
低温诱导胁迫下不同烟草品种电导率及抗氧化酶活性的变化
2018-06-19 13:19王欣亚金大翔朴世领
延边大学农学学报 2018年1期
尹 航, 王欣亚, 金大翔, 万 悦, 朴世领
(延边大学农学院,吉林延吉133002)
低温胁迫是植物体受到胁迫的一种,容易导致农作物细胞内生理生化指标失衡,急剧胁迫下更会导致农作物死亡从而减产,是除土壤成分和水成分以外农作物生长与产量的另一个重要影响因素[1]。对植物叶片的抗寒性研究发现,随着胁迫温度的下降,植物叶片细胞膜受损加剧呈“S”型变化[2],抗氧化酶活性和丙二醛含量均呈先上升后下降趋势[3-4]。品种的抗寒性与相对电导率、抗氧化酶活性和丙二醛含量密切相关[5-6]。
烟草是通过叶片带来经济效益的农作物,最适生长温度为24~28 ℃,10~6 ℃则会抑制其生长。我国的烟草种植区域面积广且各地温度的差异大,按照生态生长环境分为烟草种植最适宜区、适宜区、次适宜区和不适宜区[7]。吉林省烟草大田移栽时期一般为5月中下旬,此期正处于早春冷害发生时期。因此当地早春冷害的发生对烟草的品种选择及田间栽培管理带来了极大挑战。低温冷害会使烟草生理生化指标出现紊乱,不仅影响烟株的正常生长,而且还会导致烟叶化学成分比例严重失调,从而会使烟叶的产量和品质下降。本研究选取6种烟草品种作为试材,研究其在低温胁迫下生理生化指标的变化,为对烟草抗寒品种的选择及合理应用提供一定的理论参考,也为进一步探索烟草的抗寒机理提供理论依据。
1 材料与方法
1.1 供试品种、烤烟品种
分别为吉烟9号(当地主推品种)、NC95、云87;晒烟品种分别是延晒7号(当地主推品种)、漂河1号(当地地方品种)、建平大人头。
1.2 试验处理及设计
试验所需种子播种前用浓度为0.02%赤霉素浸种24 h之后播种于25 cm×60 cm的装有基质(腐殖土、草炭土、细沙为10∶10∶1)的育苗盘中,在温室内育苗,3叶龄时选择壮苗移至假植简塑盘(100株/盘)。苗期达到5~6叶龄时,开始低温处理,每个处理选取150株形态及长势基本一致的健壮的烟苗,将其转移至人工气候箱(QHX-40085-Ⅲ)。
低温胁迫设置10、8、6、4、2、0 ℃,以24 ℃常温培养为对照,做6个温度处理。首先在24 ℃预处理2 d,然后以2 ℃/h的下降速率,达到10 ℃时开始低温胁迫,每个低温胁迫处理24 h,随后又以0.5 ℃/h下降速率,降至下一个温度进行低温胁迫,以此类推。
1.3 取样及测定方法
1.3.1 取样
每梯度低温胁迫后,选择长势一致的5~6叶龄烟苗,选取由上至下第3和4片烟草叶片作为混合样品,每次取样3次重复。
1.3.2 相对电导率的测定
先用蒸馏水将叶片表面清洗干净,再用去离子水冲洗1遍,用滤纸吸干表面水分,将叶片剪成1~3 mm宽度的长条(避开主脉),快速称取3份,每份0.1 g,分别置于10 mL去离子水的刻度试管中。用ORION TDS型电导仪测定叶片电导率。
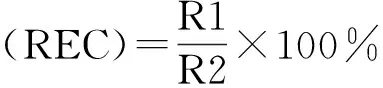
式中,R1:室温下浸提液浸泡处理12 h后,浸提液电导率。R2:水浴锅沸水浴30 min后,冷却至室温浸提液电导率。
1.3.3 半致死温度(LT50)计算
利用SPSS统计分析软件对不同处理温度与烟草叶片相对电导率之间的关系做线性回归方程[8-9]。
1.3.4 抗氧化酶活性及MDA含量的测定
过氧化物酶(POD)采用愈创木酚氧化比色法测定[10],过氧化氢酶(CAT)采用紫外分光光度法测定[10],超氧化物歧化酶(SOD)参照氮蓝四唑光化还原法测定[11],MDA含量采用硫代巴比妥酸(TBA)法测定[12]。
2 结果与分析
2.1 低温胁迫下不同烟草品种叶片相对电导率的变化
不同烟草品种苗期低温诱导处理下电导率变化见图1。随着低温胁迫加剧,6个烟草品种叶片的相对电导率均呈递增趋势。可见烟草叶片相对电导率与低温诱导温度呈负相关,叶片细胞膜伤害不断加重,电导率不断升高。低温诱导10~8 ℃时,6种烟草品种叶片相对电导率均平稳上升;而8~4 ℃时,烟草叶片电导率急剧上升;随后4 ~0 ℃时,又平缓上升,且在所有低温处理下各烟草品种间增幅均存在显著性差异(P<0.05)。6 ℃时云烟87增幅最大,与对照相比增长了4.28倍,延晒7号增幅最小,仅增长了1.52倍;4 ℃时增幅由大到小依次为云烟87、NC95、建平大人头、吉烟9号、漂河1号和延晒7号。由此说明,延晒7号、漂河1号受低温伤害较小,抗寒能力较强;吉烟9号、建平大人头受低温伤害次之,抗寒能力一般;NC95和云烟87受低温伤害较大,抗寒能力较弱。
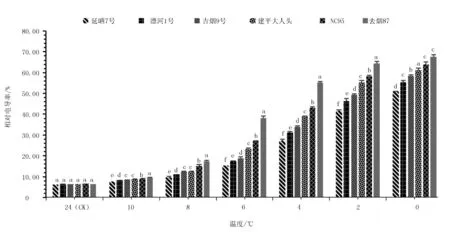
图1 不同烟草品种苗期低温诱导处理下电导率变化
2.2 低温胁迫下不同品种烟草的半致死温度
由表1可知,利用线性方程对相对电导率与处理温度进行拟合,拟合值(R2)为0.943 3~0.889 6,表明拟合结果比较精确可靠,能够较准确的反应出两者之间存在的线性关系,并能够算出半致死温度(LT50/℃),半致死温度由小到大依次是延晒七号(0.86 ℃)、漂河一号(1.5 ℃)、吉烟九号(1.89 ℃)、建平大人头(2.64 ℃)、NC95(3.08 ℃)和云烟87(5.17 ℃)。由此说明延晒7号、漂河一号有较强的抗寒性,而云烟87和NC95抗寒性最弱。吉烟九号、建平大人头抗寒性介于两者之间。

表1 6种烟草品种线性方程和半致死温度
2.3 不同烟草品种叶片在低温胁迫下生理生化指标的变化
2.3.1 低温胁迫对烟草叶片SOD活性的影响
SOD是生物体内能够消除新陈代谢产生的有害物质的活性酶[13-14]。由表2可知,随着温度的降低,6种烟草的SOD活性均呈先缓慢上升后急剧下降的趋势。从10~8 ℃时,各品种SOD活性缓慢上升,6~2 ℃时,开始急剧下降,各品种均在8 ℃时达到峰值。低温处理8 ℃时延晒7号增幅最大,为对照组的1.46倍,且与其他品种之间均有显著性差异(P<0.05),而漂河1号、建平大人头、吉烟9号、NC95和云烟87增幅均较小,分别为对照组的1.3、1.14、1.12、1.12和1.05倍,而它们之间无显著性差异(P>0.05)。6 ℃时SOD活性增长幅度由大到小依次为延晒7号、漂河1号、吉烟9号、建平大人头、NC95和云烟87。其中,吉烟9号与建平大人头增幅无显著性差异(P>0.05),但与其他品种间均存在显著性差异(P<0.05)。有试验表明,SOD活性增长幅度越大,品种抗逆性越强[15],延晒7号在整个处理期间增幅均最大,而云烟87增幅最小。由此可知,延晒7号抗寒性最强,而云烟87抗寒性最弱。
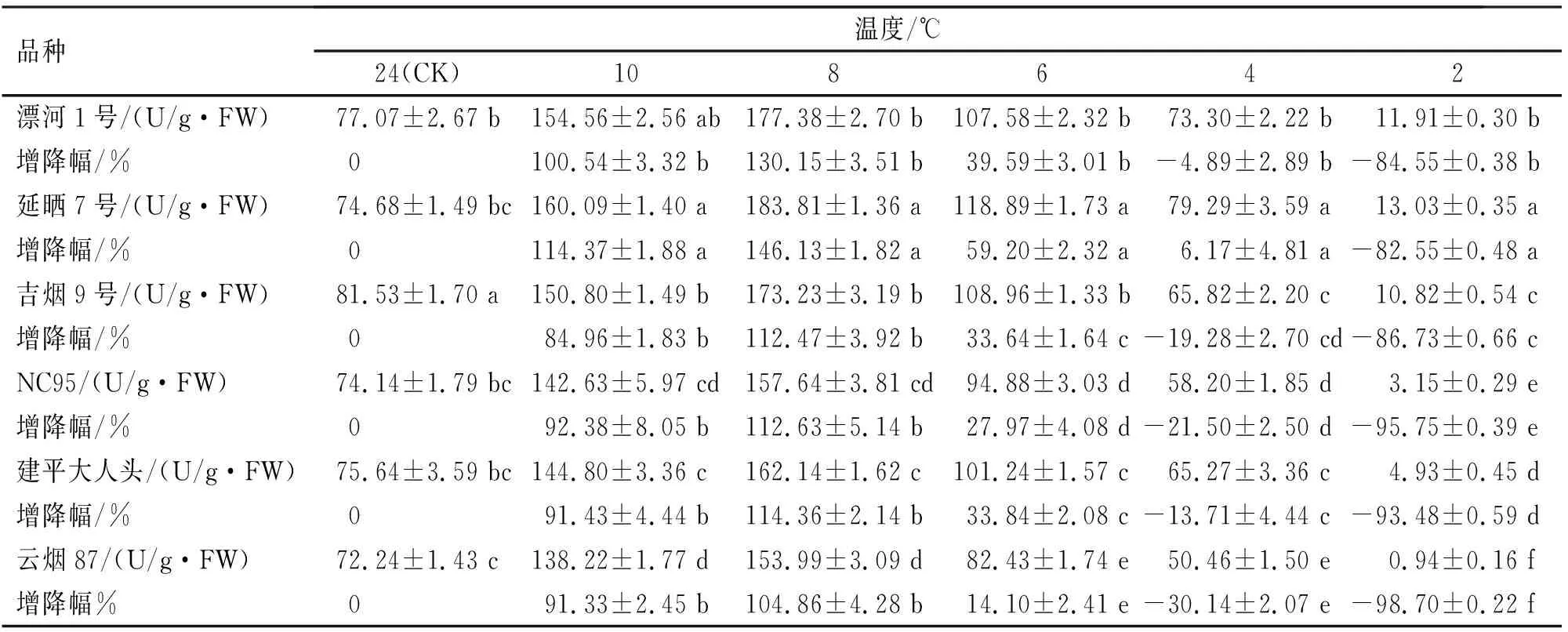
表2 不同烟草品种苗期低温诱导处理SOD活性变化
2.3.2 低温胁迫对烟草叶片POD活性的影响
POD也是植物体内用于清除自由基的抗氧化酶,它与植物蒸腾作用及生长素调节等都有关系[16]。由表3可知,低温胁迫期间,各品种POD活性变化与SOD活性变化规律基本一致,而达到峰值的低温有所不同。6种烟草品种均在10 ℃时达到峰值,随后在8~2 ℃期间持续下降。低温胁迫10 ℃时,漂河1号与吉烟9号之间增幅无显著性差异(P>0.05),但与其他品种间均存在显著性差异(P<0.05)。其中,延晒7号增幅最大,为对照组的0.72倍;漂河1号和吉烟9号次之,分别为0.63倍和0.49倍;建平大人头和NC95较小,为0.36倍和0.26倍;云烟87最小,仅为对照组的0.16倍。6 ℃时,POD活性与对照相比均出现降幅现象。其中,延晒7号降幅最低仅为对照组的0.1倍,而云烟87降幅最大为0.4倍。说明延晒7号具有更强的清除活性氧及自由基的能力,能更好地抵御低温伤害;而云烟87反之。

表3 不同烟草品种苗期低温诱导处理POD活性变化
2.3.3 低温胁迫对烟草叶片CAT活性的影响
植物受到胁迫后,会开始积累过氧化氢,此时,CAT迅速启动并发挥清除过量H2O2的作用,当CAT活性增幅开始下降时,则植物抗逆性也会随之减弱[17]。低温胁迫对不同烟草CAT活性的影响见表4。
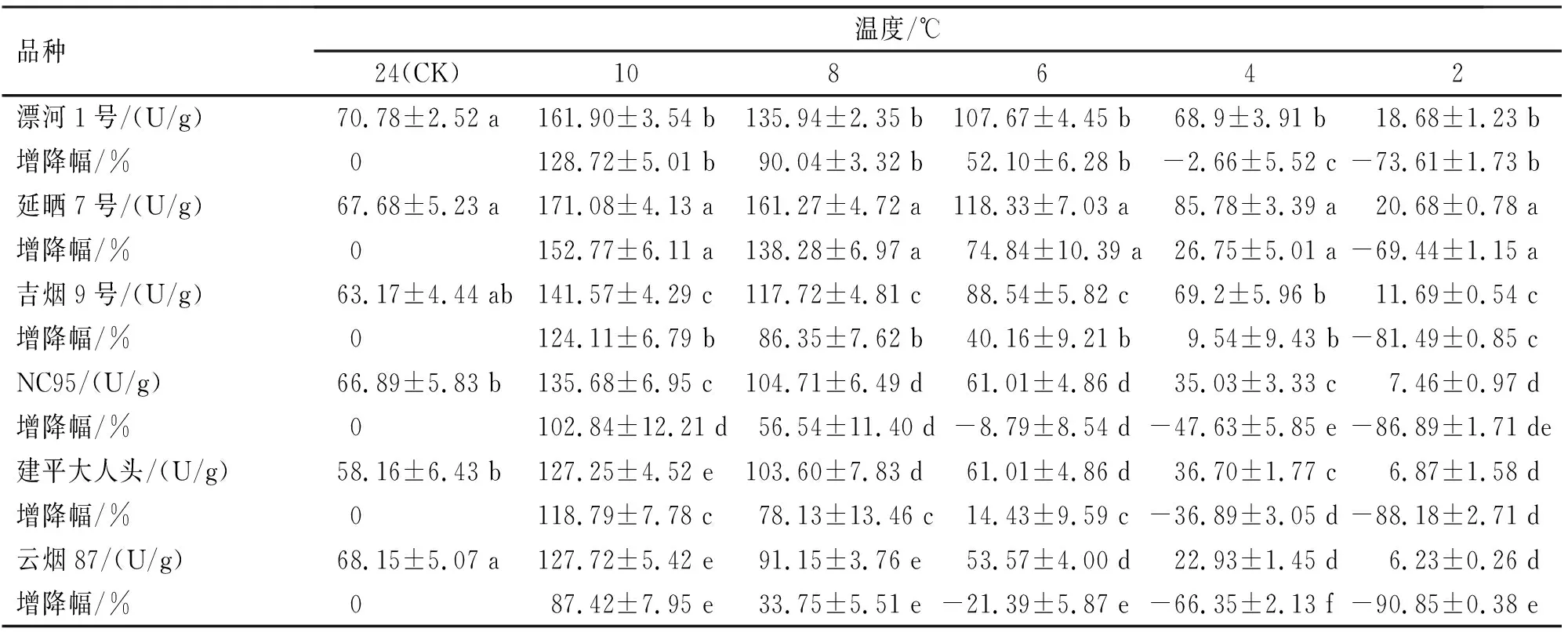
表4 不同烟草品种苗期低温诱导处理CAT活性变化
6种烟草品种叶片CAT活性随低温胁迫的加剧呈现先升高后下降的趋势。在低温诱导10 ℃时,6种烟草品种CAT活性均达到峰值,延晒7号增幅最大,为对照组的1.53倍,云烟87最小,仅为0.87倍。10 ℃时,延晒7号与其他5个品种间均有显著性差异(P<0.05),漂河1号和吉烟9号与其他4种品种之间均有显著性差异(P<0.05),但二者之间无显著性差异(P>0.05),建平大人头、云烟87和NC95之间也存在显著性差异(P<0.05);6 ℃处理时,NC95和云烟87出现降幅现象,而延晒7号、漂河1号、吉烟9号和建平大人头CAT活性增幅仍处于上升趋势。降低幅度由大到小依次为延晒7号(74.84 %)、漂河1号(52.1 %)、吉烟9号(40.16 %)、建平大人头(14.43 %)、NC95(-8.79 %)和云烟87(-21.39 %),且各品种之间均存在显著性差异(P<0.05);4 ℃时,除延晒7号以外,其他5个烟草品种均出现降幅,其中,延晒7号增幅最大,而云烟87增幅最小。且各品种间均存在显著性差异(P<0.05)。有分析指出,CAT增幅越大,表明品种抗逆性越强[18]。由此表明,延晒7号低温诱导处理下CAT酶抗寒性最强,漂河1号、吉烟9号、建平大人头、NC95次之,云烟87抗寒性较弱。
2.3.4 低温胁迫对烟草叶片MDA含量的影响
MDA是细胞膜脂过氧化最重要的产物之一,它能够增大细胞膜的损伤,导致植物衰老,是植物生理生化研究中常用的指标。低温胁迫对烟草叶片MDA含量的变化见表5。随着处理温度的下降,6种烟草品种MDA含量均呈先增高后下降的趋势,其达到峰值的温度有所差异。前人研究表明,植物的抗寒性与MDA含量有关,即峰值出现越晚、增幅越低,则抗寒性越强[19]。其中,云烟87在8 ℃时达到峰值,且增长幅度最大,NC95和建平大人头均在6 ℃时达到峰值,且增长幅度次之,而延晒7号、漂河1号和吉烟9号较晚,均在4 ℃时达到峰值,且增长幅度均较低。从MDA含量增幅来看,各品种间均存在显著性差异(P<0.05)。在低温胁迫8 ℃时,云烟87增幅最大,为对照组的1.67倍;延晒7号最小,仅为1.31倍;2 ℃时,各品种MDA含量均出现降幅,降幅最大的是云烟87,最小的是延晒7号。由此可见,延晒7号细胞膜脂过氧化产生的MDA含量最少,细胞膜损伤最轻,抗寒性最强;漂河1号、吉烟9号和建平大人头、NC95次之;而云烟87产生的MDA含量最多,细胞膜损伤最重,抗寒性最弱。
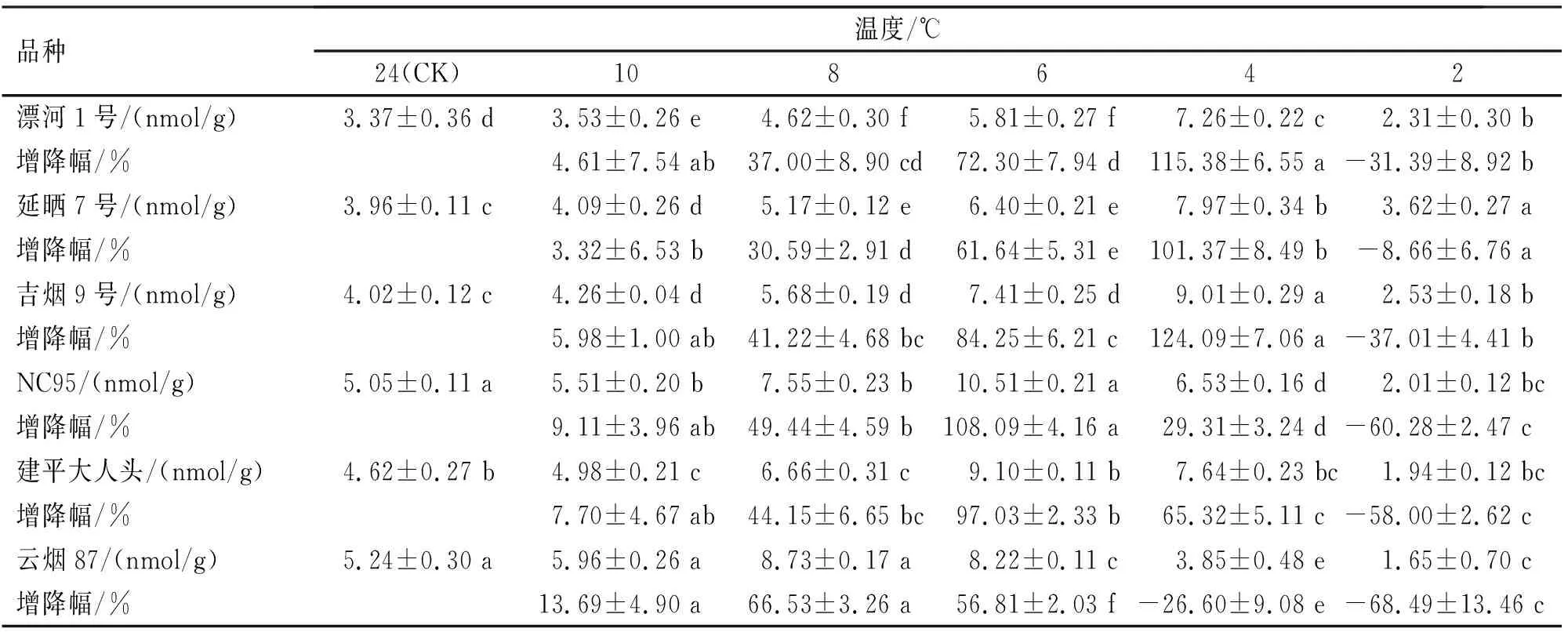
表5 不同烟草品种苗期低温诱导处理MDA含量变化
3 讨论与结论
植物细胞膜是植物受到低温胁迫最初感受器官,也是评价植物品种间抗逆性强弱的重要指标[20]。低温胁迫会导致植物细胞膜系统紊乱,细胞内物质渗透量随低温胁迫加剧呈负相关,渗透越大,相对电导率越大;抗氧化酶活性也会随之增强,在临近半致死温度时又开始下降;细胞膜脂过氧化产物MDA会加大细胞膜损伤,含量越高增幅越强,抗逆性越弱[21]。
本试验所选6种烟草品种叶片的相对电导率也是随处理温度的降低呈上升趋势,表明受低温胁迫越重,烟草叶片细胞受伤害越严重。通过线性方程所得到各品种的半致死温度由小到大依次是延晒七号(0.86 ℃)、漂河一号(1.5 ℃)、吉烟九号(1.89 ℃)、建平大人头(2.64 ℃)、NC95(3.08 ℃)和云烟87(5.17 ℃)。
本试验结果表明,各品种烟草叶片3种抗氧化酶活性均随着温度的下降呈先升高后下降的趋势,低温胁迫的加剧积累了过量的活性氧,导致细胞内物质失衡,加剧了膜脂过氧化作用。延晒7号的抗氧化酶活性始终显著高于同期的NC95、云烟87等品种,表明延晒7号的抗寒性最强;漂河1号、吉烟9号、建平大人头、NC95次之;云烟87最弱,这与王家川等[17]对低温诱导下不同烤烟品种相关酶变化的研究结果基本一致。植物的抗寒性与MDA含量有关,即峰值出现越晚、增幅越低,抗寒性越强。云烟87在8 ℃时达到峰值且增长幅度最大,NC95和建平大人头均在6 ℃时达到峰值且增长幅度次之,而延晒7号、漂河1号和吉烟9号较晚,在4 ℃达到峰值且增长幅度均较低,这与陈有军等[19]研究结果相一致。所以,烟草膜脂间MDA含量达到峰值越早,增幅越强,抗逆性越弱;反之,峰值出现的越晚,增幅越弱,抗逆性越强。
试验结果表明,在抗寒性表现上,当地培育品种优于外地引进品种,且晒烟品种优于烤烟品种。结合烟草叶片相对电导率、半致死温度、抗氧化酶活性和MDA含量的增长幅度来看,延晒7号、漂河1号抗寒能力较强;吉烟9号、建平大人头抗寒性中等;云烟87和NC95抗寒能力较弱。所以,本地培育品种更适用于当地气候环境,可选做杂交亲本,用于烟草抗寒品种的改良和筛选,提高后代的抗寒能力。
参考文献:
[1] 秦立金.光照时间对黄瓜叶片结构及光合特性的研究[J].赤峰学院学报(自然科学版),2016,32(15):25-27.
[2] 屈春媛,金毅,金喜军,等.苗期抗旱花生品种的筛选[J].黑龙江八一农垦大学学报,2016,28(05):1-8.
[3] 令凡,焦健,李朝周,等.不同油橄榄品种对低温胁迫的生理响应及抗寒性综合评价[J].西北植物学报,2015,35(03):508-515.
[4] 尉晶.浅析盐碱胁迫对燕麦种子萌发和幼苗的影响[J].黑龙江八一农垦大学学报,2017,29(05):13-15,88.
[5] 徐卫平,蒋景龙,任绪明,等.低温胁迫对3种柑橘幼苗细胞膜及渗透调节的影响[J].分子植物育种,2017,15(03):1104-1108.
[6] 蒲媛媛,孙万仓.白菜型冬油菜抗寒性与生理生化特性关系[J].分子植物育种,2010,8(02):335-339.
[7] 彭聪.近50年吉林东部地区农业气候条件变化及农业生产对气候变化的响应研究 [D].长春:东北师范大学,2010.
[8] 刘锦,王挺黎,念林,等.电导法配合Logistic方程测定樱花抗寒性研究[J].江苏林业科技,2016,05(43):25-27,31.
[9] 令凡,李朝周,回振龙,等.应用电导率法及Logistic方程测定油橄榄品种的抗寒性[J].广东农业科学,2015,42(01):13-17.
[10] 王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[11] 沈文飚,徐朗莱,叶茂炳,等.氮蓝四哇光化还原法测定超氧化物歧化酶活性的适宜条件[J].南京农业大学学报,1996,19(2):101-102.
[12] 朱广廉,钟诲文,张爱琴.植物生理学实验[M].北京:北京大学高出版社,1990:51-249.
[13] 王鹏翔,艾复清,钟蕾,等.成熟期干旱胁迫对烤烟叶绿素含量及相关酶活性的影响[J].安徽农业科学,2007(31):9945-9946.
[14] 陈卫国.不同烤烟品种抗寒性及相关生理生化指标的研究[D].长沙:湖南农业大学,2007.
[15] 黄希莲,罗充,宋丽莎.低温胁迫对贵阳市9种绿篱植物抗寒性生理生化指标的影响[J].广东农业科学,2012,39(02):47-50,59.
[16] 姚侠妹,偶春,殷明明,等.两种外源物对盐胁迫下栀子幼苗抗氧化酶活性的影响[J].河南科技学院学报(自然科学版),2015,43(05):15-19.
[17] 王家川. 低温诱导下不同烤烟品种相关酶、花芽分化进程及化学成分的变化研究[D].延吉:延边大学,2017.
[18] 袁晓霞,贾娟霞,冀照君,等.盐胁迫条件下蓖麻生长发育研究进展[J].内蒙古民族大学学报(自然科学版),2016,31(01):48-51.
[19] 陈有军,周青平,孙建,等.六份乡土牧草苗期干旱胁迫的对比研究[J].西南民族大学学报(自然科学版),2016,42(06):598-603.
[20] 潘晓云,曹琴东,王根轩.膜脂过氧化作为扁桃品种抗寒性鉴定指标研究(英文)[J].生态学报,2002(11):1902-1911.
[21] 齐志广,赵俊霞.油菜素内酯对大豆苗期生长及硝酸还原酶活性的影响[J].河北师范大学学报,1999(02):132-134.
猜你喜欢
石油库与加油站(2022年3期)2022-08-30
热带作物学报(2022年5期)2022-06-01
矿冶工程(2021年4期)2021-09-15
农业技术与装备(2020年10期)2020-11-03
世界有色金属(2020年15期)2020-10-10
云南农业(2020年5期)2020-06-12
浙江工业大学学报(2017年5期)2018-01-22
中国马铃薯(2015年5期)2016-01-09
诗歌月刊(2014年3期)2015-04-14
传奇故事(破茧成蝶)(2015年8期)2015-02-28
