之久,能达到类似临床中重度CO中毒患者的程度,更利于DEACMP发病[25]。
Morris水迷宫实验发现TC、CO组大鼠在染毒后前7 d并未出现智能障碍,这与临床DEACMP患者发病经历潜伏期相似。Tunel染色发现TC组大鼠出现不同程度的细胞凋亡,染毒后前7 d凋亡指数逐步攀升,7 d达高峰期,TC组与CO组、AC组相同时间点比较差异具有统计学意义,CO组大鼠细胞凋亡也为类似走势但TC组凋亡指数均低于CO组,且高于AC组。免疫组化结果显示染毒后大鼠海马区NQO1的表达从1 d开始逐渐增加,3 d达峰点,且TC组与CO组、AC组每个相同时间点比较差异均具有统计学意义,TC组大鼠海马CA1区NQO1的表达明显高于CO组、AC组。我们有理由相信在大鼠急性一氧化碳中毒后体内发生了明显的氧化应激反应,染毒前期(染毒后3 d)通过每日一次不间断运用Nrf2激动剂tBHQ,NQO1一定程度高表达的同时细胞凋亡指数有所下降,通过稳定某些保护性蛋白活性、减少自由基生成、减少蛋白酶体降解等作用发挥正性防御作用从而阻止大鼠高级神经功能损伤。
而Morris水迷宫实验发现14 d开始TC、CO组大鼠穿越平台频次减少,活动路径随机自由散漫,日趋严重,提示大鼠发生了进行性的学习记忆能力障碍。而此时TUNEL染色发现7~28 d两组大鼠均发生明显细胞凋亡现象,免疫组化结果显示染毒3 d后NQO1持续高表达、缓慢下降走势,且TC组与CO组、AC组每个相同时间点比较均明显增高,差异均具有统计学意义。同时TUNEL染色结果显示此时段TC组细胞凋亡指数逐渐较CO组升高,直至28 d水迷宫实验结果显示TC组高级神经功能损伤重于CO组,提示在染毒后期尽管细胞内NQO1表达增高发挥了足够保护作用但仍不能阻止其毒性作用持续、细胞凋亡进展直至凋亡高峰出现,脑细胞因凋亡大量丢失致神经功能缺损,TC组中NQO1持续高表达不但不能发挥正性保护作用反而进一步加重细胞凋亡,高级神经活动发生进行性损害。这与Lee等发现细胞在缺血缺氧环境下,NQO1蛋白的持续过表达抑制细胞存活甚至促进细胞凋亡的观点不谋而合,其机制可能是持续过表达的NQO1激活AMPK通路,而后遏制mTOR信号通路有关[26]。因免疫组化仅对海马CA1区NQO1表达进行了统计,为增加结果的准确性我们对整个海马的NQO1表达进行Western Blot法检测,得出的结果与免疫组化结果一致。TC、CO组凋亡高峰出现时间推后于NQO1蛋白表达峰点,或许与细胞功能变化到细胞器实质发生变化所需时间有关,而其行为学的改变基本与明显细胞凋亡发生时间平行,进一步说明在DEACMP发病机制中海马区细胞发生的迟发性凋亡起着举足轻重的作用。
大鼠急性CO中毒后,对缺氧反应灵敏的海马区细胞受到氧化应激作用,前期NQO1表达升高通过稳定保护性蛋白活性、减少自由基生成等途径防御脑细胞损伤,但后期NQO1持续高表达导致其氧化还原产物氢醌生成达一定浓度、堆积达一定时间诱导凋亡途径的发生,且细胞持续缺氧难以抵抗钙离子集聚、炎症介质释放等的发生,甚至推动细胞迟发性凋亡的发生发展。经Pearson 直线相关分析显示CO及TC组NQO1表达与凋亡指数趋势一致,CO组NQO1与细胞凋亡呈正相关(r=0.505,P<0.001),TC组NQO1与细胞凋亡无相关性(r=0.0049,P<0.001),实质上TC组中两者并不是没有相关性,而是NQO1升高与细胞发生凋亡之间密切程度不高,即NQO1高表达在染毒后前期发挥抗凋亡作用和后期发挥促凋亡作用更显著,将两时段结合分析得出相关性不高的结果。
近年来临床中仍有大量急性CO中毒所致精神心理行为异常、智能减退、运动障碍、甚至瘫痪痴呆、尿便失禁、生活不能自理的患者,严重影响患者及家人的生活质量,DEACMP致病病理生理途径多样且繁杂,是临床工作者持续面临的挑战。研究发现Nrf2/ARE通路在其中起着至关重要的作用,其下游抗氧化蛋白NQO1在一定时间内一定程度的发挥着正性积极作用,而随着NQO1持续高表达后对细胞的消极负性作用逐渐浮出水面。本实验为进一步研究其致病机制及靶向治疗提供了实验基础,为临床减少DEACMP的发生和研究其靶向治疗提供一定的基础和方向。
[参考文献]
[1]Yueh-Feng S,Chen MH,Peng GS,et al. Generalized chorea due to delayed encephalopathy after acute carbon monoxide intoxication[J]. Annals of Indian Academy of Neurology,2015,18(1):108-110.
[2]Hu HG,Scholten I,Gruss C,et al. The distribution of the DEK protein in mammalian chromatin[J]. Biochemical & Biophysical Research Communications,2007,358(4):1008-1014.
[3]李勇军,刘 强,赵春梅,等. 改良式腹腔注射CO法建立急性CO中毒迟发脑病大鼠模型[J]. 宁夏医科大学学报,2012,34(4):334-336.
[4]赵风华,庄 玉,呼格吉乐图. 绒山羊肥大细胞类胰蛋白酶的免疫组化及图像分析研究[J]. 畜牧与饲料科学,2012,33(1):8-11.
[5]Smith MT. Benzene,NQO1,and genetic susceptibility to cancer[J]. Proceedings of the National Academy of Sciences of the United States of America,1999,96(14):7624-7626.
[6]Radjendirane V,Joseph P,Lee YH,et al. Disruption of the DT diaphorase (NQO1) gene in mice leads to increased menadione toxicity[J]. Journal of Biological Chemistry,1998,273(13):7382-7389.
[7]Ma Y,Kong J,Yan G,et al. NQO1 overexpression is associated with poor prognosis in squamous cell carcinoma of the uterine cervix[J]. BMC Cancer,2014,14(1):414.
[8]Yang Y,Yan Z,Wu Q,et al. Clinical implications of high NQO1 expression in breast cancers[J]. Journal of Experimental & Clinical Cancer Research Cr,2014,33(1):14.
[9]Siegel D,Gustafson DL,Dehn DL,et al. NAD(P)H:Quinone Oxidoreductase 1:Role as a Superoxide Scavenger[J]. Molecular Pharmacology,2004,65(5):1238-1247.
[10]Shrader WD,Akiko A,Adam B,et al. Towards a modern definition of vitamin E-evidence for a quinone hypothesis[J]. Bioorganic & Medicinal Chemistry Letters,2012,22(1):391-395.
[11]Shih AY,Li P,Murphy TH. A small-molecule-inducible Nrf2-mediated antioxidant response provides effective prophylaxis against cerebral ischemia in vivo[J]. Journal of Neuroscience the Official Journal of the Society for Neuroscience,2005,25(44):10321.
[12]Wang YX,Santa-Cruz K,De Carli C,et al. NAD(P)H:quinone oxidoreductase activity is increased in hippocampal pyramidal neurons of patients with Alzheimer’s disease [J]. Neurobiol Aging,2000,21:525-531.
[13]Stringer JL,Gaikwad A,Gonzales BN,et al. Presence and induction of the enzyme NAD(P)H:quinone oxidoreductase 1 in the central nervous system. [J]. Journal of Comparative Neurology,2004,471(3):289-297.
[14]Munday R,Smith BL,Munday CM. Effect of inducers of DT-diaphorase on the toxicity of 2-methyl- and 2-hydroxy-1,4-naphthoquinone to rats[J]. Chemico-Biological Interactions,1999,123(3):219-237.
[15]Mcguinness SM,Johansson R,Lundstrom J,et al. Induction of apoptosis by remoxipride metabolites in HL60 and CD34+/CD19- human bone marrow progenitor cells:potential relevance to remoxipride-induced aplastic anemia[J]. Chemico-Biological Interactions,1999,121(3):253-265.
[16]Chevallet M,Wagner E,Luche S,et al. Regeneration of peroxiredoxins during recovery after oxidative stress:only some overoxidized peroxiredoxins can be reduced during recovery after oxidative stress. [J]. Journal of Biological Chemistry,2003,278(39):37146.
[17]Kuribayashi K,Mayes PA,Eldeiry WS. What are caspases 3 and 7 doing upstream of the mitochondria[J]. Cancer Biology & Therapy,2006,5(7):763-765.
[18]Zhang F,Lau SS,Monks TJ. A Dual Role for Poly(ADP-Ribose) Polymerase-1 During Caspase-Dependent Apoptosis[J]. Toxicological Sciences,2012,128(1):103-114.
[19]Brunet A. Bonni A. Zigmend MJ,et al. Akt promotes cell survival by phospherylating and inhihiting a frokhead transcription factor [J]. Cell,1999,96:857-868.
[20]Zhu J,Wang H,Yang S,et al. Comparison of toxicity of benzene metabolite hydroquinone in hematopoietie stem cells derived from murine embryonic yolk sae and adult bone marrow[J]. PLoS One,2013,8:e71153.
[21]Ying MD,Tu CX,Ying HZ,et al. MSFTZ,a flavanone derivative,induces human hepatoma cell apoptosis via a reactive oxygen species-and caspase-dependent mitochondrial pathway[J]. Journal of Pharmacology and Experimental Therapeutics,2008,325:758-765.
[22]Nogueira V,Park Y,Chen CC,et al. Akt determines replicative senescence and oxidative or oncogenic premature senescence and sensitizes cells to oxidative apoptosis[J]. Cancer Cell,2008,14(6):458-470.
[23]Keum YS,Han YH,Liew C,et al. Induction of heme oxygenase-1(HO-1)and NAD [P]H:quinone oxidoreductase 1 (NQO1)by a phenolic antioxidant,butylated hydroxyanisole (BHA)and its metabolite,tert -butylhydroquinone (tBHQ)in primary -cultured human and rat hepatocytes[J]. Pharm Res,2006,23:2586-2594.
[24]武小媛,曲丽艳,全 康,等. tBHQ和Sulforaphane对Caco2细胞Nrf2-ARE信号通路的影响[J]. 浙江大学学报(医学版),2010,39(1):17-23.
[25]Sun Q,Cai J,Zhou J,et al. Hydrogen-rich saline reduces delayed neurologic sequelae in experimental carbon monoxide toxicity[J]. Critical Care Medicine,2011,39(4):765-769.
[26]Lee H,Oh ET,Choi BH,et al. NQO1-induced activation of AMPK contributes to cancer cell death by oxygen-glucose deprivation[J]. Sci Rep,2015,5:7769.
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
科学导报·学术(2020年19期)2020-07-09
创新作文(小学版)(2019年10期)2019-09-25
意林原创版(2019年6期)2019-06-30
癌变·畸变·突变(2018年6期)2018-12-07
环境与生活(2018年7期)2018-09-10
小学生优秀作文(低年级)(2018年3期)2018-02-08
小学生时代·综合版(2016年4期)2016-11-19
少年文艺·开心阅读作文(2016年11期)2016-09-10
意林(2009年1期)2009-08-11
