
封育对荒漠草原两种植物群落土壤种子库的影响
2018-06-19 01:06李国旗邵文山赵盼盼靳长青
草业学报 2018年6期
李国旗,邵文山,赵盼盼,靳长青
(1.宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;2.宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021)
土壤种子库作为植被种群生活史的一个阶段,在植被更新与恢复、生物多样性维护、植被演替和扩散过程中有着不可低估的作用[1-2]。土壤种子库在植被经历干扰后是其恢复的主要种源,在退化草地的恢复与重建中承担着重要的角色,而且能够反映种群对环境变化的响应[3]。植被演替的过程不仅包括地上植被组成与结构的变化,同时也包括土壤种子库的数量与格局的变化[4]。荒漠草原生态系统是草原生态系统向荒漠生态系统过渡的一个最干旱的草原生态系统类型;随着全球气候变化、人为开发过度等因素,作为我国草原生态系统的重要组成部分的荒漠草原,目前也承受着巨大的压力[5-6]。宁夏草地面积约为300万hm2左右,其中荒漠草原的面积超过了近一半;而宁夏盐池县地处典型的农牧交错带,县境由东南至西北为广阔的干草原和荒漠草原,生态系统十分脆弱,特别是在近几十年来,由于自然因素及人类活动干扰等因素,草原植被退化、沙化非常严重。自2000年以来,宁夏在全区范围内实施了以封育禁牧为主的一系列生态保育措施,已经取得了一定的成效。由于荒漠草原是宁夏草原的主体类型,因此宁夏退牧还草的效果也主要取决于荒漠草原的恢复程度[7]。
苦豆子(Sophoraalopecuroides)是一种多年生豆科草本植物,茎直立,奇数羽状复叶,总状花序顶生,荚果串珠状,具多数卵球形种子。芨芨草(Achnatherumsplendens)是一种多年生禾本科草本植物,须根粗壮具沙套,秆直立,圆锥花序开展,花果期6-9月。所选的苦豆子群落中出现了15种植物,隶属于9科15属,其中以禾本科的居多,有5种;在芨芨草群落中出现的植物有14种,隶属于7科11属,其中菊科有4种,禾本科和藜科各有3种。苦豆子群落土壤容重较低,土壤全氮含量相对较高;芨芨草群落土壤pH明显较高,土壤有机碳含量相对较高。苦豆子和芨芨草群落的土壤磷酸酶活性较高,而蔗糖酶活性普遍较低[8]。苦豆子和芨芨草是宁夏盐池县荒漠草原极端盐渍化生境的代表性植物,研究这两种植物群落封育前后土壤种子库的变化特征,为封育禁牧措施的评价和荒漠草原极端生境中植物群落恢复潜力的评估提供了参考依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏盐池县沙边子(E 107°24′,N 37°43′)地区,属于典型的荒漠草原区,是陕、甘、宁、蒙4省(区)的交界地带,地处毛乌素沙地西南边缘处,海拔1300~1370 m,自然条件较为恶劣,属于典型的中温带大陆性气候,年降水量250 mm左右,且主要集中在6-9月,潜在年蒸发量达2500 mm左右,年平均气温7.7 ℃,年均无霜期165 d。地形主要为缓坡丘陵,土壤类型主要为灰钙土、风沙土等,土壤质地以风沙土为主[8]。研究区封育植被主要有苦豆子、芨芨草、盐爪爪(Kalidiumfoliatum)、白刺(Nitrariatangutorum)和黑沙蒿(Artermisiaordosica)等。
1.2 研究方法
1.2.1样地选择 试验样地位于盐池县沙边子试验站。该样地自2000年围栏封育后,一直未曾放牧。于2015年7月,选取围栏封育内外苦豆子群落和芨芨草群落类型草地(表1),在每块样地分别选取地势相对平坦、环境条件较为一致的25 m×30 m的4个大的样方,在每个大样方内布设5条“之”字形样线,每条样线间距5 m,每条样线上距5 m取1个1 m × 1 m的样方,每条样线上取6个样方,共计120个样方进行植被调查。

表1 样地基本特征Table 1 Main features of sampling plots
1.2.2取样与试验处理 于2016年3月在植被调查的4块大样方内,各随机选择1 m×1 m的小样方10块,用20 cm×20 cm×10 cm的取土器,分别取0~2 cm、2~5 cm、5~10 cm土层的土,将同一大样方内所取的2份土样均匀混合在一起,共计60个土样,带回实验室自然风干,过筛,进行种子萌发试验。本试验中种子库的鉴定主要采取种子过筛直接辨认和种子萌发试验鉴定[9]。其中种子辨认鉴定主要是肉眼可见部分种子,即通过与已知种子对比进行辨认;种子萌发是本试验种子库鉴定的主要方式,即用无种子的毛细沙做基质置于花盆(大小:42 cm×28 cm×15 cm)底部(120 ℃高温处理),约10 cm,再将土样过筛去除杂物后均匀平铺在萌发用的花盆中(土层1~2 cm),萌发时间为2016年5到9月,开始萌发后可辨认的幼苗鉴定后去除,无法辨认的继续生长直至鉴定出,萌发2个月翻动土样,至9月连续4周没有幼苗萌发,结束试验。
1.3 计算方法
将鉴定的土壤种子数量换算成1 m2种子数量[10],根据土壤种子库物种和种子密度计算Shannon-Wiener多样性指数、Simpson 多样性指数、Pielou均匀度指数、Margalef丰富度指数[11-12]。
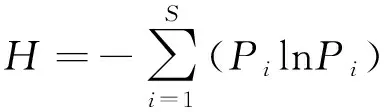
式中:N为物种的个体总数;S为物种数目;Pi为第i个物种占总物种数的比例。
采用Sorensen相似性系数计算土壤种子库封育内外群落组成的相似性[13]。
Sc=2w/(a+b)
式中:Sc为相似性系数;w为样地共有植物种数;a和b为两样地各自拥有的植物种数。
1.4 数据处理
用Excel 2007和SPSS 17.0进行数据统计和部分图表制作,用LSD最小显著差法进行显著性分析。
2 结果与分析
2.1 封育内外土壤种子库的物种组成
由表2可知,2种植物群落封育内外土壤种子库物种组成、同一物种密度组成均存在差异。苦豆子群落封育内土壤种子库中有23种物种,种子密度达4550粒·m-2,隶属于5科19属,其中禾本科和藜科物种数占主要优势,分别达到39.13%和30.43%,种子密度分别占32.42%和36.59%,地上优势种苦豆子在土壤种子库中密度为525粒·m-2,占种子密度的11.54%。苦豆子群落封育外土壤种子库中存在19种物种,种子密度为4710粒·m-2,隶属于6科16属,禾本科植物种数下降仅有3种,种子密度为17.62%,藜科植物种数占优势达31.58%,其种子密度占28.45%,菊科植物有增加趋势,其种子密度占到28.13%,出现蒺藜科、萝藦科植物种,苦豆子种子比例下降。芨芨草群落封育内土壤种子库植物种有29种,种子密度达5350粒·m-2,隶属于9科24属,其中菊科、藜科、禾本科植物种较多,占总物种数的65.52%,种子密度达到80.09%,地上优势种芨芨草在土壤种子库中仅占4.32%。芨芨草群落封育外土壤种子库中物种数有26种,种子密度为4285粒·m-2,隶属于8科22属,菊科、藜科、禾本科仍为土壤种子库的主要物种,占到69.23%,种子密度达到80.86%,旋花科、远志科植物消失,新增加大戟科地锦植物种,地上优势种芨芨草种子比例变化不大。
2.2 土壤种子库垂直分布格局
封育对2种植物群落土壤种子库的垂直分布产生了一定的影响(图1)。封育后2种植物群落土壤种子库不同土层物种数均呈增加趋势;而随着土层深度的增加,物种数逐渐下降。此外,芨芨草群落封育内外土壤种子库物种数均高于苦豆子群落封育内外土壤种子库物种数。封育不仅对2种植物群落封育内外土壤种子库物种数在垂直分布上产生了影响,对土壤种子库数量在垂直分布上也产生了影响。由表3 可知,封育对2种植物群落土壤种子库数量影响不同,2种植物群落封育内外随着土层的加深,土壤种子库数量均呈下降趋势,苦豆子群落土壤种子库种子数量总体上未封育种子数量高于封育内土壤种子库,而芨芨草群落中则反之;但封育对2种群落不同土层土壤种子数量分布的影响有所差异,表层土(0~2 cm)中2种植物群落封育内土壤种子库种子数量均高于未封育土壤种子库,而苦豆子群落中亚表层(2~5 cm)和深层土(5~10 cm)中未封育土壤种子库中种子数高于封育土壤种子库中的种子数,芨芨草群落封育内外土壤亚表层和深层中土壤种子库种子数量分布特征与苦豆子群落中相反。
2.3 土壤种子库多样性分析
由表4可知,封育内2种植物群落土壤种子库多样性、均匀度和丰富度指数均高于封育外,说明围栏封育可提高植物群落土壤种子库的多样性、均匀度和丰富度。芨芨草群落封育内外土壤种子库多样性、均匀度和丰富度指数均分别高于苦豆子群落土壤种子库,说明芨芨草群落可能在研究区的适应度相对更高。此外,芨芨草群落封育外土壤种子库的Margalef丰富度指数高于苦豆子群落封育内,但其Shannon-Wiener 多样性指数、Simpson 多样性指数和Pielou 均匀度指数低于苦豆子群落封育内,说明苦豆子群落封育后土壤种子库的多样性、均匀度明显增加,其丰富度虽有所增加但在一定时间内增加范围有限,可能与群落生存生境本身有关。
2.4 土壤种子库相似性分析
由表5可知,封育对群落土壤种子库相似性变化有影响,且对不同植物群落土壤种子库相似性的变化影响不同。芨芨草群落封育内外间土壤种子库相似性最高,但苦豆子群落封育内外土壤种子库相似性低于苦豆子群落封育内与芨芨草群落封育外间的相似性,说明不同植物群落土壤种子库对封育措施的响应有所差异,芨芨草群落封育后相似性明显提高,苦豆子群落对封育的响应程度并不高。芨芨草群落封育内土壤种子库与苦豆子群落封育内外土壤种子库相似性均相对较低,低于封育外土壤种子库与苦豆子群落封育内外土壤种子库的相似性,这一方面反映出苦豆子群落在荒漠草原退化中要早于芨芨草群落出现,另一方面在植被恢复中,苦豆子群落有向芨芨草群落演替的趋势。


图1 不同土层不同植物群落土壤种子库的物种数Fig.1 Species number of SSB of different communities in different soil layer

项目 Item0~2 cm 2~5 cm 5~10 cm 苦豆子内 S. alopecuroides inside3075±342Aa950±105Bb530±77Ba苦豆子外 S. alopecuroides outside2895±481Aa1245±75Bab570±110Ba芨芨草内 A. splendens inside2795±460Aa1940±265Ba615±94Ca 芨芨草外 A. splendens outside2665±392Aa1205±469Bab415±173Ca
注:不同大写字母表示同一群落处理不同土层差异显著,不同小写字母表示同一土层不同群落处理间差异显著(P<0.05)
Note: Different capital letters indicate the significant difference in same sample plots among different soil layers, and the different lowercase letters indicate the significant difference between two sample plots (P<0.05).
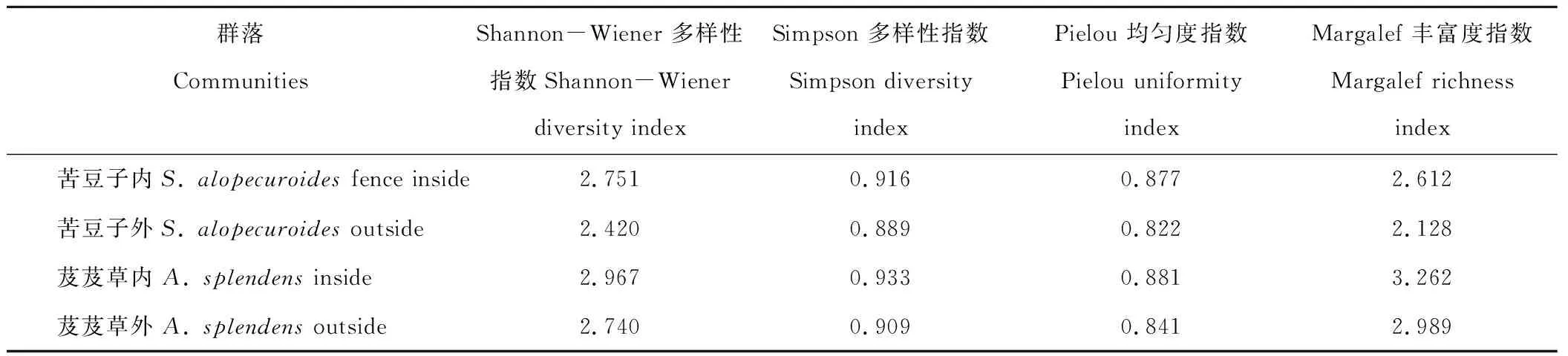
表4 不同群落土壤种子库多样性、均匀度和丰富度指数Table 4 Indexes of diversity, richness and pielou of the SSB in different communities

表5 不同群落土壤种子库相似性分析Table 5 Similarity analysis of SSB in different communities
3 讨论
3.1 封育内外土壤种子库的物种组成
不同草地类型的土壤种子库差异较大,用不同的研究方法也会导致研究结果的不同[1,14]。Silvertown[15]研究表明草地土壤种子库的范围在103~106粒·m-2,本试验中土壤种子库的规模符合Silvertown[15]的研究结果。封育3年的高寒草地群落,封育内外3种高寒草地群落土壤种子库物种数目差异不显著,不同草地类型封育内外土壤种子库密度变化不同[1]。还有研究结果认为退化样地土壤种子库种子密度显著高于封育样地[16]。目前有关荒漠草原上土壤种子库对封育措施的响应研究较少。本研究发现封育措施提高了2种植物群落中的物种数目;苦豆子群落封育内土壤种子密度低于封育外,但差异不大,芨芨草群落封育内土壤种子密度明显高于封育外,说明荒漠草原不同植物群落土壤种子库物种数目及种子密度对封育措施的响应不同,这和在高寒草地的研究结果类似,可能一方面与地上植物的繁殖策略不同及土壤本身特性不同有关,另一方面可能还与封育年限、研究方法不同有关,有关这方面的研究还需要进一步验证。此外,本研究中,苦豆子群落封育内外土壤种子库中藜科植物物种数比例均较高,封育后禾本科植物明显增加,菊科植物减少,蒺藜科、萝藦科植物消失;芨芨草群落封育内外土壤种子库中藜科、禾本科、菊科植物种均占优势,封育后旋花科、远志科植物出现,大戟科植物消失,说明封育改变了土壤种子库中潜在植物种群的结构。
3.2 土壤种子库垂直分布格局
土壤种子库具有明显的垂直分布格局,这种立体结构影响着种子库种子的萌发和留存,进而影响着未来植被的恢复与重建[17]。Bekker等[18]提出在没有遭到破坏的土壤生境中,0~5 cm土壤层中的种子垂直分布符合一定的数学模型,有研究表明[19]土壤种子在土壤层中呈二项式分布或泊松分布。本研究中随着土层的加深,2种植物群落封育内外土壤种子库中种子数量和物种数均呈递减趋势,且种子数量递减显著(P<0.05),这与大多数的研究结果一致[17,20-22];封育后2种植物群落不同土层物种数均有增加,说明封育一定程度上可增加群落物种,但具体封育时间与不同群落土壤种子库中物种数量的变化关系还需要进一步研究。此外本研究中,芨芨草群落封育后土壤种子库各土层种子数量均增加,说明封育可提高种子数量;但在苦豆子群落土壤种子库中土壤表层和深层封育内反而低于封育外,可能是因为封育后下层土壤生境改善有利于种子的萌发,也可能是封育后干扰减少,种子储藏在土壤表层;封育对2种群落土壤种子库数量的垂直分布影响不同,说明封育对不同植物群落土壤种子库的影响不同,可能一方面与地上植被种子大小、繁殖策略等有关,另一方面封育后土壤生境有变化影响了种子的萌发与储藏。
3.3 土壤种子库多样性分析
本研究表明,围栏封育可增加不同植物群落土壤种子库的多样性、均匀度和丰富度,这与有关封育对土壤种子库影响的研究结果相一致[16,23-24],有学者对典型草原封育过程中土壤种子库的变化进行了研究,结果表明土壤种子库多样性和物种丰富度变化与封育年限有关[25];而在高寒草地,封育3年的3种群落土壤种子库在封育后围栏内外多样性、物种丰富度、均匀度变化不同[1]。本试验中,2种植物群落封育后土壤种子库多样性、均匀度及丰富度虽然均增加,但不同植物群落对封育的响应有所差异,2种植物群落围栏内外多样性特征的不同,芨芨草群落在荒漠草原的稳定性更高,而具体有关封育年限对不同植物群落土壤种子库的多样性特征影响还需要进一步研究。
3.4 土壤种子库相似性分析
群落相似性可以在一定程度上表征群落间相互关系,其研究对群落替换、群落演替具有重要意义。本研究中,2种植物群落封育内外土壤种子库间的相似性相对较高,一方面荒漠草原植物种本身相对较少,大多是适应度较高的植物种,有关苦豆子和芨芨草作为伴生群落的研究也有报道[26-27];另一方面与种子传播有关,荒漠草原自然条件更有利于种子的水平传播。不同植物群落封育内外相似性不同,说明封育对不同植物群落土壤种子库的影响不同。此外本研究中,苦豆子群落封育内与芨芨草群落封育外土壤种子库间的相似度较高,说明苦豆子群落封育后有向芨芨草群落演替的趋势。
4 结论
主要研究了围封内外苦豆子群落和芨芨草两种植物群落的土壤种子库物种组成、种子密度、多样性及相似性等特征。初步结论如下:(1)封育可提高两种植物群落土壤种子库中物种数目,且苦豆子群落无论是围封还是未围封样地,其土壤种子库物种数均比芨芨草群落的多;(2)围封对两种植物群落土壤种子库种子密度的影响不同,封育内外苦豆子群落种子密度变化较小,但封育能显著提高外芨芨草群落种子密度;(3)随着土层的加深,两种植物群落封育内外土壤种子库种子密度均呈递减趋势,且表层土壤(0~2 cm)的种子密度均显著高较深的2个土层;(4)封育可提高两种植物群落土壤种子库的多样性、均匀度和丰富度。两种植物群落封育内外土壤种子库相似性相对较高,苦豆子群落封育后有向芨芨草群落演替的趋势。
参考文献References:
[1] Deng B, Ren G H, Liu Z Y,etal. Effect of 3-year fencing on soil seed banks of three alpine grassland communities. Acta Prataculturae Sinica, 2012, 21(5): 23-31.
邓斌, 任国华, 刘志云, 等. 封育三年对三种高寒草地群落土壤种子库的影响. 草业学报, 2012, 21(5): 23-31.
[2] Yang N, Fu M Y, Yang M Y,etal. Soil seed bank properties under different land used models on sloping land with purple soils in Hengyang of Hunan Province, south central China. Acta Botanica Boreali Occidentalia Sinica, 2014, 34(11): 2324-2330.
杨宁, 付美云, 杨满元, 等. 衡阳紫色土丘陵坡地不同土地利用模式下土壤种子库特征. 西北植物学报, 2014, 34(11): 2324-2330.
[3] Zhou H K, Zhou L, Liu W,etal. Influence of fencing on degradedKobresiahumilismeadows and nondegraded. Grassland of China, 2003, 25(5): 15-22.
周华坤, 周立, 刘伟, 等. 封育措施对退化与未退化矮嵩草草甸的影响. 中国草地学报, 2003, 25(5): 15-22.
[4] Yang L, Wang Y R, Yu J D. Review of research on seed banks of desert regions. Acta Prataculturae Sinica, 2010, 19(2): 227-234.
杨磊, 王彦荣, 余进德. 干旱荒漠区土壤种子库研究进展. 草业学报, 2010, 19(2): 227-234.
[5] Lü G F, Wu Y S, Li H,etal. Microorganisms, soil nutrient and enzyme activity changing with the proceeding of desert steppe degradation in Inner Mongolia. Journal of Desert Research, 2010, 30(1): 104-109.
吕桂芬, 吴永胜, 李浩, 等. 荒漠草原不同退化阶段土壤微生物、土壤养分及酶活性的研究. 中国沙漠, 2010, 30(1): 104-109.
[6] Sun H L. Chinese ecosystems. Beijing: Science Press, 2005: 550-665.
孙鸿烈.中国生态系统.北京: 科学出版社, 2005: 550-665.
[7] Wang G H, Wang J J, Tao L B,etal. Effects of enclosure on soil aggregate distribution and stability of desert steppe in Ningxia. Acta Agrestia Sinica, 2017, 25(1): 76-81.
王国会, 王建军, 陶利波, 等. 围封对宁夏荒漠草原土壤团聚体组成及其稳定性的影响. 草地学报, 2017, 25(1): 76-81.
邵文山, 李国旗, 陈科元, 等. 荒漠草原4种常见植物群落土壤酶活性比较. 西北植物学报, 2016, 36(3): 579-587.
[9] Li F R, Zhao L Y, Wang S F,etal. Effects of enclosure management on the structure of soil seed bank and standing vegetation in degraded sandy grasslands of eastern Inner Mongolia. Acta Prataculturae Sinica, 2003, 12(4): 90-99.
李锋瑞, 赵丽娅, 王树芳, 等. 封育对退化沙质草地土壤种子库与地上群落结构的影响. 草业学报, 2003, 12(4): 90-99.
[10] Ma Q L, Lu Q, Wei L Y,etal. Varying characteristics of soil seed banks during the succession process ofNitrariatangutorumvegetation in an arid desert area. Acta Ecologica Sinica, 2015, 35(7): 2285-2294.
马全林, 卢琦, 魏林源, 等. 干旱荒漠白刺灌丛植被演替过程土壤种子库变化特征. 生态学报, 2015, 35(7): 2285-2294.
[11] Liu R X, Zhan J, Shi Z H,etal. Soil seed bank and its correlations with aboveground vegetation and environmental factors in water level fluctuating zone of Danjiangkou Reservoir, Central China. Chinese Journal of Applied Ecology, 2013, 24(3): 801-808.
刘瑞雪, 詹娟, 史志华, 等. 丹江口水库消落带土壤种子库与地上植被和环境的关系. 应用生态学报, 2013, 24(3): 801-808.
[12] Zhang J T. Quantitative ecology. Beijing: Science Press, 2004: 77-92.
第二,施肥时间和方法。小麦的施肥时期和方法也有着局限性,存在区域差距,小麦施肥应分成底肥和追肥两种,以某县举例:当前传统播肥使用量少,玉米秸秆是麦季使用有机肥的有效途径,秸秆还田的条件上,根据目标施肥,将氮肥、钾肥和磷肥均作为底肥施入,使用方法是撒于表面进行翻耕,有利于发挥肥效。因为氮素的积累量是在小麦的全育期,同时氮肥易挥发和流失,所以在小麦施肥过程中应将氮肥作为追肥在拔节时期追施,方法可使用随水冲施。另外,观察土壤养分是否丰缺,主要观察是否添加微量元素肥料。
张金屯. 数量生态学.北京: 科学出版社, 2004: 77-92.
[13] Yan R R, Wei Z J, Xin X P,etal. Effects of grazing systems on germinable soil seed bank of desert steppe. Journal of Desert Research, 2011, 31(3): 703-709.
闫瑞瑞, 卫智军, 辛晓平, 等. 放牧制度对荒漠草原可萌发土壤种子库的影响. 中国沙漠, 2011, 31(3): 703-709.
[14] Shang Z H, Long R J, Ma Y S,etal. Soil seed banks of degraded alpine grassland in headwater region of the Yellow River: quantities and dynamics of seed germination. Chinese Journal of Applied & Environmental Biology, 2006, 12(3): 313-317.
尚占环, 龙瑞军, 马玉寿, 等. 黄河源区退化高寒草地土壤种子库: 种子萌发的数量和动态. 应用与环境生物学报, 2006, 12(3): 313-317.
[15] Silvertown J W. Introduction to plant population ecology. London and New York: Longman, 1982: 22-37.
[16] Ma M J, Zhou X H, Lü Z W,etal. A comparison of the soil seed bank in an enclosed vs. a degraded alpine meadow in the eastern Tibetan Plateau. Acta Ecologica Sinica, 2009, 29(7): 3658-3664.
马妙君, 周显辉, 吕正文, 等. 青藏高原东缘封育和退化高寒草甸种子库差异. 生态学报, 2009, 29(7): 3658-3664.
[17] Li H Y, Mo X Q, Hao C. A review of study on soil seed bank in the past thirty years. Ecology and Environmental Sciences, 2009, 18(2): 731-737.
李洪远, 莫训强, 郝翠. 近30年来土壤种子库研究的回顾与展望. 生态环境学报, 2009, 18(2): 731-737.
[18] Bekker R M, Bakker J P, Grandin U,etal. Seed size, shape and vertical distribution in the soil: indicators of seed longevity. Functional Ecology, 1998, 12(5): 834-842.
[19] Ambrosiol, Doradoj, Delmonte J P. Assessment of the sample size to estimate the weed seed bank in soil. Weed Research, 1997, 37: 129-137.
[20] Zeng Y J, Wang Y R, Nan Z B,etal. Soil seed banks of different grassland types of Alashan arid desert region, Inner Mongolia. Chinese Journal of Applied Ecology, 2003, 14(9): 1457-1463.
曾彦军, 王彦荣, 南志标, 等. 阿拉善干旱荒漠区不同植被类型土壤种子库研究. 应用生态学报, 2003, 14(9): 1457-1463.
[21] Ma M, Ma Z, Du G. Effects of water level on three wetlands soil seed banks on the Tibetan Plateau. Plos One, 2014, 9(7): e101458.
[22] Ma M, Zhou X, Du G. Effects of disturbance intensity on seasonal dynamics of alpine meadow soil seed banks on the Tibetan Plateau. Plant and Soil, 2013, 369(1): 283-295.
[23] Zhao L P, Cheng J M, Wan H E,etal. Characteristics of soil seed banks in grassland under fencing and grazing in Loess Plateau. Pratacultural Science, 2008, 25(10): 78-83.
赵凌平, 程积民, 万惠娥, 等. 黄土高原草地封育与放牧条件下土壤种子库特征. 草业科学, 2008, 25(10): 78-83.
[24] Huang X Y, Wang K, Wang Y T,etal. Soil seed bank dynamics during natural restoration by fence in a typical steppe of Inner Mongolia. Acta Agrestia Sinica, 2011, 19(1): 38-42.
黄欣颖, 王堃, 王宇通, 等. 典型草原封育过程中土壤种子库的变化特征. 草地学报, 2011, 19(1): 38-42.
[25] Shen Y, Liu C F, Ma H B,etal. Response of a soil seed bank to modes of grassland management on a desert steppe. Acta Ecologica Sinica, 2015, 35(14): 4725-4732.
沈艳, 刘彩凤, 马红彬, 等. 荒漠草原土壤种子库对草地管理方式的响应. 生态学报, 2015, 35(14): 4725-4732.
[26] Liu X K, Liu M S, Huang Z,etal. Pattern analysis of inter specific relationships in four arid communities in sand lake Ningxia Hui Autonomous region, China. Chinese Journal of Plant Ecology, 2009, 33(2): 320-330.
刘小恺, 刘茂松, 黄峥, 等. 宁夏沙湖4种干旱区群落中主要植物种间关系的格局分析. 植物生态学报, 2009, 33(2): 320-330.
[27] Chen L M G, Liu M S, Huang Z,etal. Distribution pattern of root biomass and inter specific relationship inAchnatherumsplenoens-Sophoraalopecuroidescommunity in Northwest China. Chinese Journal of Plant Ecology, 2009, 33(4): 748-754.
晨乐木格, 刘茂松, 黄峥, 等. 我国西北地区芨芨草——苦豆子群落根系分布与种间关系. 植物生态学报, 2009, 33(4): 748-754.
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
第二课堂(课外活动版)(2022年2期)2022-04-23
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
绿色科技(2021年10期)2021-06-23
植物保护(2020年3期)2020-06-08
绿色中国(2019年14期)2019-11-26
草业学报(2019年2期)2019-02-25
土壤(2017年1期)2017-04-24
儿童故事画报·智力大王(2016年7期)2017-02-08
