
光照对反刍动物生产性能与繁殖性能的影响
2018-06-14 00:54毛晨羽史彬林徐元庆王哲奇
动物营养学报 2018年6期
毛晨羽 史彬林 徐元庆 王哲奇
(内蒙古农业大学动物科学学院,呼和浩特 010018)
光照是家畜生长、生产和繁殖的一个重要环境因素,是畜禽环境的重要组成部分,其信号可通过视网膜将神经冲动传递给下丘脑室交叉上核(superior chiasmal nucleus,SCN),然后经过室旁核(paraventricular nucleus,PVN),最后传递到松果体(pineal gland,PG),促使PG分泌褪黑素(melatonin,MLT),进而影响家畜的生理机能和生产性能[1]。光照是家畜保持良好生产和繁殖不可缺少的条件,对于反刍动物而言,更为重要。
1 光照对绒山羊产绒的影响
影响山羊绒形成和生长的因素很多,其中光照是重要的影响因素之一。生产实践表明,羊绒的生长有明显的季节性,每年的秋季开始长绒,长绒速度由慢到块,在冬季11月份长绒速度达到巅峰,到1月份,绒毛的生长基本停止。每年8~12月,绒毛生长总量大约为全年生长量的88%[2-3]。研究表明,山羊产绒量的这种季节性变化,主要是由于产地自然光照周期的季节性变化所致[4]。可见,光照是影响绒山羊绒生长的关键因素。
已有研究表明,在非长绒时期,人工缩短光照周期,可促使山羊长绒。李丰田[5]将山羊按照不同光照时间分为5组,试验时间为3个月,结果对累计羊绒产量产生了极显著影响。在光照对藏西北绒山羊产绒影响的研究中,研究人员发现控制光照可显著提高藏西北绒山羊的产绒量,光照虽然对羊绒纤维直径没有影响,但可显著提高羊绒长度[6]。同样,郭礼祥[7]在光照对陕北白绒山羊绒毛长度影响的研究中也发现,短光照试验组绒山羊羊绒毛长度比自然光照组提高了67.24%,差异显著。在光照对羊绒生长速度及绒品质的研究中,短光照组绒山羊前3个月绒毛生长较快,绒毛生长速度是1.53~1.71 cm/月,显著高于长光照组;随着时间的延长,绒山羊对短光照的敏感性降低,后3个月羊绒生长较慢,绒毛生长速度是0.15~0.86 cm/月,试验组羊绒总长度均超过了纺织工业对绒毛长度的要求;短光照组新生羊绒纤维直径下降幅度较大[8]。
山羊绒生长的季节正好处于日照渐短的秋季,在长光照周期和短光照周期变化的过程中,MLT的分泌表现出一种明显的周期性变化,山羊绒毛的生长周期与MLT的分泌周期一致[9]。因此可以认为光照周期变化及其导致的MLT分泌的变化是影响山羊绒纤维生长的主要因素。Wuliji等[10]简要说明了山羊绒的生长机理:光照作为电子信号在绒山羊神经系统产生作用,使PG的MTL分泌量发生变化,直接影响山羊绒纤维的生长。近年来,通过人工控制光照时间,刺激绒山羊PG,使其增加MLT的分泌,进而使绒山羊在非产绒季节产绒已经成为一种提高年绒产量的常见方法。例如内蒙古农牧科学院营养所的科技人员通过限时放牧、遮光饲养等技术来促使绒山羊的产绒量升高,使绒山羊由秋季长绒变为全年长绒,个体年均产绒量可提高70%[11]。但一些专家认为,山羊实施短日照处理促使其长绒须经过一段长日照过程,否则,短日照将不会对山羊绒生长产生作用[12],这一观点还需要以后进一步的试验研究去验证。
2 光照对奶牛、奶山羊产奶量的影响
自从1987年报告周期性(16 h明期、8 h暗期)光照具有催乳作用以来[13],许多报道指出长光照对产奶量具有促进作用。现有的研究表明,光照能直接影响动物的生理机能,如对奶牛进行光照周期试验,发现不同光照周期能够对奶牛的采食量、产奶量和牛奶的乳脂率产生规律性的影响[14]。光照周期影响奶牛泌乳性能的原因可能主要是长光照周期促进了奶牛催乳素(PRL)和类胰岛素生长因子Ⅰ(IGF-Ⅰ),血液中高浓度的PRL和IGF-Ⅰ是奶牛提高产奶量的关键因素[15]。但PRL与IGF-Ⅰ的分泌受到MLT的影响,随着光照时间的延长,血液中高浓度MLT的维持时间缩短,进而使PRL与IGF-Ⅰ的分泌量增加,促进奶牛产奶。同时,IGF-Ⅰ还可以介导牛生长激素(BST)产生作用[16],BST是牛脑垂体分泌的一种肽类激素,可促进动物生长,外源BST注射可促进奶牛产奶量增多。长光照与BST结合应用,可使产奶量增幅加大。Dahl等[17]发现奶牛在产奶期接受长日照处理之后,干物质采食量会随着奶牛产奶量的提高而增加,但是饲料增加的成本远少于产奶量增加带来的收益。然而值得注意的是,产奶量提高在前,采食量增加在后,所以并不是因为增加光照时间可以让奶牛采食量增加进而导致产奶的提高。此外,密歇根大学的研究人员也证实,假如在泌乳期的最初60 d内,能让奶牛每天接受16 h的光照,就会比相同时期处在短光照的奶牛多产10%~15%的牛奶,与对照组相比差异显著[18]。有趣的是,相比于长光照,在干奶期进行短光照处理,可使奶牛在下一个泌乳期产奶量大幅度增加[19]。Lacasses等[20]的研究结果表明,干奶期和妊娠后期接受短光照处理的奶牛比同一时期接受长光照处理的奶牛,产奶量增加了3.2 kg/d,主要原因可能是短光照对奶牛光照周期反应系统产生了影响。但长光照不是一味地增加光照时间,过长的光照时间有可能会导致激素分泌紊乱,从而使泌乳能力下降[21]。同样,长光照也可以对奶山羊泌乳性能产生影响,但其影响机制现在还不够清楚[22]。以崂山奶山羊为例,当光照时间为13 h时,奶山羊的产奶量均显著高于光照时间为10和16 h时,而乳脂率和乳中干物质含量等指标无显著变化。
3 光照对反刍动物繁殖性能的影响
季节性繁殖动物的繁殖活动受到多种因素的影响,如光照、温度、纬度、营养条件,其中光照起着重要的作用。奶牛虽然是常年发情动物,但与短光照相比,长光照可以对奶牛的繁殖性能起到促进作用,并且在冬季进行长光照会促使母畜发情和提高受孕率[23];绵羊是季节性发情动物,性腺在光照时间逐渐缩短的秋季开始活动。光照信息以电信号的模式通过视神经传到SCN,经SCN调节,传到颈上神经节,再传至PG,使PG分泌MLT[24]。MLT可以调控下丘脑-垂体-性腺轴上相关激素的分泌变化,最终影响绵羊的繁殖。如MLT刺激下丘脑分泌促性腺激素释放激素(GnRH),促进垂体分泌促黄体素(LH)、卵泡生成素(FSH),引起动物发情排卵[25]。大量研究表明,MLT参与绵羊GnRH/LH日变化的调节,在繁殖季节中,母羊血液中MLT与GnRH/LH的每日变化趋势存在着一定联系:日落后母羊血液中GnRH与LH浓度往往上升,如果在14:00注射MLT,在15:00—16:00时对血液中GnRH与LH浓度进行检测,可以发现它们的浓度也会上升;GnRH与LH的浓度不仅存在着日变化,同时也存在着明显的昼夜节律性变化,夜间GnRH与LH浓度显著上升[26]。该现象是绵羊及山羊共有的特性,这可能与夜间MLT浓度升高有关。但是有关这方面的研究报道结果往往不明确,例如在同一个试验中,在日落后绝大部分绵羊繁殖激素浓度有升高的趋势,但仍有部分羊繁殖激素变化没有规律性[27]。
光照在繁殖期与非繁殖期对绵羊在繁殖上有不同的影响,特别是母羊,这主要由机体内部的调节机制在繁殖期与非繁殖期的差异引起的[28]。机体内部存在3种调节机制,相互发挥作用以维持机体繁殖内分泌的稳定:1)中枢神经系统对下丘脑分泌激素的调节:大脑皮层对机体内外环境信息进行收集与分析,再对下丘脑发出指令,下丘脑随之决定相关激素的分泌量与释放量;2)GnRH对垂体前叶FSH、LH合成和分泌的调节;3)由上到下的正反馈调节与负反馈调节[29]。负反馈:乏情季节,血液中高浓度的雌激素(E2)会抑制GnRH、FSH和LH的分泌。正反馈:发情季节,高浓度的E2则会加强GnRH、FSH和LH的分泌。这3种激素的合成与释放是母畜排卵前LH-FSH形成峰值引起排卵的主要因素[30]。
Hafez[31]推测了母绵羊季节性繁殖的内分泌调节机制,如图1所示。在乏情季节或处于长日照下,长日照通过视神经激活绵羊下丘脑“LH抑制中枢”,使抑制中枢对E2负反馈作用的敏感性增强,E2负反馈作用可以使下丘脑分泌GnRH的能力减弱,因此垂体对FSH、LH的分泌量减少,最终使卵泡E2的分泌量减少,低浓度的E2抑制了排卵前的LH峰,最终抑制卵泡的成熟和排出,使机体处于持续乏情的状态;而在发情季节或处于短日照下,绵羊下丘脑“抑制中枢”没有被激活,对E2负反馈作用的敏感性也随之减弱,最终使母羊正常发情排卵。
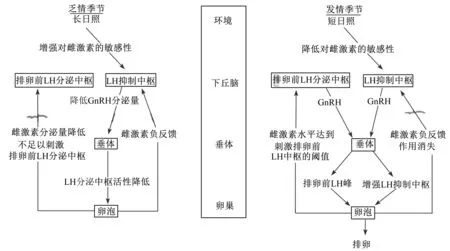
LH:促黄体素 lutropin;GnRH:促性腺激素释放激素 gonadotropin-releasing hormone。
图1母绵羊季节性繁殖的内分泌调节机制
Fig.1 Endocrine regulation mechanism of seasonal reproduction of female sheep[31]
光照可以调控绵羊繁殖内分泌,其中很大一部分原因是MLT在发挥作用[32]。在乏情期给母羊进行外源MLT注射可以使秋季开始发情的时间提前;另外,延长光照时间,减少MLT分泌则会起到相反的作用[33]。在繁殖季节,母羊血液中的LH、FSH浓度存在着明显的昼夜节律性变化,夜间LH浓度显著上升,公羊在繁殖季节里,LH浓度同样有类似的变化。饲养在光控棚中的公羊,在光照由亮变暗的情况下,血液中LH浓度会上升,这种情况在秋冬季节表现得更为突出。体格健康的母羊在日落一段时间之后,血液中LH浓度开始上升,另外,在14:00注射MLT,会发现LH的分泌在15:00左右开始上升。因此,MLT对发情期母羊LH的分泌有调控作用,进行外源MLT注射,血液中LH浓度开始上升的时间会早于自然状态下的母羊,由于LH的脉冲式分泌受到GnRH的调控,向发情期绵羊注射MLT可能引起GnRH的上升[34]。
此外,一些研究表明,绵羊繁殖季节开始于秋季,并不是因为秋季的光照周期缩短所致,而是因为绵羊对长光照周期的抑制作用失去了反应性[35]。同样,绵羊在晚冬停止繁殖活动,也并非对逐渐变长的光照周期产生了抑制作用,而是因为对短光照失去了反应性。由此看来,光照周期的作用应该是引导内源性繁殖节律在适当的时间开始或停止繁殖活动。Malpaux等[36]的研究指出,春夏季长光照所起的作用是协调动物的内源节律性,使绵羊在光照逐渐缩短的秋季开始繁殖。目前,国内外学者普遍认为,内源性节律与光照周期形成的动态匹配,是导致绵羊形成规律性的发情期与非发情期的主要原因[37]。
4 小 结
目前很多研究表明,光照周期作为主要的环境因子,协同MLT及其受体的内源性节律变化,可以影响反刍动物的生产与繁殖[38]。在光照周期调控生产与繁殖的过程中,PG分泌的MLT是整个神经内分泌调控系统的中枢。MLT通过调控下丘脑-垂体-性腺轴上相关激素的内分泌变化,影响反刍动物的生产性能与繁殖性能;MLT又受褪黑素受体的限制,只有在两者相互匹配时才能发挥其重要功能[39]。光照作为人工干预生物节律的手段,也会带来一些负面影响,例如动物经历长时间光照会使机体内分泌紊乱,造成生产性能下降、繁殖性能减弱等影响。当前,关于光照与褪黑素协同作用对动物生产性能和繁殖性能产生何种影响是繁殖生物学研究的一个热点。另外,表观遗传等先进技术手段在生理学领域的进一步应用,将有助于揭示褪黑素在调控动物生产、繁殖上的作用通路和分子机理。
[1] MALPAUX B,ROBINSON J E ,WAYNE N L.Regulation of the onset of the breeding season of the ewe,importance of long days and of an endogenous reproductive rhythm[J].Endocrinology,1989,122(1):269-278.
[2] 许鑫.光照控制对陕北白绒山羊绒毛生长的影响研究.硕士学位论文[D].杨凌.西北农林科技大学,2015:15.
[3] 王宏博,高雅琴.绒山羊皮肤毛囊结构及其与产绒量关系的研究进展[J].安徽农业科学,2008,36(29):12701-12703.
[4] 刘海英,张微,岳春旺,等.年份对内蒙古绒山羊体重和产绒性能的影响[J].草食家畜,2007(1):48-50.
[5] 李丰田.不同光照时间对辽宁绒山羊羊绒生长、增重和繁殖性能的影响[J].现代畜牧兽医,2012(8):53-55.
[6] 索朗达,益西多吉,吴玉江,等.非产绒期控制光照和埋植褪黑激素对藏西北绒山羊增绒效果的影响[J].繁殖与生理,2015,35(2):15-16.
[7] 郭礼祥.控制光照对陕北白绒山羊体重和绒毛长度的影响研究[J].家畜生态学报,2012,33(5):20-22.
[8] 王林枫.光照和埋植褪黑激素对内蒙古白绒山羊含氮物质分配和产绒性能的影响及调控的研究[D].博士学位论文,呼和浩特:内蒙古农业大学,2005.
[9] DICKS P,RUSSEL A J F,LINCOLN G A.The effect of melatonin-imlants from December until April,on plasma prolactin,triiodothy ronine and thyroxine concentrations and the timing of the spring moult in cashmere goats[J].Animal Science,1995,60:1239-247.
[10] WULIJI T,LITHERLAND A,GOETSCH A L,et al.Evaluation of melatonin and bromocryptine administration in Spanish goats:III.Effects on hair follicle activity,density and relationships between follicle characteristics[J].Small Ruminant Research,2006,66(1):11-21.
[11] 绒山羊增绒有了“光控技术”[N/OL].农民日报,2014-01-14(3).http://www.caaa.cn/show/newsartical/php? ID=328434.
[12] MITCHELL R J,BETTERIDGE,GURNSEY M P.Fibre growth cycles of cashmere-bearing,reproducing does in Southern Hawkes Bay,New Zealand,over a 30-month period[J].New Zealand Journal of Agricultural Research.1991,34(3):287-294.
[13] 张英来.光周期技术对奶牛的影响[J].饲料广角,2001(13):21.
[14] 范玲霞,李昊,梁云,等.光照时间对崂山奶山羊泌乳性能的影响[J].安徽农业科学,2015(15):161-163.
[15] DAHL G E,ELSASSER T H,CAPUCO A V,et al.Effects of a long daily photoperiod on milk yield and circulating concentrations of insulin-like growth factor-Ⅰ[J].Journal of Dairy Science,1997,80(11):2784-2789.
[16] 张忠诚.家畜繁殖学[M].北京:中国农业出版社,2007.
[17] DAHL G E,BUCHANAN B A,TUCKER H A.Photoperiodic effects on dairy cattle:a review[J].Journal of Dairy Science,2000,83(4):885-893.
[18] PHILLIPSAL C J C.The effect of supplementary light during winter on the growth,body composition and behaviour of steers and heifers[J].Animal Science,1997,65:173-181.
[19] AHARONI Y,BROSH A,EZRA E.Short communication:Prepartum photoperiod effect on milk yield and composition in dairy cows[J].Journal of Dairy Science,2000,83(12):2779-2781.
[20] LACASSES P,VINET C M,PETITCLERC D.Effect of prepartum photoperiod and melatonin feeding on milk production and prolactin concentration in dairy heifers and cows[J].Journal of Dairy Science,2014,97(6):3589-3598.
[21] 李云甫,杨和平.光照时间对奶牛泌乳和采食性能的影响[J].家畜生态学报,2009,30(4):37-39.
[22] 范玲霞,李昊,梁云等.光照时间对崂山奶山羊泌乳性能的影响.安徽农业科学[J].2015(15):161-163.
[23] 朱荣康.环境因素对奶牛繁殖性能的影响[J].黑龙江动物繁殖,2017,25(1):14-16.
[24] 李胜利,孙海洲,赵存发,等.控制光照对内蒙古白绒山羊母羊营养物质消化率的影响研究[J].家畜生态学报,2014(9):28-32.
[25] 郭礼祥,王永军,黄艳平,等.控制光照对陕北白绒山羊体重和绒毛长度的影响研究[J].家畜生态学报,2012,33(5):20-22.
[26] SMITH J D,DOUGLASS L W,COYNE J A,et al.Melatonin feeding that simulates a short day photoperiod (SDPP) suppresses circulating insult in like growth factor I (IGF-Ⅰ) in prepubertal heifers[J].Journal of Animal Science,1997,75(1):215.
[27] 张海容,李发弟.褪黑素埋植对非繁殖季节绵羊发情和血浆中激素的影响[J].实验研究,2005,26(4):23-25.
[28] LINCOLN G A,ALMEIDA O F,KLANDORF H.Hourly fluctuations in the blood levels of melatonin and prolactin,follicle-stimulating hormone,luteinizing hormone,testosterone,tri-iodothyronine,thyroxine and cortisolin rams under artificial photoperiods and the effects of cranialsym pathectomy[J].Endocrinol,1982,92(2):237-250.
[29] 张海容.褪黑素调控绵羊季节性发情的研究进展[J].河北农业科学,2008,12(12):44-46.
[30] 王元兴.动物繁殖学[M].南京:江苏科学技术出版社,1993.
[31] HAFEZ E S E.Reproduction in farm animals[M]//Reproduction in farm animals.[S.l.]:Lea & Febiger,1962.
[32] PELLETIER J,BODIN L,HANOCQ E.Association between expression of reproductive seasonality and alleles of the gene for Mella receptor in the ewe[J].Biology of Reproduction,2000,62:(4)1096-1101.
[33] 茆达干,杨利国,吴结革.褪黑素对动物生殖的作用及其调控[J].草食家畜,2001(3):5-8.
[34] VANECEK J,KLEIND C.Melatonin inhibition of GnRH-induced LH release from neonatal rat gonadotroph:involvement of Ca2+not cAMP[J].AJP:Endocrinology and Metabolism.1995,269(1):85-90.
[35] ROBINSON J E,KARSCH F J.Refractoriness to inductive day length terminates the breeding season of the suffolk ewe[J].Biology of Reproduction.1985,32(5):656-663.
[36] MALPAUX B,DAVEAU A,MAURICEMANDON F,et al.Evidence that melatonin acts in the premammillary hypothalamic area to control reproduction in the ewe:presence of binding sites and stimulation of luteinizing hormone secretion byinsitumicroimplant delivery.[J].Endocrinology,1998,139(4):1508-1516.
[37] BERLINGUER F,LEONI G G,SUCCU S A,et al.Exogenous melatonin positively influences follicular dynamics,oocyte developmental competence and blastocyst output in a goat model[J].Journal of Pineal Research,2009,46(4):383-391.
[38] DUBOCOVICH M L,MARKOWSKA M.Functional MT1 and MT2 melatonin receptors in mammals[J].Endocrine,2005,27(2):101-110.
[39] SOARES J M J,MASANA M I,ERSAHIN C O,et al.Functional melatonin receptors in rat ovaries at various stages of the estrous cycle[J].Journal of Pharmacology and Experimental Therapeutics.2003,306(2):694-702.
猜你喜欢
今日农业(2022年14期)2022-09-15
现代畜牧科技(2021年8期)2021-10-13
中国奶牛(2019年12期)2020-01-08
意林(儿童绘本)(2019年6期)2019-08-05
小学生必读(低年级版)(2019年12期)2019-04-20
幼儿画刊(2018年7期)2018-07-24
新农业(2016年23期)2016-08-16
新农业(2016年22期)2016-08-16
数学大王·低年级(2016年8期)2016-05-14
饲料博览(2016年3期)2016-04-05
