
SREBPs天然小分子抑制剂的药理学研究进展Δ
2018-06-14 08:40谢治深赵振彪张振强王辉河南中医药大学药学院郑州450046
中国药房 2018年10期
谢治深,赵振彪,张振强,王辉(河南中医药大学药学院,郑州450046)
肥胖症、高脂血症、脂肪肝等代谢性疾病的发病率逐年攀升,已经成为21世纪威胁人类健康的重要原因,其发生、发展与体内糖脂代谢的紊乱密切相关。机体脂质的稳态主要是通过调节外源性脂质的吸收、机体内源性脂质的合成、脂质的降解转化等3个方面的平衡来维持,当机体吸收、合成的脂质多于降解转化的脂质,则易诱发肥胖症、高脂血症、脂肪肝等疾病[1]。胆固醇是脂质的重要组成部分,在肝脏合成的内源性胆固醇占体内胆固醇总量的比例高达70%,因此抑制胆固醇的合成成为降脂药物研发的热点。固醇调节元件结合蛋白(SREBPs)几乎涉及了游离脂肪酸和胆固醇从头合成的全过程,在脂代谢的调节过程中有着举足轻重的作用[2]。本课题组前期研究发现,以壳聚糖将二甲双胍和SREBPs的小干扰RNA制成纳米粒传递至小鼠体内,能够提升二甲双胍的降糖作用[3];另有大量研究表明,SREBPs抑制剂能有效抑制游离脂肪酸和胆固醇的合成、促进棕色脂肪燃烧、缓解胰岛素抵抗,从而改善肥胖症、糖尿病、动脉粥样硬化等多种糖脂代谢紊乱性疾病[2],成为了降脂药物研发的新方向。由于天然小分子化合物因具有较好的生物相容性、丰富的结构多样性而成为药物先导化合物的主要来源。近年来,SREBPs天然小分子抑制剂的研究迅速发展。基于此,笔者以“SREBPs”“Inhibitor”“Compound”“Natural products”等为关键词,组合查询2012年1月-2017年10月在PubMed、Web of Science数据库中的相关文献。结果,共检索到文献82篇,其中有效文献52篇,通过总结酚酸类、生物碱类、萜类、黄酮类、挥发油类等SREBPs天然小分子抑制剂的药理学研究进展,以期为脂代谢紊乱性疾病治疗药物的研发提供参考。
1 SREBPs调节糖脂代谢的分子机制
SREBPs是位于细胞内质网上的胆固醇敏感性转录因子,是调节胆固醇和脂肪酸从头合成的关键蛋白家族,其亚型分为SREBP-1a、SREBP-1c和SREBP-2,其中SREBP-1主要调节游离脂肪酸的合成和胰岛素介导的糖代谢,SREBP-2主要调节胆固醇的合成[2]。SREBPs在内质网中合成并与SREBPs剪切激活蛋白(SCAP)特异性结合形成复合体后,被胰岛素诱导基因蛋白(INSIG)锚定于内质网中形成前体SREBPs(pre-SREBPs);pre-SREBPs在低胆固醇或胰岛素的刺激下,被转运至高尔基体中,通过S1P和S2P蛋白剪切形成成熟SREBPs(m-SREBPs);m-SREBPs被转运至细胞核中发挥转录调控作用,从而促进羟甲基戊二酰辅酶A还原酶(HMGCR)、脂肪酸合成酶(FASN)、乙酰-辅酶A羧化酶(ACC)、硬脂酰-辅酶A脱氢酶1(SCD-1)、低密度脂蛋白受体(LDLR)等脂质合成关键酶的基因表达[4-5]。在转录阶段,SREBPs受多种转录因子的调控:SREBPs本身会受到核内转录的调控;胰岛素信号通路和肝X受体(LXR)的激活都能够强烈上调SREBP-1c的表达[6-7];雷帕霉素靶蛋白C1(mTORC1)的激活也能促进SREBP-1的表达[8];此外,miRNA-33也能负向调控SREBP-1的表达[9]。在剪切阶段,SREBPs主要受到哺乳动物雷帕霉素靶蛋白-S6核糖体蛋白激酶(mTOR-S6K)通路的负向调控[6],而这一通路又受到蛋白激酶B(PKB,又称为AKT)和5′-磷酸腺苷依赖的蛋白激酶(AMPK)的调控[8]。m-SREBPs形成后,或被泛素化降解[10],或在输入蛋白β(Importinβ)等蛋白的作用下被转运至细胞核内,在雷帕霉素靶蛋白-脂素蛋白(mTOR-lipin)的调控下发挥转录活性[11]。SREBPs抑制剂通过抑制SREBPs的合成、剪切、转运全过程的不同环节,从而抑制其转录活性,继而起到抑制脂质合成关键酶表达、降低体内脂质水平的作用。
2 各类SREBPs天然小分子抑制剂的药理学研究进展
2.1 酚酸类
酚酸类化合物是一类含有酚环的有机酸,由于具有明显的抗氧化、抗炎等作用而备受关注,主要代表化合物为鼠尾草酸、3-咖啡酰-4-二氢咖啡酰奎宁酸、丁香酸等(结构见图1)。
鼠尾草酸广泛存在于鼠尾草、丹参、迷迭香中[12]。本课题组前期研究发现,迷迭香的主要有效成分鼠尾草酸能够显著改善西式饮食诱导的小鼠血脂紊乱;同时还发现,鼠尾草酸对胆固醇的吸收以及吸入氧气/呼出二氧化碳的比例并无显著影响,但体内外均能抑制SREBPs的活性和进入细胞核中的表达量,且不升高SREBP-1c的表达;进一步研究分子机制发现,鼠尾草酸降低三酰甘油和胆固醇含量的作用依赖于抑制SREBPs[13];采用放射性同位素示踪实验发现,鼠尾草酸能抑制胆固醇和游离脂肪酸的从头合成[14]。以上结果表明,鼠尾草酸主要是通过26S蛋白酶体降解了剪切成熟后的m-SREBPs从而发挥作用[14],其机制不同于经典的SREBPs抑制剂,这为寻找新型SREBPs抑制剂提供了新思路。
Pil Hwang Y等[15]通过小干扰RNA试验研究发现,3-咖啡酰-4-二氢咖啡酰奎宁酸对SREBPs的调节作用是依赖于肝脏激酶B1(LKB1)和沉默信息调节因子1(SIRT1)调节AMPK来实现的,从而能减少高糖诱导的HepG2细胞的脂质合成。
Ham JR等[16]研究发现,丁香酸可通过抑制肥胖模型小鼠的炎症相关因子及基因的表达、促进脂质的氧化分解、抑制脂肪酸合成基因SREBP-1c以及靶基因等多种途径,缓解高脂饮食引起的小鼠脂代谢紊乱。
2.2 萜类
萜类化合物广泛存在于植物体内,是目前最热门也是作用机制研究最深入的一类SREBPs天然小分子抑制剂,主要代表化合物为维生素D、白桦脂酸、白桦脂醇、穿心莲内酯等(结构见图2)。
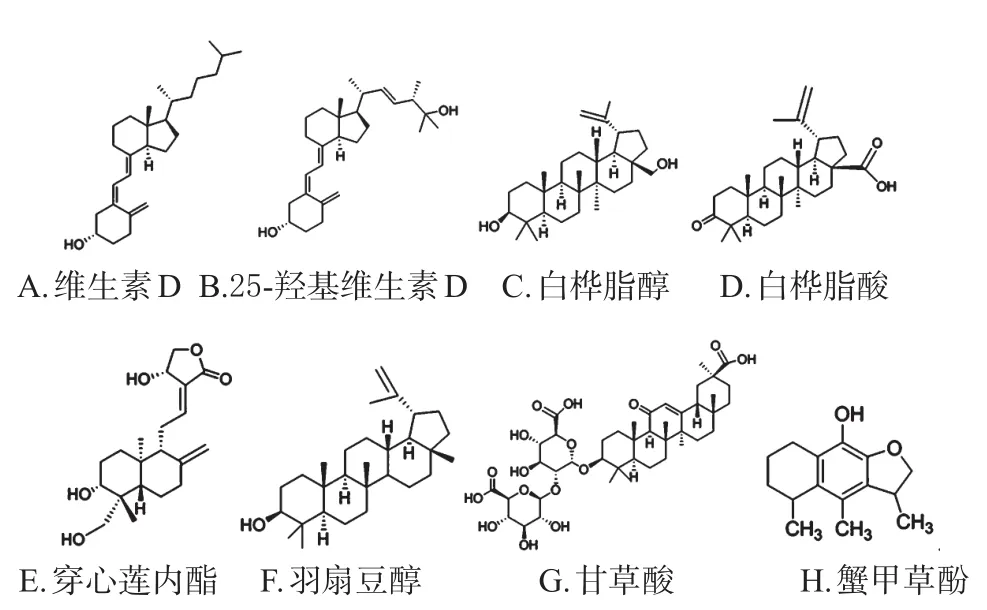
图2 萜类SREBPs小分子抑制剂代表化合物结构
Asano L等[17]研究发现,维生素D的体内代谢产物25-羟基维生素D能直接结合SCAP的C端,并促进其泛素化降解,从而抑制SREBPs的转运和剪切。这一方面解释了维生素D降脂的作用机制,同时也解释了膳食中缺乏维生素D以及体内缺乏维生素D羟化酶(细胞色素P450R1、7A1)容易诱发脂肪肝的原因。
Quan HY等[18]研究发现,白桦脂酸和白桦脂醇在体内外均能抑制SREBPs活性,改善糖脂代谢紊乱;当有STO-609(钙调素依赖蛋白激酶抑制剂)或Compound C(AMPK抑制剂)存在时,白桦脂酸未能表现出对SREBP-1的调节作用及降脂作用,提示白桦脂酸的降脂作用依赖于AMPK-mTOR-SREBPs途径。然而Tang JJ等[19]发现,白桦脂醇能够与SCAP结合从而抑制SREBPs被转运至高尔基体中剪切这一过程,从而改善高脂血症和动脉粥样硬化;当小鼠体内SCAP编码基因被沉默后,白桦脂醇亦未能表现出降脂作用。白桦脂醇与白桦脂酸的分子结构仅在于28位上的羟基和羧基,且对28位结构修饰后并没有发现活性差异,提示28位可能是非必需的药效基团[19]。
穿心莲内酯是穿心莲的主要有效成分,被誉为天然抗生素,在体内外均能抑制SREBPs并下调胆固醇和游离脂肪酸合成的相关靶基因,从而改善高脂诱导的小鼠肥胖症、高脂血症、脂肪肝等[20]。羽扇豆醇能通过多种途径抑制脂质合成,涉及IGR-1R/PI3K/AKT/SREBP-1通路[21]。甘草酸能下调SREBP-1,改善高脂或甲硫氨酸、胆碱缺乏引起的小鼠非酒精性脂肪肝[22]。此外,蟹甲草酚[23]、羊毛甾烷[24]均被发现具有抑制SREBPs的作用。
2.3 生物碱类
生物碱类化合物广泛存在于植物体内,是一类含氮的碱性有机化合物,是目前研究较多的一类SREBPs天然小分子抑制剂,主要代表化合物为小檗碱、咖啡因、槐果碱、罗希吐碱等(结构见图3)。
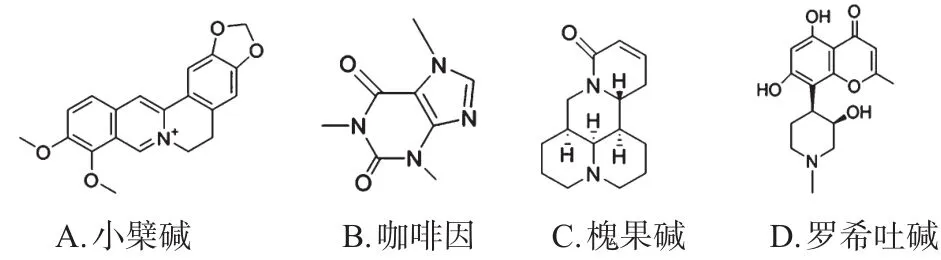
图3 生物碱类SREBPs小分子抑制剂代表化合物结构
近年来,小檗碱由于其不同于他汀类降脂药物的独特作用机制而备受学界关注。Jang J等[25]研究发现,小檗碱能够呈时间和浓度依赖性地激活AMPK,从而促进SREBP-1c的磷酸化并减少后者与靶DNA的结合,进而降低相关靶基因的表达量。此外,小檗碱可通过AMPKSREBPs途径逆转奥氮平引起的细胞内脂质增加[26]。Quan HY等[27]研究发现,咖啡因也能够呈剂量依赖性地激活AMPK从而下调FAS、ACC、HMGCR、LDLR等基因的表达,而AMPK的抑制剂Comound C能够逆转这一作用,表明咖啡因抑制SREBPs从而产生的降脂作用依赖于激活AMPK。此外,槐果碱也能通过激活AMPK调节脂质的合成,改善脂肪肝[28];罗希吐碱则通过激活过氧化物酶体增殖剂激活受体γ(PPARγ)和LXR,从而体内外抑制SREBP-1c和SREBP-2的表达,改善脂肪肝[29]。
2.4 黄酮类
黄酮类SREBPs天然小分子抑制剂的主要代表化合物有黄腐醇、异黄腐醇、柚皮素、8-异戊烯基柚皮素、木犀草素、芒柄花素等(结构见图4)。
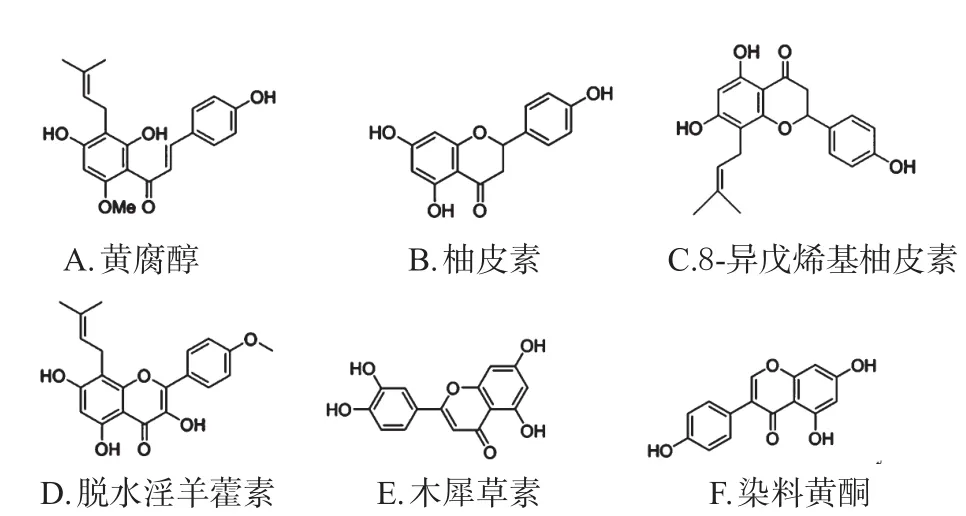
图4 黄酮类SREBPs小分子抑制剂代表化合物结构
Miyata S等[30]研究发现,黄腐醇能够直接结合在Sec23/24蛋白上,阻止SCAP-SREBP复合物与衣被蛋白Ⅱ(COPⅡ)蛋白的相互作用,继而阻断SCAP-SREBP复合物从内质网向高尔基体的转运和剪切,从而改善体内外脂代谢紊乱;同时还发现,结构相似的异黄腐醇、柚皮素、8-异戊烯基柚皮素也有相似的作用机制。Zheng ZG等[31]研究发现,脱水淫羊藿素及其类似物通过体内外调节LKB1/AMPK/mTOR途径,从而阻断SCAP-SREBPs复合物与Sec23/Sec24的相互作用,继而抑制SREBPs的剪切,发挥减少脂质合成的作用。
有研究发现,木犀草素能够抑制PI3K/AKT/mTOR/SREBPs通路从而起到降脂作用,并具有抗人绒毛膜癌细胞的作用[32]。异槲皮素[33]、染料木黄酮[34]能够激活AMPK,从而抑制SREBPs活性,减少脂质在细胞中堆积。
细胞成脂和成骨之间的分化平衡是肥胖和骨质疏松疾病进程之间的重要环节。Gautam J等[35]研究发现,芒柄花素能稳定β-联蛋白(β-catenin),激活AMPK以下调SREBPs的表达,激活PPARγ以减少三酰甘油在3T3-L1细胞中蓄积并抑制其成脂分化,从而改善高脂诱导的肥胖和骨流失。
2.5 皂苷类
皂苷类SREBPs小分子抑制剂的代表化合物有人参皂苷Re[36]、牛蒡子苷[37]、连翘苷[38](结构见图5),三者在体内外均能激活AMPK、抑制SREBP-1c,从而减少脂质的蓄积。人参皂苷Re对脂代谢和SREBP-1c的调控能够被AMPK的抑制剂Compound C所逆转,说明该调控作用依赖于AMPK激活。此外,獐牙菜苦苷[39]、虎杖苷[40]对SREBP-1c也有调节作用;采用免疫荧光法测定发现,虎杖苷能使SREBP-1蓄积在细胞质中,而使细胞核中的SREBP-1显著减少[41]。
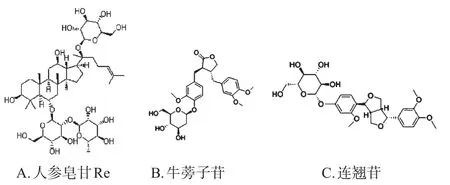
图5 皂苷类SREBPs小分子抑制剂代表化合物结构
2.6 其他类
其他类SREBPs小分子抑制剂主要代表化合物有姜黄素、厚朴酚、和厚朴酚、川芎嗪、白藜芦醇、褐藻糖、红曲等(结构见图6)。
图6 其他类SREBPs小分子抑制剂代表化合物结构
姜黄素以往以其突出的抗癌、抗氧化等药理活性备受关注,而近年来发现其还具有突出的降脂活性,如Ding L等[42]发现姜黄素能显著抑制SREBP-1c和SREBP-2的mRNA及蛋白质的表达,以及相应靶基因FAS、ACC、HMGCR、LDLR的表达,同时发现LXR的靶基因腺苷三磷酸结合盒蛋白G5与G8(ABCG5、ABCG8)并没有显著改变,且pre-SREBP-1的表达也并未显著变化,提示姜黄素能抑制SREBPs但不激活LXR,是效果优良的SREBPs抑制剂。
厚朴酚、和厚朴酚均为中药厚朴的药效成分,二者依赖于磷酸化LKB1激活AMPK,从而阻断LXR诱导的SREBP-1c升高,改善脂肪肝[43]。
川芎嗪是川芎的有效成分之一,是目前心脑血管疾病研究的热点化合物,其能够抑制SREBPs、改善载脂蛋白E基因敲除(ApoE-/-)模型小鼠的动脉粥样硬化、抑制磷酸化AKT(p-AKT)和磷酸化mTOR(p-mTOR)的表达,因此推测PI3K/AKT/mTORC1可能是其作用途径[44]。
Andrade JM等[45]发现白藜芦醇能抑制脂质合成,抑制SREBP-1c及其靶基因,从而改善脂肪肝;Tang LY等[46]发现白藜芦醇能通过激活AMPK-Lipin-1通路,抑制SREBP-1c,从而改善脂代谢紊乱;然而Yashiro T等[47]却发现白藜芦醇能通过激活SREBPs,从而上调LDLR的表达、促进肝细胞摄取代谢胆固醇,这一结论与其他研究截然相反。因此,白藜芦醇对SREBPs的作用还有待进一步深入探究。
此外,多糖类化合物褐藻糖在体内能够有效抑制SREBP-2及其靶基因,改善糖脂代谢紊乱[48]。红曲可以应用于他汀类药物耐受的患者,又能够避免他汀类的肝损伤、肌纤维溶解等副作用;从中分离得到的红曲霉素和红曲黄色素均能够激活PPARγ和AMPK,并能够抑制SREBP-1c,从而改善非酒精脂肪肝[49-50]。莶萜四醇在激活Wnt/β-catenin信号通路的同时,能够抑制SREBP-1c,从而抑制3T3-L1细胞的成脂分化[51]。
3 结语
SREBPs作为降脂的潜在靶点,因为涉及游离脂肪酸和胆固醇从头合成的全过程,因而比起其他众多单一靶点而言更具优势。SREBPs抑制剂作为潜在的调脂药,引起了医药研发人员的关注。然而目前经典的SREBPs抑制剂如25-羟胆固醇在抑制SREBPs的同时也激活了LXR,继而激活SREBP-1c基因,使游离脂肪酸合成基因表达增强、血浆中三酰甘油的含量升高,因此极大限制了其成药性[52]。综合本文论述,笔者发现近年来研究者发现了大量在体内外均能抑制SREBPs、有强降脂作用的天然小分子化合物,其中有些化合物在作用于SREBPs的同时不影响LXR,可作为调脂药研发的先导化合物,还有不少化合物在抑制胆固醇合成的同时能够降低血浆中三酰甘油的含量,极大地鼓舞了研究者的信心。有部分天然小分子化合物在激活AMPK的同时还能够促进葡萄糖的糖酵解过程,从而同时降低血脂和血糖,对改善糖脂代谢紊乱有积极意义,笔者建议对这部分化合物重点关注。此外,虽然部分已发现的天然小分子化合物对SREBPs的调控机制已经明晰,研究也较为深入,但还有很大一部分化合物是否依赖于AMPK调节SREBPs、如何通过AMPK调节SREBPs等作用机制问题尚未研究清楚,有待进一步深入探讨。
[1] REISS AB,VOLOSHYNA I,DE LEON J,et al.Cholesterol metabolism in CKD[J].Am J Kidney Dis,2015,66(6):1071-1082.
[2] XIAO X,SONG BL.SREBP:a novel therapeutic target[J].Acta Biochim Biophys Sin:Shanghai,2013,45(1):2-10.
[3] WANG FZ,XIE ZS,XING L,et al.Biocompatible polymeric nanocomplexes as an intracellular stimuli-sensitive prodrug for type-2 diabetes combination therapy[J].Biomaterials,2015,73:149-159.
[4] HORTON JD,GOLDSTEIN JL,Brown MS.SREBPs:activators of the complete program of cholesterol and fatty acid synthesis in the liver[J].J Clin Invest,2002,109(9):1125-1131.
[5] HORTON JD.Sterol regulatory element-binding proteins:transcriptional activators of lipid synthesis[J].Biochem Soc Trans,2002,30(Pt 6):1091-1095.
[6] OWEN JL,ZHANG Y,BAE SH,et al.Insulin stimulation of SREBP-1c processing in transgenic rat hepatocytes requires p70 S6-kinase[J].Proc Natl Acad Sci USA,2012,109(40):16184-16189.
[7] YOSHIKAWA T,SHIMANO H,AMEMIYA-KUDO M,et al.Identification of liver X receptor-retinoid X receptor as an activator of the sterol regulatory element-binding protein 1c gene promoter[J].Mol Cell Biol,2001,21(9):2991-3000.
[8] PORSTMANN T,SANTOS CR,GRIFFITHS B,et al.SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth[J].Cell Metab,2008,8(3):224-236.
[9] FERNANDEZ-HERNANDO C,SUÁREZ Y,RAYNER KJ,et al.MicroRNAs in lipid metabolism[J].Curr Opin Lipidol,2011,22(2):86-92.
[10] BENGOECHEA-ALONSO MT,ERICSSON J.A phosphorylation cascade controls the degradation of active SREBP1[J].J Biol Chem,2009,284(9):5885-5895.
[11] PETERSON TR,SENGUPTA SS,HARRIS TE,et al.mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway[J].Cell,2011,146(3):408-420.
[12] BEGUM A,SANDHYA S,SHAFFATH ALI S,et al.An in-depth review on the medicinal flora Rosmarinus officinalis(Lamiaceae)[J].Acta Sci Pol Technol Aliment,2013,12(1):61-73.
[13] XIE ZS,ZHONG LJ,WAN XM,et al.Petroleum ether sub-fraction of rosemary extract improves hyperlipidemia and insulin resistance by inhibiting SREBPs[J].Chin J Nat Med,2016,14(10):746-756.
[14] XIE ZS,WAN XM,ZHONG LJ,et al.Carnosic acid alleviates hyperlipidemia and insulin resistance by promoting the degradation of SREBPs via the 26S proteasome[J].J Funct Foods,2017,31:217-228.
[15] PIL HWANG Y,GYUN KIM H,CHOI JH,et al.3-Caffeoyl,4-dihydrocaffeoylquinic acid from Salicornia herbacea attenuates high glucose-induced hepatic lipogenesis in human HepG2 cells through activation of the liver kinase B1 and silent information regulator T1/AMPK-dependent pathway[J].Mol Nutr Food Res,2013,57(3):471-482.
[16] HAM JR,LEE HI,CHOI RY,et al.Anti-steatotic and antiinflammatory roles of syringic acid in high-fat diet-induced obese mice[J].Food Funct,2016,7(2):689-697.
[17] ASANO L,WATANABE M,RYODEN Y,et al.Vitamin D metabolite,25-hydroxyvitamin D,regulates lipid metabolism by inducing degradation of SREBP/SCAP[J].Cell Chem Biol,2017,24(2):207-217.
[18] QUAN HY,KIM DY,KIM SJ,et al.Betulinic acid alleviates non-alcoholic fatty liver by inhibiting SREBP1 activity via the AMPK-mTOR-SREBP signaling pathway[J].Biochem Pharmacol,2013,85(9):1330-1340.
[19] TANG JJ,LI JG,QI W,et al.Inhibition of SREBP by a small molecule,betulin,improves hyperlipidemia and insulin resistance and reduces atherosclerotic plaques[J].Cell Metab,2011,13(1):44-56.
[20] DING L,LI J,SONG B,et al.Andrographolide prevents high-fat diet-induced obesity in C57BL/6 mice by suppressing the SREBP pathway[J].J Pharmacol Exp Ther,2014,351(2):474-483.
[21] KWON HH,YOON JY,PARK SY,et al.Activity-guided purification identifies lupeol,a pentacyclic triterpene,as a therapeutic agent multiple pathogenic factors of acne[J].J Invest Dermatol,2015,135(6):1491-1500.
[22] WANG C,DUAN X,SUN X,et al.Protective effects of glycyrrhizic acid from edible botanical glycyrrhiza glabra against non-alcoholic steatohepatitis in mice[J].Food Funct,2016,7(9):3716-3723.
[23] LIU W,FURUTA E,SHINDO K,et al.Cacalol,a natural sesquiterpene,induces apoptosis in breast cancer cells by modulating AKT-SREBP-FAS signaling pathway[J].Breast Cancer Res Treat,2011,128(1):57-68.
[24] LEE I,KIM J,RYOO I,et al.Lanostane triterpenes from Ganoderma lucidum suppress the adipogenesis in 3T3-L1 cells through down-regulation of SREBP-1c[J].Bioorg Med Chem Lett,2010,20(18):5577-5581.
[25] JANG J,JUNG Y,SEO SJ,et al.Berberine activates AMPK to suppress proteolytic processing,nuclear translocation and target DNA binding of SREBP-1c in 3T3-L1 adipocytes[J].Mol Med Rep,2017,15(6):4139-4147.
[26] LI Y,ZHAO X,FENG X,et al.Berberine alleviates olanzapine-induced adipogenesis via the AMPK alpha-SREBP pathway in 3T3-L1 cells[J].Int J Mol Sci,2016,17(11):e1865.
[27] QUAN HY,KIM DY,CHUNG SH.Caffeine attenuates lipid accumulation via activation of AMP-activated protein kinase signaling pathway in HepG2 cells[J].BMB Rep,2013,46(4):207-212.
[28] SONG CY,SHI J,ZENG X,et al.Sophocarpine alleviates hepatocyte steatosis through activating AMPK signaling pathway[J].Toxicol in Vitro,2013,27(3):1065-1071.
[29] VARSHNEY S,SHANKAR K,BEG M,et al.Rohitukine inhibits in vitro adipogenesis arresting mitotic clonal expansion and improves dyslipidemia in vivo[J].J Lipid Res,2014,55(6):1019-1032.
[30] MIYATA S,INOUE J,SHIMIZU M,et al.Xanthohumol improves diet-induced obesity and fatty liver by suppressing sterol regulatory element-binding protein(SREBP)activation[J].J Biol Chem,2015,290(33):20565-20579.
[31] ZHENG ZG,ZHOU YP,ZHANG X,et al.Anhydroicaritin improves diet-induced obesity and hyperlipidemia and alleviates insulin resistance by suppressing SREBPs activation[J].Biochem Pharmacol,2016,122:42-61.
[32] LIM W,YANG C,BAZER FW,et al.Luteolin inhibits proliferation and induces apoptosis of human placental choriocarcinoma cells by blocking the PI3K/AKT pathway and regulating sterol regulatory element binding protein activity[J].Biol Reprod,2016,95(4):82.
[33] ZHOU J,YOSHITOMI H,LIU T,et al.Isoquercitrin activates the AMP-activated protein kinase(AMPK)signal pathway in rat H4ⅡE cells[J].BMC Complement Altern Med,2014,14:42.
[34] LIU H,ZHONG H,YIN Y,et al.Genistein has beneficial effects on hepatic steatosis in high fat-high sucrose diettreated rats[J].Biomed Pharmacother,2017,91:964-969.
[35] GAUTAM J,KHEDGIKAR V,KUSHWAHA P,et al.Formononetin,an isoflavone,activates AMP-activated protein kinase/beta-catenin signalling to inhibit adipogenesis and rescues C57BL/6 mice from high-fat diet-induced obesity and bone loss[J].Br J Nutr,2017,117(5):645-661.
[36] QUAN HY,YUAN HD,JUNG MS,et al.Ginsenoside Re lowers blood glucose and lipid levels via activation of AMP-activated protein kinase in HepG2 cells and high-fat diet fed mice[J].Int J Mol Med,2012,29(1):73-80.
[37] MIN B,LEE H,SONG JH,et al.Arctiin inhibits adipogenesis in 3T3-L1 cells and decreases adiposity and body weight in mice fed a high-fat diet[J].Nutr Res Pract,2014,8(6):655-661.
[38] DO MT,KIM HG,CHOI JH,et al.Phillyrin attenuates high glucose-induced lipid accumulation in human HepG2 hepatocytes through the activation of LKB1/AMP-activated protein kinase-dependent signalling[J].Food Chem,2013,136(2):415-425.
[39] PATEL TP,SONI S,PARIKH P,et al.Swertiamarin:an active lead from enicostemma littorale regulates hepatic and adipose tissue gene expression by targeting PPAR-γ and improves insulin sensitivity in experimental NIDDM rat model[J].Evid Based Complement Alternat Med,2013.DOI:10.1155/2013/358673.
[40] ZHANG J,TAN Y,YAO F,et al.Polydatin alleviates nonalcoholic fatty liver disease in rats by inhibiting the expression of TNF-α and SREBP-1c[J].Mol Med Rep,2012,6(4):815-820.
[41] HAO J,CHEN C,HUANG K,et al.Polydatin improves glucose and lipid metabolism in experimental diabetes through activating the AKT signaling pathway[J].Eur J Pharmacol,2014,745:152-165.
[42] DING L,LI J,SONG B,et al.Curcumin rescues high fat diet-induced obesity and insulin sensitivity in mice through regulating SREBP pathway[J].Toxicol Appl Pharmacol,2016,304:99-109.
[43] LEE JH,JUNG JY,JANG EJ,et al.Combination of honokiol and magnolol inhibits hepatic steatosis through AMPK-SREBP-1c pathway[J].Exp Biol Med:Maywood,2015,240(4):508-518.
[44] ZHANG Y,REN P,KANG Q,et al.Effect of tetramethylpyrazine on atherosclerosis and SCAP/SREBP-1c signaling pathway in ApoE-/-mice fed with a high-fat diet[J].EvidBasedComplement AlternatMed,2017.DOI:10.1155/2017/3121989.
[45] ANDRADE JM,PARAISO AF,DE OLIVEIRA MV,et al.Resveratrol attenuates hepatic steatosis in high-fat fed mice by decreasing lipogenesis and inflammation[J].Nutrition,2014,30(7/8):915-919.
[46] TANG LY,CHEN Y,RUI BB,et al.Resveratrol ameliorates lipid accumulation in HepG2 cells,associated with down-regulation of lipin1 expression[J].Can J Physiol Pharmacol,2015.DOI:10.1139/cjpp-2015-0125.
[47] YASHIRO T,NANMOKU M,SHIMIZU M,et al.Resveratrol increases the expression and activity of the low density lipoprotein receptor in hepatocytes by the proteolytic activation of the sterol regulatory element-binding proteins[J].Atherosclerosis,2012,220(2):369-374.
[48] SEO MS,KIM JH,KIM HJ,et al.Honokiol activates the LKB1-AMPK signaling pathway and attenuates the lipid accumulation in hepatocytes[J].Toxicol Appl Pharmacol,2015,284(2):113-124.
[49] HSU WH,CHEN TH,LEE BH,et al.Monascin and ankaflavin act as natural AMPK activators with PPARalpha agonist activity to down-regulate nonalcoholic steatohepatitis in high-fat diet-fed C57BL/6 mice[J].Food Chem Toxicol,2014,64:94-103.
[50] LEE BH,HSU WH,HUANG T,et al.Monascin improves diabetes and dyslipidemia by regulating PPARγ and inhibiting lipogenesis in fructose-rich diet-induced C57BL/6 mice[J].Food Funct,2013,4(6):950-959.
[51] KIM MB,SONG Y,KIM C,et al.Kirenol inhibits adipogenesis through activation of the Wnt/β-catenin signaling pathway in 3T3-L1 adipocytes[J].Biochem Biophys Res Commun,2014,445(2):433-438.
[52] LEHMANN JM,KLIEWER SA,MOORE LB,et al.Activation of the nuclear receptor LXR by oxysterols defines a new hormone response pathway[J].J Biol Chem,1997,272(6):3137-3140.
猜你喜欢
西北林学院学报(2022年3期)2022-06-09
世界科学技术-中医药现代化(2021年9期)2021-12-31
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
中成药(2018年12期)2018-12-29
中成药(2017年7期)2017-11-22
中成药(2017年10期)2017-11-16
中成药(2017年4期)2017-05-17
中成药(2017年3期)2017-05-17
华人时刊(2016年16期)2016-04-05
