
中南半岛前期异常气候条件对中国南方稻区褐飞虱灾变性迁入的影响及其预测模型
2018-06-07 02:58包云轩唐辟如孙思思陆明红谢晓金刘万才
生态学报 2018年8期
包云轩,唐辟如,孙思思,陆明红,谢晓金,刘万才
1 南京信息工程大学气象灾害预报和评估协同创新中心/南京信息工程大学,南京 210044 2 江苏省农业气象重点实验室/南京信息工程大学,南京 210044 3 农业部全国农业技术推广与服务中心,北京 100125
褐飞虱,Nilaparvatalugens(Stål),是亚洲各稻区的主要害虫之一,也是我国水稻生产上最为严重的害虫,其大面积发生给水稻生产带来了巨大的损失[1]。褐飞虱为喜温湿、耐寒极弱、无休眠性昆虫,由于冬季温度较低和水稻寄主的缺乏,褐飞虱只能在热带和亚热带南部地区生存[2- 3],国内仅能在华南南部、西南南部等部分南方地区越冬,国外主要在中南半岛越冬[4],其种群发生受气候影响明显,褐飞虱的越冬边界随着气候变暖呈现出北移的趋势,相应的越冬区也随之增大[5- 8]。春、夏两季自南向北迁飞,秋季则从北向南回迁。20世纪 80 年代以来,褐飞虱在我国年发生面积为1.33×107—2.0×107hm2,约占水稻种植面积的50%,年损失稻谷达1.0×109—1.5×109kg。2005—2010年,褐飞虱连续6a在我国南方稻区大发生,引起水稻大面积的虱烧和“冒穿”,造成了严重的损失。
近30多年来,全球气候的变化对农业生态系统产生了重大的影响,对农作物害虫而言,也不例外。为此,研究气候变化背景下害虫的发生规律,及时有效地控制其危害,对减轻农作物产量的损失,确保我国粮食安全至关重要。近30a以来,昆虫雷达遥感监测技术已被应用于研究褐飞虱的迁飞观测,并证实了褐飞虱在中国进行着长距离的季节性往返迁飞[9- 11],另一方面,也证明了褐飞虱的迁移与天气、气候等因素密切相关[12- 13]。褐飞虱广泛分布于中国、日本、朝鲜半岛、东南亚、太平洋岛屿和澳大利亚等国家和地区[4]。受全球气候变化和农业产业结构调整等因素的影响,褐飞虱的迁入和致灾性趋于复杂化,出现了持续性大发生的新特点,给我国水稻的生产带来了严重的威胁[14- 15]。
我国目前褐飞虱难以治理主要是因为对前期境外虫源地的种群动态变化、气候条件及其对后期迁飞过程、迁入量的影响了解和预测还不够准确,我国水稻主产区位于秦岭、淮河以南,每年的主要虫源来自中南半岛[15- 17],在国内很难对虫源的越冬进行有效的检测,也无法监测中南半岛春季虫源的迁出量,从而无法在国内实施全方位而准确的预测和综合治理。想要从根源上控制我国褐飞虱的危害,必须将研究区域扩展到国界以外,对境外虫源地的褐飞虱种群动态及其气候条件进行研究。由于前期境外虫源地种群动态信息无法准确而全面地获知,但天气、气候数据全球共享,因此,本文拟通过前期中国褐飞虱境外主要虫源地中南半岛异常气候条件的分析及其与后期中国南方水稻主产区褐飞虱发生等级之间定性、定量关系的研究,探明中南半岛前期异常气候条件对中国后期褐飞虱灾变性迁入的影响,构建以前期虫源地异常气候条件为关键预报因子的我国南方稻区褐飞虱迁入年发生等级的预测模型,为我国褐飞虱的发生机理研究和中长期测报业务提供理论和技术支持。
1 资料和方法
1.1 研究区概况
1.1.1 中国南方稻区
中国南方稻区主要分布在淮河以南,包括海南、广西、广东、云南、贵州、福建、浙江、江西、湖南、重庆、四川、湖北、安徽、江苏、上海等省、市、自治区,以热带季风气候和亚热带季风气候为主,研究区夏季炎热多雨,冬季温和少雨,多丘陵、山地,热量条件较好,降水丰沛,以水田为主,多种植水稻、玉米、油菜等作物,一年可种植二至三熟作物,属于世界上水稻产量最高的地区。但近40a来,这一地区水稻产量因受来自中南半岛的迁飞性害虫褐飞虱的严重危害而损失明显。并且进入21世纪以来,褐飞虱在中国南方稻区的发生有加重和扩张的趋势(如越冬边界和发生区北移、发生和为害的海拔高度升高等)。
1.1.2 中南半岛
中南半岛(2°—28°N,90°—110°E)包括越南、老挝、柬埔寨、缅甸、泰国五国以及马来西亚西部,属于典型的热带季风气候。由于柬埔寨和马来西亚西部与中国没有直接的虫源交换[17],所以本文不作研究。该研究区全年高温,最冷月平均气温在18℃以上,降水丰沛,年降水量大部分地区为1500—2000mm,且光热资源充足,多河流冲积平原,土壤肥沃,优越的气候条件和自然资源为水稻的种植提供了有利的生态环境,同时也利于多种稻作病虫害的发生,其中褐飞虱虫害的发生程度最为严重,并且春、夏季褐飞虱种群会随气流北迁到中国境内,对中国水稻的生产带来严重的影响。
1.2 数据来源及处理方法
1.2.1 虫情资料
基于资料的完整性和系统性考虑,本文收集了中国南方稻区20个植物保护站1980—2016年褐飞虱逐日灯诱资料,该资料由农业部全国农业技术推广服务中心测报处提供,站点分布见图1。结合中国农业部1995年颁发的稻飞虱单站发生程度等级标准[18],将中国南方稻区这20个站点褐飞虱发生危害程度分为5个等级,全区平均的褐飞虱年发生等级定义为:某年各等级发生站点数与相应等级的乘积除以总站点数,分别统计1980—2016年单站褐飞虱年发生等级、年内迁入总量的对数值、始见期、终见期,再通过加权平均计算出区域年褐飞虱发生等级等。
1.2.2 气象资料
中南半岛1979—2016年18个气象台站(图1)冬春季温度场、湿度场资料来源于美国国家环境预测中心(National Center for Environmental Prediction,NCEP)提供的水平分辨率为2.5°×2.5°的气象再分析资料。
1.2.3 气候分析
分别统计好18个站点37年冬季和春季的平均气温和平均相对湿度,根据陈正洪等的冷冬和暖冬等级标准[19],对其进行冷暖划分。
(1)单站冷暖冬划分标准:单站冬季(春季)平均气温距平ΔT≤-0.5℃,定义为单站冷冬(冷春);反之,ΔT≥0.5℃,定义为单站暖冬(暖春);若-0.5℃<ΔT<0.5℃,则定义为气温正常冬(春)。
(2)区域冷暖冬划分标准:区域内冷冬站数与该区域总站数的百分比定义为冷冬指数Icw (%),若Icw ≥50%,则为区域冷冬;同理,区域内暖冬站数与该区域总站数的百分比定义为暖冬指数Iww (%),若Iww≥50%,则为区域暖冬;若区域内冷冬指数Icw <50%且暖冬指数Iww<50%,则根据冷冬单站数、暖冬单站数和相应的Icw和Iww值对冷暖冬进行划分,若冷冬单站数多于暖冬单站数,则定义为区域正常偏冷冬,反之,暖冬单站数多于冷冬单站数,则定义为区域正常偏暖冬。利用同样的方法可统计出单站冷春、暖春和气温正常春,并划分出区域冷春、暖春和区域正常偏冷春、区域正常偏暖春。
依据相对湿度进行气候干湿划分:
(1)单站干湿冬划分标准:单站平均相对湿度距平ΔRH≤-3,定义为单站干冬,反之,ΔRH≥3,定义为单站湿冬;若-3<ΔRH<3,则定义为单站湿度正常冬;类似的可以定义出单站干春、单站湿春、单站湿度正常春。
(2)区域干湿冬划分标准:利用与气温相同的方法定义和划分出区域干冬、区域湿冬和区域正常偏干冬、区域正常偏湿冬及区域干春、区域湿春和区域正常偏干春、区域正常偏湿春。
为了便于预测,定义:温度正常冬季(春季)为2,冷冬(春)及正常偏冷冬(春)为3,暖冬(春)及正常偏暖冬(春)为1;相对湿度正常冬季(春季)为2,干冬(春)及正常偏干冬(春)为3,湿冬(春)及正常偏湿冬(春)为1。
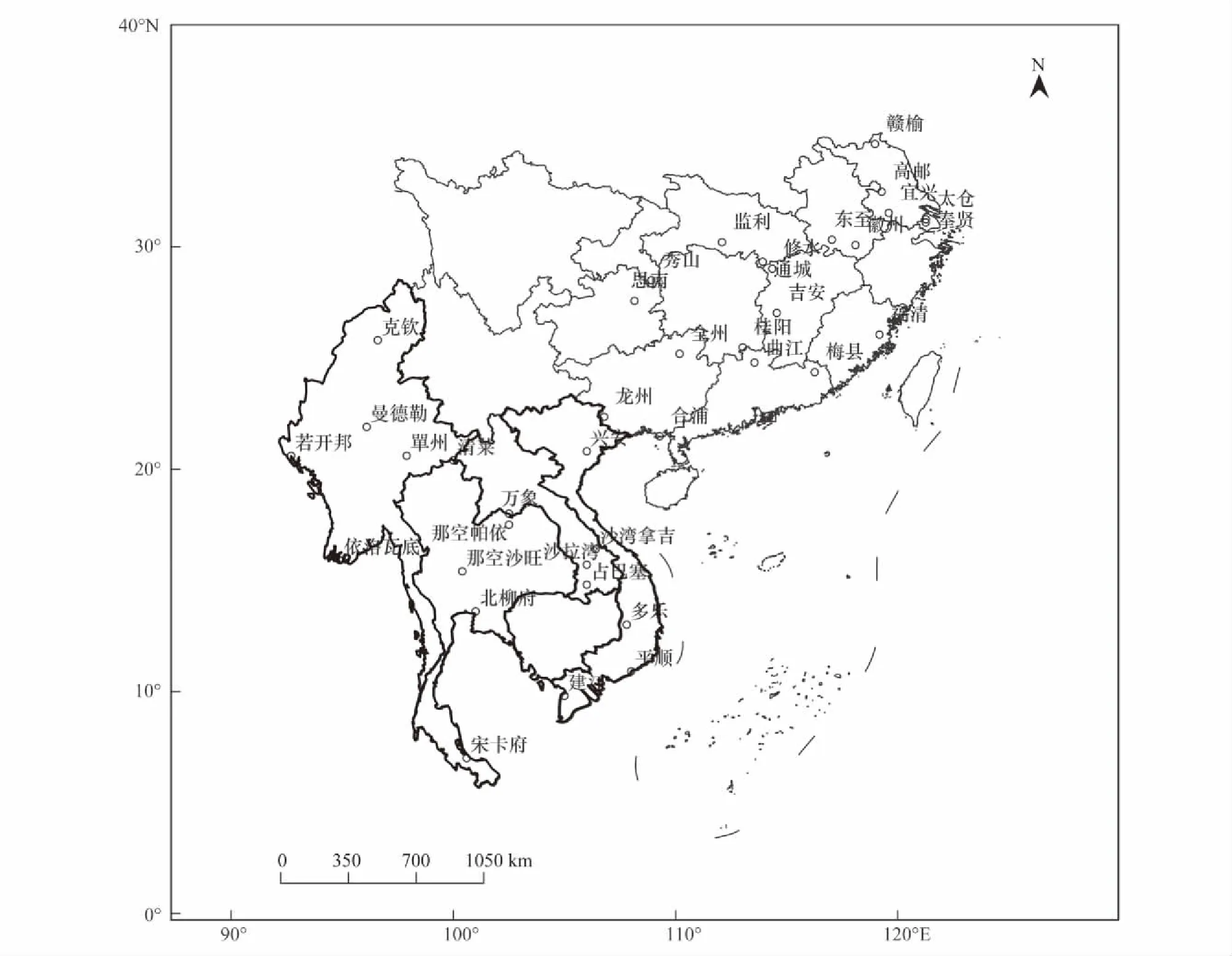
图1 中国南方稻区褐飞虱虫情监测站点分布和中南半岛气象站点分布Fig.1 Distribution of BPH′s monitoring stations in the main rice-growing region of the south China and the distribution of meteorological observational stations in the Indochina Peninsula
根据上述对区域气候冷暖、干湿的定义及相关数据处理的结果,本文分别对中南半岛前期异常温湿条件进行了下列时空分布特征的分析:
在温度异常的时间变化上,从表1中可以看出,20世纪80年代和90年代初春季和冬季温度较正常偏冷,21世纪初温度较正常偏暖,符合全球气温变暖的大背景。
在湿度异常的时间变化上,可以看出,20世纪80年代初期和末期、90年代末以及21世纪10年代前,冬季湿度较正常偏干,20世纪80年代中期、90年代初以及21世纪10年代后,冬季湿度较正常偏湿。20世纪末,春季湿度较正常偏干,21世纪初,春季湿度较正常偏湿。

表1 1980—2016年中南半岛的异常温度场和湿度场
冬季数据为前一年12月至当年2月的数据,春季数据为当年3—5月的数据;CW:冷冬,cold winter; WW:暖冬,warm winter; NCW:正常偏冷冬,normal partial cold winter; NWW:正常偏暖冬,normal partial warm winter; DW:干冬,dry winter; MW:湿冬,moist winter; NDW:正常偏干冬,normal partial dry winter; NMW:正常偏湿冬,normal partial moist winter; CS:冷春,cold spring; WS:暖春,warm spring; NCS:正常偏冷春,normal partial cold spring; NWS:正常偏暖春,normal partial warm spring; MS:湿春,moist spring; DS:干春,dry spring; NMS:正常偏湿春,normal partial moist spring; NDS:正常偏干春,normal partial dry spring
在空间变化上,图2反映了中南半岛1979—2016年单站温度场的空间分布情况,从图2可以看出,缅甸北部异常冬发生次数较高,37a里异常冬出现次数大于20次。而中南半岛南部异常冬出现次数大都在9次以下。还可以看出,缅甸地区异常春发生次数也较高,其中37a里异常春出现次数高于20次,而中南半岛南部地区异常春出现次数大多在5次以下。

图2 中南半岛1979—2016年气温异常冬、异常春发生次数的空间分布Fig.2 Spatial number distribution of abnormal winter and abnormal spring of temperature field in Indochina Peninsula during the period from 1979 to 2016
图3 反映了中南半岛1979—2016年单站湿度场的空间分布情况,从图3可以看出,缅甸、泰国、老挝和越南北部地区冬季湿度异常次数均较高,其中37a里湿冬出现次数在7次以上,干冬出现次数在6次以上,而中南半岛东南沿海地区异常冬出现次数均较低,越南的坚江和平顺等地区异常冬出现次数均低于2次。还可以看出,缅甸东北部、泰国北部湿春发生次数较高,37a里湿春出现次数大于10次。而中南半岛东南部地区湿春出现次数大都在5次以下。缅甸、泰国和老挝的交界处干春发生次数较高,37a干春出现次数在9次以上。而泰国、越南南部干春发生次数大多在4次以下。
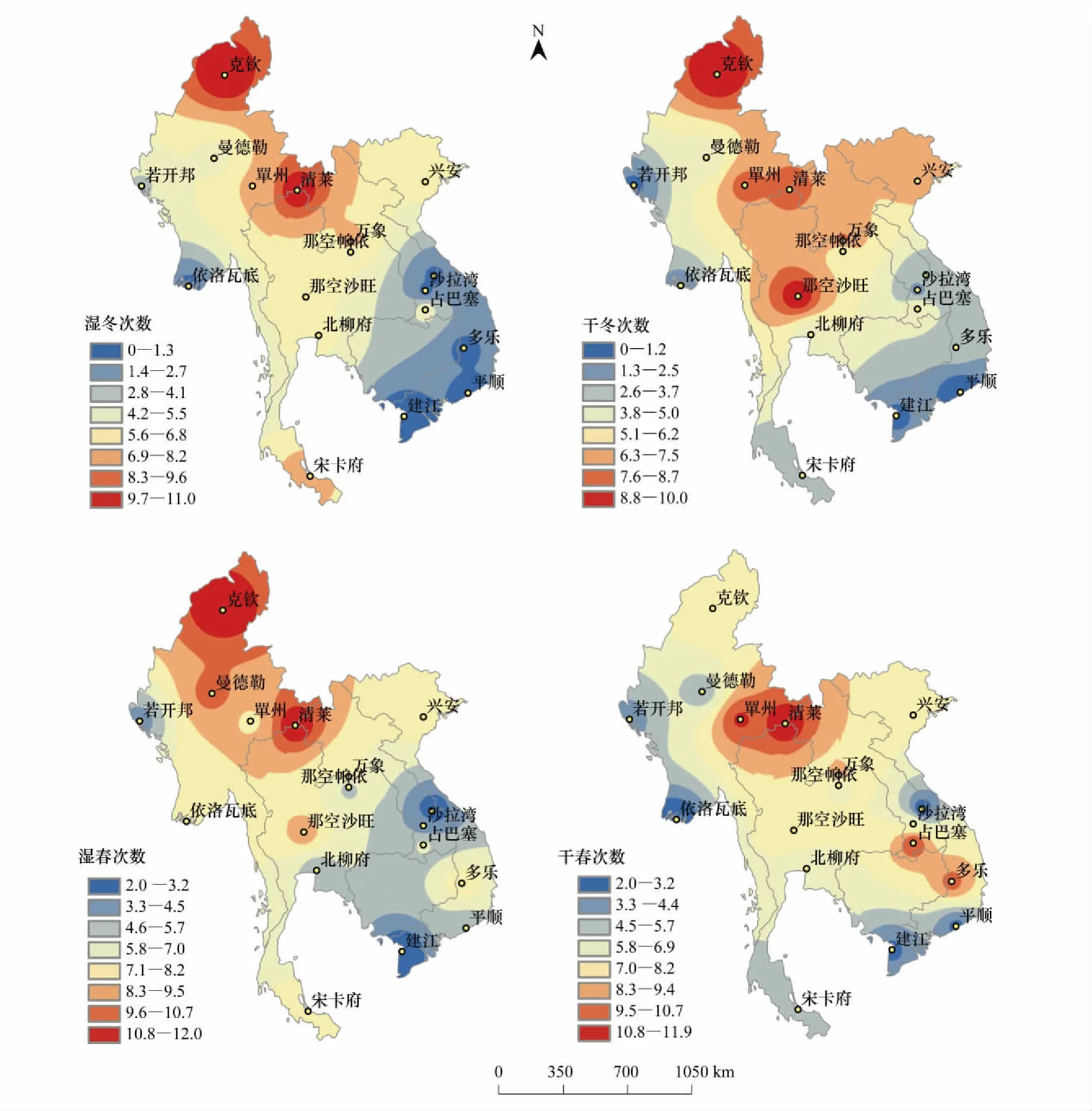
图3 中南半岛1979—2016年湿度场异常冬、异常春次数的空间分布Fig.3 Spatial distribution of abnormal winter number and abnormal spring number of relative humidity field in Indochina Peninsula during the period from 1979 to 2016
从中南半岛温、湿度空间分布图中可以看出,中南半岛37a中冬、春季的温度异常和冬、春季的湿度异常发生次数在空间上呈现出惊人的相似性,均呈现出北高南低的规律,并从北向南成环状递减。
1.2.3 基础地理信息
研究所用的基础图层为国家基础地理信息中心提供的1∶4000000世界电子地图。
1.2.4 模型的建立及效果分析
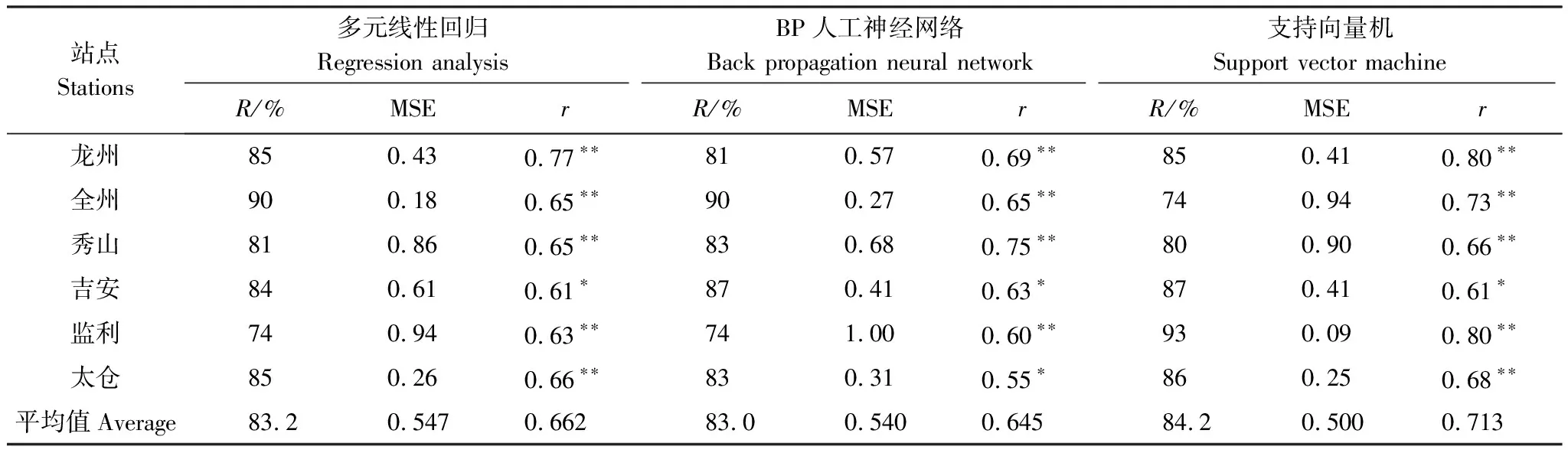
在考虑数据的完整性和站点空间分布的代表性等因素下,本研究在中国水稻主产区20个站点中选取龙州、全州、秀山、吉安、监利和太仓6个站作为预测的代表站点。将代表站点褐飞虱资料从始见期到终见期的逐日灯诱虫量(头数)进行累加,记为年内总迁入量(Y),再对其进行对数处理Lg(Y+1)。1980—2005年数据用于建模,2006—2016年数据用于预检。分别利用SVM、BP神经网络和逐步回归分析方法来进行预测,并比较3种方法的优劣。
(1) 预报因子的选取和多元线性回归预测模型的建立
通过SPSS软件对6个单站的年内褐飞虱迁入总量的对数值与前期中南半岛气象因子之间进行相关性分析,选取相关系数通过P<0.05显著性水平的因子作为预报因子,建立以下统计预报模型(表2)。
(2) BP人工神经网络模型
BP(Back Propagation)神经网络是一种通过误差逆传播算法来训练的多层前馈网络,可以学习和存贮大量的输入-输出模式映射关系,而无需事前揭示描述这种映射关系的数学方程。它的学习规则是使用梯度下降法,通过反向传播来不断调整网络的权值和阈值,具有较强的非线性处理的能力[20]。
由于每个站点的预报因子已经确定,使用反向传播算法对网络的权值和误差进行调整,从而使输出层的误差平方和小于初定的精度0.001,利用网络结构和初始条件,用1980—2005年的数据为训练样本,预测2006—2016年褐飞虱年内迁入总量的对数值,并通过历史回代和预报准确率来验证模型的准确性和稳定性。
(3) 支持向量机模型
支持向量机 (Support Vector Machine,简称SVM) 是一项功能强大的分类和回归的技术,可将模型的预测准确度最大化,善于处理小样本、高维数和非正态的数据[21- 22]。SVM的工作原理是将原始数据通过变换映射到高维的特征空间,这样针对那些不是线性可分的数据,也可以对该数据点进行分类。接着,使用变换后的新数据进行预测分类。
表2中国南方水稻主产区代表性站褐飞虱年迁入量对数的多元回归预测模型
Table2MultipleregressivepredictingmodelsofBPH′sannuallogarithmimmigrationamountattherepresentativestationsinthemainrice-growingregionsofthesouthChina

站点Stations模型ModelsFrP因子Factors龙州Y=8.635-0803×X1-0.04×X2-1.248×X310.3490.79∗∗0.000X1:平顺冷暖冬∗∗BinhThuancww∗∗X2:那空帕依冷暖冬∗NakhonPhanomcww∗X3:依洛瓦底干湿冬∗∗Ayeyarwadydmw∗∗全州Y=2.479+0.535×X110.3440.65∗∗0.004X1:依洛瓦底冷暖冬∗∗Ayeyarwadycww∗∗秀山Y=1.672+0.961×X115.9700.76∗∗0.000X1:克钦冷暖冬∗∗KachinStatecww∗∗吉安Y=4.546+0.35×X14.8370.61∗0.038X1:克钦干湿春∗KachinStatedms∗监利Y=1.041+1.1×X114.9510.71∗∗0.001X1:清莱干湿冬∗∗ChiangRaidmw∗∗太仓Y=4.539-0.323×X1-0.609×X25.2840.68∗∗0.003X1:克钦干湿冬∗KachinStatedmw∗X2:依洛瓦底冷暖春∗Ayeyarwadycws∗
** 表示通过P<0.01的显著性检验,* 表示通过P<0.05的显著性检验
在本研究中,采用模型class-rbf,内核设置为RBF,用1980—2005年的数据作为训练样本,对数据进行分类,再使用变换后的新数据进行预测分类,从而预测2006—2016年6个代表站点的褐飞虱年内迁飞总量的对数值,并将历年数据进行回代检验。
1.2.5 误差分析
选取平均平方误差(Mean squared error,MSE)和预测准确率(R)等指标来评价三种预测模型误差的大小及预报准确率。
(1)
(2)
式中,At:某一时段t的实际值;Ft:某一时段t的预测值;n:整个预测期内的时段个数(或预测总次数)。MSE表示预测值相对于实测值的离散程度,MSE越小,表示模型的稳定性能越好。R为预测准确度(%),R值越大,预测模型的效果越好。
2 结果与分析
2.1 中国南方水稻主产区褐飞虱发生情况
从表3中可以看出,1980—1983、1985、1987、1988、1991、1995、2005—2010年为偏重以上发生年,其余均为中等或偏轻以下发生年。
结合表3、图4可以看出,1980—2016年中国南方稻区20个站褐飞虱始见期在每年的3月到6月之间,1996年之前,始见期出现较迟,其中最晚的始见期出现在1996年5月16日,该年我国褐飞虱发生等级为偏轻发生,1996年到2014年始见期呈现出提前的趋势,其中最早的始见期出现在2010年的3月2日和2013年的3月2日,这两年我国褐飞虱发生程度分别为重发生和偏重发生。终见期出现在每年的9月到11月之间,1992年之前,终见期出现较早,其中最早的终见期出现在1992年9月26日,该年我国褐飞虱发生等级为轻发生,1992年到2014年终见期呈现出推迟的趋势,其中最晚的终见期出现在2010年11月30日,该年我国褐飞虱发生等级为重发生。
表31980—2016年中国南方稻区20个站点褐飞虱年发生等级及其区域平均
Table3BPH′soccurrencegradesof20stationsinmainrice-growingregionofChinaandtheaverageoccurrencegradesofthewholeregionduringtheperiodfrom1980to2016

年份Year1级Grade 12级Grade 23级Grade 34级Grade 45级Grade 5全区年发生等级Annual occurrence grade in the whole region(Yi)19802304113.95(4)19812311134.0(4)19821301154.35(5)19834210133.8(4)1984944032.2(3)19851313124.1(5)19861030162.5(3)19875002133.9(4)19884302113.65(4)1989922072.7(3)1990733162.8(3)19913212123.9(4)19921431021.65(2)1993822082.9(3)1994821272.9(3)1995334283.45(4)19961142032.0(2)1997761062.6(3)19981142032.0(2)19991431111.6(2)20001621012.4(3)2001745042.5(3)2002923062.6(3)2003843142.45(3)2004724072.9(3)20052000184.6(5)20060010194.9(5)20071131144.3(5)20081112154.45(5)2009722183.05(4)2010622283.2(4)20111130152.3(3)20121022062.5(3)2013833242.55(3)20141320142.05(3)20151332111.7(2)20161530011.3(2)
表3中“()”内数字为近似发生等级,当0.0 图4 1980—2016年中国南方稻区褐飞虱迁入始见期、终见期Fig.4 Beginning dates and ending dates of BPH′s immigration in the southern china from 1980 to 2016 20世纪90年代末至21世纪初期,褐飞虱始见期推迟、终见期提前,此时,我国褐飞虱发生等级多为轻发生和偏轻发生。20世纪80年代、21世纪初至今,褐飞虱始见期提前、终见期推迟,我国褐飞虱发生等级多为重发生和偏重发生。 图5为中国水稻主产区1980—2016年单站褐飞虱发生等级次数空间分布图,发生等级依次为一级、二级、三级、四级和五级。由图5可知,江苏一带褐飞虱发生程度为一级的次数最多,达到25次;贵州东北部二级发生次数最多,为4次;安徽西南部三级发生次数最多,为4次;广东、福建南部地区四级发生次数都较多,最多为10次;广西、湖南、贵州、江西西南部和安徽南部地区五级发生次数较多,最多为23次。37a里发生某等级的次数超过18次,则该区域定义为该等级发生较多,由此可见,褐飞虱在我国水稻主产区分布较广,发生程度都较大。 图5 中国南方稻区1980—2016年褐飞虱发生等级次数的空间分布Fig.5 Spatial distribution of the frequencies of BPH′s different occurrence grades in the main rice-growing region of Southern China during the period from 1980 to 2016 对褐飞虱的年内迁入总量的对数值与中南半岛冬、春季温湿条件进行相关分析,筛选出通过P<0.05显著性水平的气象因子,分别用SVM、BP人工神经网络和多元线性回归模型3种方法对我国南方水稻主产区代表站点的褐飞虱年迁入量进行模拟和预测。 2.2.1 3种方法历史回代准确率比较 表4为反映3种预测模型的历史回代检验情况,从表中可知,1980—2005年3种模型的平均历史拟合率都在80%以上,3种模型的预报效果都较好。但是,三者之间的模型稳定性和相关性还是存在明显的差异。支持向量机模型(SVM)的预测值与实际值的相关系数最大,其平均值为0.713,明显高于BP神经网络模型的0.645和多元线性回归模型的0.662,说明SVM模型预测值与实际值的变化趋势更为接近。SVM模型的平均均方误差为0.500,低于多元回归模型的0.547,也低于BP神经网络模型的0.540,显然 SVM模型的稳定性大于其他两种模型。由此可见,3种预测模型中, SVM模型更具有实际推广价值。 表41980—2005年6个代表站褐飞虱迁入量对数的3种预测模型拟合效果评价 Table4FittingeffectevaluationofBPH′sannualimmigrationamountbasedonthethreemodelsat6representativestationsduringtheperiodfrom1980to2005 站点Stations多元线性回归Regression analysisBP人工神经网络Back propagation neural network支持向量机Support vector machineR/%MSErR/%MSErR/%MSEr龙州850.430.77∗∗810.570.69∗∗850.410.80∗∗全州900.180.65∗∗900.270.65∗∗740.940.73∗∗秀山810.860.65∗∗830.680.75∗∗800.900.66∗∗吉安840.610.61∗870.410.63∗870.410.61∗监利740.940.63∗∗741.000.60∗∗930.090.80∗∗太仓850.260.66∗∗830.310.55∗860.250.68∗∗平均值Average83.20.5470.66283.00.5400.64584.20.5000.713 2.2.2 3种预测模型的预测准确性比较 以1980—2005年褐飞虱迁入量为训练样本,比较支持向量机(SVM)模型、BP神经网络和逐步回归模型预测2006—2016年褐飞虱迁入量的效果。将褐飞虱的预测值换算成褐飞虱的发生等级,与褐飞虱实际发生等级进行对比,预报结果以一个级别的差距认为是基本正确,3个模型预测准确率见表5。 由表5可知,3种模型的平均预测准确率(即预测等级一致或基本一致)在65%—80%之间,3种模型的预报效果较好。其中SVM模型的预测准确率为79%,明显高于BP神经网络的68%和逐步回归模型的67%,再比较预测等级与实测等级在二个级别以上的出现次数,发现逐步回归预报出现次数为16,高于BP神经网络模型的7次和SVM模型的11次,说明就精确性而言,SVM模型和BP神经网络优于逐步回归模型。由表4和表5可知,3种预测模型的比较结果为:SVM模型优于BP神经网络模型,也优于逐步回归模型。 表5 三种模型预测结果的比较 本文利用1980—2016年中国南方水稻主产区褐飞虱灯诱数据、1979—2016年中南半岛气象数据,从时间和空间上揭示了褐飞虱的迁入规律,分析了中南半岛前期温湿异常气候的时空特征,利用SVM、BP人工神经网络和多元线性回归方法对我国南方水稻主产区代表站点褐飞虱年迁入量进行了预测,在此基础上比较了3种模型的历史回代率和预测准确率,得到如下结论: (1)中南半岛气候异常区主要分布在北部一带,异常气候发生频率在中南半岛呈现出北高南低的规律,并从北向南呈环状递减。这可能与中南半岛独特的地形有关。中南半岛地势具有3个明显的特征,首先,中南半岛地势大致北高南低,多山地、高原,山川大致为南北走向,且山川相间排列,整个半岛地势犹如掌形。其次,其地势久经侵蚀而呈现出准平原状,喀斯特地形发育旺盛。第三,平原多分布在东南部沿海地区,主要是大河下游面积广大的冲积平原和三角洲。中南半岛上山脉、河流多由北向南延伸,形成了山河相间、纵列分布的特点。因此,在中南半岛的北部地势高,且多山川河流相间,地势复杂,一旦冷暖空气入侵不易排出,导致该地区异常天气出现次数增加。而中南半岛东南部地势平坦,靠近沿海地区,多平原和三角洲,冷暖空气畅通无阻,使得东南部沿海地区异常气候次数相对较少。有研究表明,海拔越高,气温变化率越明显,气候变化很大程度上取决于海拔的高低、地形的复杂程度以及所处的纬度等[23]。中南半岛北部地区多高山,海拔高、山川相间,地势复杂,南部(特别是东南沿海)地形比较平坦,因此,中南半岛北部异常气候发生频率明显高于东南沿海地区。 (2)进一步分析可知,1980年到1989年,中南半岛偏冷冬出现概率为90%,冷春出现概率为50%,则该段时期内中南半岛气温较往年都偏冷。偏湿冬出现概率为60%,偏干冬出现概率为30%,其余为正常冬;湿春出现概率为30%,其余为正常春,则该段时期中南半岛相对湿度较大,同时,我国褐飞虱发生程度为4级及以上的概率为70%,其余年份均为中等发生,大发生和偏重发生的概率明显高于中等发生的概率。调查资料发现,20世纪80年代,东南亚国家推广了高产、矮杆、耐肥的品种和多肥、密植的栽培技术以及滥用广谱性、杀生性化学农药,破坏了生态系统,削弱了自然因素的控制,从而改善了褐飞虱发生的生态条件,促使褐飞虱种群爆发成灾[4]。因此,尽管该段时间气温偏冷,冬季平均气温在20—21℃,温度不是褐飞虱生长发育的最适温度,但是能满足其种群生存的热量条件。由于外界人为因素的原因,导致褐飞虱种群密度急剧增大,并且相对湿度偏湿为种群提供了较适宜的湿度条件,在外界人为因素和湿度条件的共同作用下导致了该段时间内中国南方稻区褐飞虱始见期早、终见期晚、持续时间长、发生程度大。1990年到2005年,该段时间内中南半岛偏冷冬出现概率为44%,偏暖冬出现概率为56%,偏冷春出现概率为38%,偏暖春出现概率为62%,则该段时期中南半岛气温偏暖。而相对湿度表现为:偏湿冬和偏干冬出现概率同为50%,偏湿春出现概率为38%,偏干春出现概率为62%,则该段时间内中南半岛相对湿度较往年偏干,该段时间内,我国褐飞虱发生程度为4级及以上的概率仅为19%,大发生和偏重发生的概率明显低于中等及中等以下发生的概率。分析原因,虽然全球气候变暖给褐飞虱北迁和大发生提供了有利的热量条件,但是要造成褐飞虱在中国境内大发生,气象因素并不是唯一的影响因子,通过查阅资料,发现上世纪九十年代高效低毒的噻嗪酮和吡虫啉及综合防治技术在中国的研制与推广[24- 25],较好地控制了褐飞虱的北迁和南返,使得迁飞种群的密度和迁入峰次整体下降,从而从虫量上遏制了褐飞虱的大发生,此外,气候偏干也不利于褐飞虱的迁飞,因此,这段时期在自身和外部因素共同作用下褐飞虱在中国的发生程度偏轻。2006年到2014年,中南半岛偏冷冬出现概率为22%,偏暖冬出现概率为78%,偏冷春出现概率为33%,偏暖春出现概率为67%,则该段时间中南半岛气候偏暖。相对湿度表现为偏湿冬出现概率为44%,偏干冬出现概率为56%,偏湿春出现概率为67%,偏干春出现概率为33%;则该段时间相对湿度总的来说偏湿,该段时间内,褐飞虱在我国发生程度为5级的概率为56%,其余年份均为中等发生,大发生的概率略高于中等发生。在具备良好温湿条件的前提下,没有外界对褐飞虱种群的抑制作用,这段时期褐飞虱迁入始见期提前,终见期推迟,在我国发生时间长,发生程度大。通过上述分析可以发现:在没有外界人为因素严重干扰下,中南半岛前期温度偏暖(暖冬、暖春)、湿度偏湿(湿冬、湿春),易引起褐飞虱在中国南方稻区的偏重以上发生;而气候偏冷(冷冬、冷春)、偏干(干冬、干春),则易导致褐飞虱在中国南方稻区的偏轻以下发生。这与笔者以往研究的有关温场影响的结论相一致[26]。此外,前期中南半岛温湿气候条件的变化是由低纬大气环流变化所决定的,大气环流的异常会导致温湿气候条件的异常;而前期温湿气候条件的异常会反过来影响中南半岛夏季风的开始期,中南半岛夏季风开始是以印度洋上空低层西南季风向东北方向推进和来自苏门答腊群岛的热带对流向北增强和扩展为特征的[27],西南季风的发生早晚和强弱会显著地影响到中南半岛虫源地褐飞虱种群的迁出,继而影响到褐飞虱迁入中国的始见期、迁入量、降落区和分布范围[28]。 (3)通过比较3种模型的历史回代率和预测准确率,发现SVM模型的预测效果最好,BP神经网络次之,多元线性回归模型最差。多元线性回归的历史回代的准确率低于SVM模型、优于BP人工神经网络,但是预测准确率最低,不能从机理上很好的解释气象因子对褐飞虱迁入量影响的非线性关系,而其它两个模型则能更好的反映出气象因子与生物之间的关系[29- 30],因此预测效果较好。由于BP神经网络模型参数的选定和初始值的随机性对网络结构的不稳定有较大的影响[31],不利于推广使用,而SVM模型在结构风险最小化的基础上,提高了结论的规范化和精确度[30],更加适用于农业生产实际中的褐飞虱发生程度预测预报的推广。 参考文献(References): [1] 汤金仪, 胡伯海, 王建强. 我国水稻迁飞性害虫猖獗成因及其治理对策建议. 生态学报, 1996, 16(2): 167- 173. [2] 陈若篪, 程遐年, 杨联民, 殷向东. 褐飞虱卵巢发育及其与迁飞的关系. 昆虫学报, 1979, 22(3): 280- 288. [3] Kisimoto R, Sogawa K. Migration of the brown planthopper Nilaparvata lugens and the white-backed planthopper Sogatella furcifera in east Asia: the role of weather and climate // Drake V A, Gatehouse A G, eds. Insect Migration: Tracking Resources Through Space and Time. Cambridge: Cambridge University Press, 1995: 67- 92. [4] 张强翔, 任应党, 林克剑, 侯茂林, 鲁传涛, 刘红彦, 黄兴奇. 沿黄稻区灰飞虱越冬种群的时空分布及抽样技术研究. 应用昆虫学报, 2011, 48(3): 616- 621. [5] Otuka A, Zhou Y J, Lee G S, Matsumura M, Zhu Y Q, Park H H, Liu Z W, Sanada-Morimura S. Prediction of overseas migration of the small brown planthopper,Laodelphaxstriatellus(Hemiptera: Delphacidae) in East Asia. Applied Entomology & Zoology, 2012, 47(4): 379- 388. [6] Hu C X, Hou M L, Wei G S, Shi B K, Huang J L. Potential overwintering boundary and voltinism changes in the brown planthopper,Nilaparvatalugens, in China in response to global warming. Climatic Change, 2015, 132 (2): 337- 352. [7] 卢小凤, 霍治国, 申双和, 黄大鹏, 王丽, 肖晶晶, 于彩霞. 气候变暖对中国褐飞虱越冬北界的影响. 生态学杂志, 2012, 31(8): 1977- 1983. [8] 林光国. 闽中褐飞虱越冬情况的初步调查. 昆虫知识, 1986, 23(4): 150- 152. [9] 包云轩, 翟保平, 程遐年. 褐飞虱迁飞参数的数值模拟. 生态学报, 2005, 25(5): 1107- 1114. [10] Riley J R, Cheng X N, Zhang X X, Reynolds D R, Xu G M, Smith A D, Cheng J Y, Bao A D, Zhai B P. The long-distance migration ofNilaparvatalugens(Stål) (Delphacidae) in China: radar observations of mass return flight in the autumn. Ecological Entomology, 1991, 16 (4): 471- 489. [11] 程遐年, 陈若篪, 习学, 杨联民, 朱子龙, 吴进才, 钱仁贵, 杨金生. 稻褐飞虱迁飞规律的研究. 昆虫学报, 1979, 22(1): 1- 21. [12] 齐会会, 张云慧, 蒋春先, 武俊杰, 孙明阳, 杨秀丽, 程登发. 桂东北稻区第七代褐飞虱迁飞规律及虫源分析. 生态学报, 2014, 34(8): 2039- 2049. [13] 江广恒, 谈涵秋, 沈婉贞, 程遐年, 陈若篪. 褐飞虱远距离向北迁飞的气象条件. 昆虫学报, 1981, 24(3): 251- 261. [14] 程家安, 祝增荣. 2005年长江流域稻区褐飞虱暴发成灾原因分析. 植物保护, 2006, 32(4): 1- 4. [15] 齐国君, 芦芳, 胡高, 王凤英, 程遐年, 沈慧梅, 黄所生, 张孝羲, 翟保平. 2007年广西早稻田褐飞虱发生动态及虫源分析. 生态学报, 2010, 30(2): 462- 472. [16] 沈慧梅, 吕建平, 周金玉, 张孝羲, 程遐年, 翟保平. 2009年云南省白背飞虱早期迁入种群的虫源地范围与降落机制. 生态学报, 2011, 31(15): 4350- 4364. [17] 中日稻飞虱越冬考察组. 云南稻飞虱冬季虫源考察. 云南农业科技, 2000, (4): 3- 6. [18] 国家技术监督局. GB/T 15794—1995稻飞虱测报调查规范. 北京: 中国标准出版社, 1996. [19] 陈正洪, 马德栗. 湖北省1961—2008年冷冬时空变化特征 // 中国气象局国家气候中心暨气候研究开放实验室2010年度学术年会论文集. 北京: 国家气候中心,中国气象局气候研究开放实验室, 2011. [20] 李祚泳, 彭荔红. 基于人工神经网络的农业病虫害预测模型及其效果检验. 生态学报, 1999, 19(5): 759- 762. [21] Cristianini N, Shawe-Taylor J. Support vector and Kernel methods // Berthold M, Hand D J, eds. Intelligent Data Analysis. Berlin Heidelberg: Springer, 2007: 169- 197. [22] 张学工. 关于统计学习理论与支持向量机. 自动化学报, 2000, 26(1): 32- 42. [23] Shao J A, Li Y B, Ni J P. The characteristics of temperature variability with terrain, latitude and longitude in Sichuan- Chongqing Region. Journal of Geographical Sciences, 2012, 22(2): 223- 244. [24] 田茂仁, 蒲正国, 谢雪梅, 谯青春. 扑虱灵控制稻飞虱的效果评价.昆虫知识, 1996, 33(3): 129- 132. [25] 顾正远, 刘贤金, 韩丽娟, 肖英方. 吡虫啉的杀虫机制及配套应用技术研究. 西南大学学报, 1998, 20(5): 397- 400. [26] 包云轩, 薛周华, 刘垚, 蒋蓉, 谢晓金, 杨荣明, 朱凤. 江苏省褐飞虱迁入量的中长期预测模型. 中国农业气象, 2016, 37(1): 98- 110. [27] Widrow B, Lehr M A. 30 years of adaptive neural networks: perceptron, Madaline, and backpropagation. Proceedings of the IEEE, 1990, 78(9): 1415- 1442. [28] 包云轩, 黄金颖, 谢晓金, 陆明红. 季风进退和转换对中国褐飞虱迁飞的影响. 生态学报, 2013, 33(16): 4864- 4877. [29] Rumelhart D E, Hinton G E, Williams R J. Learning representations by back-propagating errors. Nature, 1986, 323(6088): 533- 536. [30] 陈永义, 俞小鼎, 高学浩, 冯汉中. 处理非线性分类和回归问题的一种新方法(I)——支持向量机方法简介. 应用气象学报, 2004, 15(3): 345- 354. [31] 鲁娟娟, 陈红. BP神经网络的研究进展. 控制工程, 2006, 13(5): 449- 451, 456- 456.

2.2 支持向量机(SVM)、BP神经网络和逐步回归分析3种预测模型的准确性比较

3 结论与讨论
猜你喜欢
——2022 F1意大利大奖赛
世界汽车(2022年5期)2022-06-10
加油站服务指南(2021年7期)2021-10-14
雷达与对抗(2021年4期)2021-03-18
生态学报(2020年20期)2020-11-13
——稻(二)
种业导刊(2019年3期)2019-05-21
智族GQ(2017年1期)2017-09-13
Coco薇(2015年12期)2015-12-10
Coco薇(2015年1期)2015-08-13
小说月刊(2015年3期)2015-04-19
种业导刊(2015年3期)2015-01-22
