
北京山区河岸带植物群落种-面积关系
2018-06-07 02:56孔庆仙信忠保夏晓平
生态学报 2018年8期
孔庆仙,信忠保,2,*,夏晓平
1 北京林业大学水土保持学院,北京 100083 2 北京林业大学,北京市水土保持工程技术研究中心,北京 100083
植物群落最小面积也称群落的临界抽样面积,是在一个最小地段内,对一个特定群落类型能提供足够的环境空间(环境和生物的特性),或者能保证展现出该群落类型的种类组成和结构的真实特征所需要的面积[1]。群落最小面积可以有效反映植物的固有特性和一定尺度下的相对均匀性[2]。由于最小面积很大程度上决定于空间异质性[3- 4],故研究植物群落首先要确定不同类型植物群落的最小面积。最小面积的确定常采用种-面积曲线法、重要值-面积曲线法和群落系数曲线法,但是在以往的研究中对参数的拟合采用哪种函数并不统一[5- 9]。种-面积曲线法是确定植物群落最小面积最常用的方法,可以基于栖息地减少和丧失引起的物种减少来预测物种灭绝率,设计最优保护区大小,评估人类活动对多样性的影响,对于保护生物多样性意义重大[7,9-11]。
河岸带是指高低水位之间的河床以及被抬高的水位、极端洪水和土壤持水能力对植物影响完全消失的区域[12- 13]。作为陆地生态系统和水生生态系统的过渡区域,边缘效应明显,植 物多样性较高[14- 16]。河流河岸带尤其是山区河流河岸带从上游到下游和从河床至陆上泛滥平原的植物群落、土壤理化性质和水分特征不同,形成不同类型的植物群落,对于维持河流河岸带稳定、减少面源污染、为动植物提供栖息地、为人类提供旅游观光资源作用重大[12,17- 23]。近年来,人类耕作、污水排放、河流开发等人为活动对河流河岸带资源的过渡或不适当开发利用[24- 26],使河岸带破碎化,植物多样性降低,严重影响了河流河岸带功能的发挥。对河流河岸带进行生态修复,保护河流河岸带动植物资源成为亟待解决的问题之一。
对不同立地条件下河流河岸带进行生态修复必须要明确此立地条件下的植物多样性。植物多样性的调查大多是根据经验值确定调查样地面积,比如常用的热带森林的调查样方面积为40m×40m,亚热带森林为20m×20m,温带森林为10m×10m,灌丛为2m×2m,草本植物为1m×1m[27],经验值使用不当会浪费人力物力资源,还可能导致实验结果的不准确性。近年来,对最小面积的研究越来越多,但主要针对森林内部植物群落[2,9,28- 33];由于人力物力资源的限制,对河流河岸带最小面积的研究只能依赖于较少的河流河岸带断面[6,34],对河流河岸带植物调查最小面积的认识较少[8]。在河岸带地区运用种面积关系推求最小样地面积可以拓展种面积关系研究的区域类型,同时河岸带的复杂性会促进种面积关系的发展。
本文以北京山区河流怀九河河岸带为研究对象,在全河段踏查的基础上运用聚类分析的方法将河岸带进行分类,对不同类型河岸带植物群落的最小面积进行研究,揭示不同类型河岸带植物群落最小面积的变化规律以及四种种-面积关系曲线模型的适用性,有利于确保河岸带植物群落调查的科学性,对山区河流河岸带植物群落调查具有借鉴意义,促进河流河岸带的保护和修复工作。
1 研究区概况及研究方法
1.1 研究区概况
怀九河(40°21′—40°30′N,116°16′—116°21′E)是北京典型的山溪性河流,自然性、经济性和社会性均具有较高的代表性,怀九河河岸带是重要城镇建设和乡村旅游发展经济带,河岸带生态修复既有基础又有影响。怀九河是发源于怀柔区黄花城东宫,经九渡河、四渡河,于前辛庄入怀柔水库的常年山溪性河流之一,全长68.9km,流域面积347.2km2。本区属于典型的大陆性季风气候,年降水量667.2mm,冬夏温差较大,1月平均气温为-5℃,7月平均气温为25.3℃,年最高气温38℃,最低气温-18℃[35]。土壤类型主要有普通褐土、褐土、潮褐土、褐潮土及粗骨性褐土等类型。植被主要有乔木加杨(Populuscanadensis)、旱柳(Salixmatsudana)、山核桃(Caryacathayensis)、板栗(Castaneamollissima),灌木荆条(Vitexnegundo)、胡枝子(Lespedezabicolor),以及占据草本优势种的菊科、禾本科和豆科植物鬼针草(Bidenspilosa)、牛尾蒿(Artemisiadubia)、狗尾草(Setairaviridis)、野大豆(Glycinesoja)等(图1)。

图1 北京市怀九河河岸带空间位置和流域图Fig.1 Spatial position and basin in Beijing Huaijiu river riparian
1.2 样地设置和调查内容
沿北京市怀柔区怀九河河岸带每隔大约1km设置一个长80m,宽度视河岸带宽度((5.81±3.55)m)而定的样地,如遇特殊样地则略过或加测一个样地的方法,共调查50个样地,从上游至下游样地分别标记为样地1、样地2、……、样地50。将每一样地划分为8块长10m的连续样方,在长10m的样方内沿对角线(上、中、下)方向设置3个1m×1m的小样方,调查结果代表10m样方内草本植物种类及数量,再将长10m,宽度根据立地条件决定的样方进行合并,将样方合并成长分别为10、20、30、40、50、60、70、80m,宽根据立地条件决定的样方共8个面积梯度。
记录样地的经纬度、海拔、坡度、坡向、坡长、岸坡类型(根据人类干扰程度赋值,岸坡改造程度越大得分越高)等立地因子,记录河岸带砾石盖度、硬化面积比例和现有修复措施(砾石盖度和硬化面积比例是指样地内砾石覆盖面积和硬化面积与样地面积的比值)。记录样地内所有乔灌木的物种名称、物种数量、胸径(DBH≥1.0cm)、高度、郁闭度和从幅,记录1m×1m样地内草本的物种名称、物种数量、总体盖度和平均高度。
1.3 研究方法
在种-面积关系的研究中,普遍采用饱和面积曲线拟合的方法,通过这种方法可以估计出群落的总物种数,这样在计算临界抽样面积时就不需要事先知道群落总物种数[33,36],从而减少外业调查的总工作量。本文选取4种常用的饱和曲线模型来拟合各类型河岸带种-面积关系[37]:
S=aA/(1+bA)
(1)
S=c/(1+ae-bA)
(2)
S=c-ae-bA
(3)
S=a(1-e-bA)
(4)
式中:A为面积,S为A中出现的物种数,a、b、c为待估参数。方程的拟合用SPSS18.0软件完成。对应于以上4种饱和曲线,要得到群落总种数一定比例ρ(0 <ρ< 1)的物种所需要的最小面积分别为:
A=ρ/(b(1-ρ))
(5)
A=-Ln(1-ρ)/aρ/b
(6)
A=-Ln(c(1-ρ)/a)/b
(7)
A=-Ln(1-ρ)/b
(8)
种-面积模型拟合优度的评价采用赤池信息量准则(Akaike Information Criterion,AIC)评价4种不同模型的拟合优度[38],公式如下:AIC=-2L+2K,其中,L为模型对数似然值,K为模型的参数个数,AIC数值越小,说明模型拟合得越好,预测值也就越准确。
本文中聚类分析和赤池信息量准则AIC计算使用R3.2.2完成,主成分分析用SPSS 18.0软件完成,其他图和表均使用Excel 2003完成。
2 研究结果
2.1 河岸带植被群落结构组成
根据50个样地的调查结果统计,北京市怀柔区怀九河河岸带物种数为255种,隶属于70科185属。其中菊科植物种类最多,为50种,占总数的19.61%;豆科24种,占总数的9.41%;禾本科23种,占总数的9.02%(图2)。
按照《北京植物志》[39]和《中国植物志》[40]的描述进行生活型分类,50个调查样地内乔木植物28种(10.98%),主要有加杨、旱柳和桑树(Morusalba);灌木24种(9.41%),主要有胡枝子、荆条、酸枣(Ziziphusjujuba)和孩儿拳头(Grewiabiloba);藤本植物6种(2.35%),主要有萝藦(Metaplexisjaponica)、蝙蝠葛(Menispermumdauricum)、葎叶蛇葡萄(Ampelopsishumulifolia)和五叶地锦(Parthenocissusquinquefolia);一年生草本植物83种(32.55%),主要有鬼针草、狗尾草、马唐(Digitariasanguinalis)和葎草(Humulusscandens);多年生草本植物91种(35.69%),主要有牛尾蒿、茜草(Rubiacordifolia)、茵陈蒿(Artemisiacapillaries)和早开堇菜(Violaprionantha)(图3)。
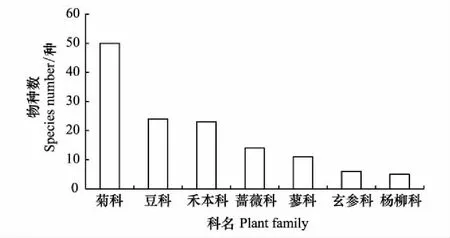
图2 主要科名的物种数变化Fig.2 Change in species number of plant family

图3 植物生活型物种数变化Fig.3 Change in species number of plant life form
河流河岸带上游植物种类(47.11±2.68)与数量(914.88±133.31)较多,中游和下游河段由于旅游业及农业的发展,植物种类(32.29±5.13,31.94±3.58)和数量(640.89±126.06,527.20±71.80)相对较少,但是随着河岸带修复越来越得到重视,中下游某些河段河岸带植物种类和数量变化明显(图4、图5)。

图4 河岸带群落植物种类统计Fig.4 Riparian community plant species statistics

图5 河岸带群落植物数量统计Fig.5 Riparian community plant number statistics
2.2 怀九河河岸带类型划分
将所调查的14个环境因子,用主成分分析法(PCA)进行主成分提取,方差最大化正交旋转后(P=0.05),提取了4个主成分,贡献率分别为28.49%、26.86%、11.70%和8.89%。第一主成分主要反映了岸坡类型和硬化面积比例的影响,其因子载荷系数分别为0.91和0.87,第二主成分主要反映了土壤容重和土壤有机碳的影响,其因子载荷系数分别为0.86和0.85,第三主成分主要反映了距入库口距离和海拔的影响,其因子载荷系数分别为0.90和0.90,第四主成分主要反映了岸坡宽度的影响,其因子载荷系数分别为0.87(表1)。
根据主成分分析结果,选取代表岸坡类型、岸坡宽度、现有修复措施类型、海拔、砾石盖度、硬化面积比例、土壤容重和土壤有机碳8项环境因子进行聚类分析,结果表明,在阈值为7.86时,将50个样地分为6类,包含了所有的干砌石河岸带样地5个,包含了所有的纯浆砌石河岸带样地7个,砾石覆盖度和海拔高的河岸带样地9个,防洪功能和景观功能主导的河岸带样地6个,岸坡宽度较大,坡顶栽植行道树的河岸带样地9个,依然保持原有岸坡立地条件,但是遭受人类干扰的河岸带样地14个(图6)。根据河岸带岸坡类型、现有修复措施、植物层次将6种类型河岸带依次命名为人工岸坡干砌石疏乔灌草河岸带、人工岸坡浆砌石河岸带、自然河岸带、人工岸坡观赏性乔灌草河岸带、人工岸坡乔灌草河岸带和近自然河岸带。
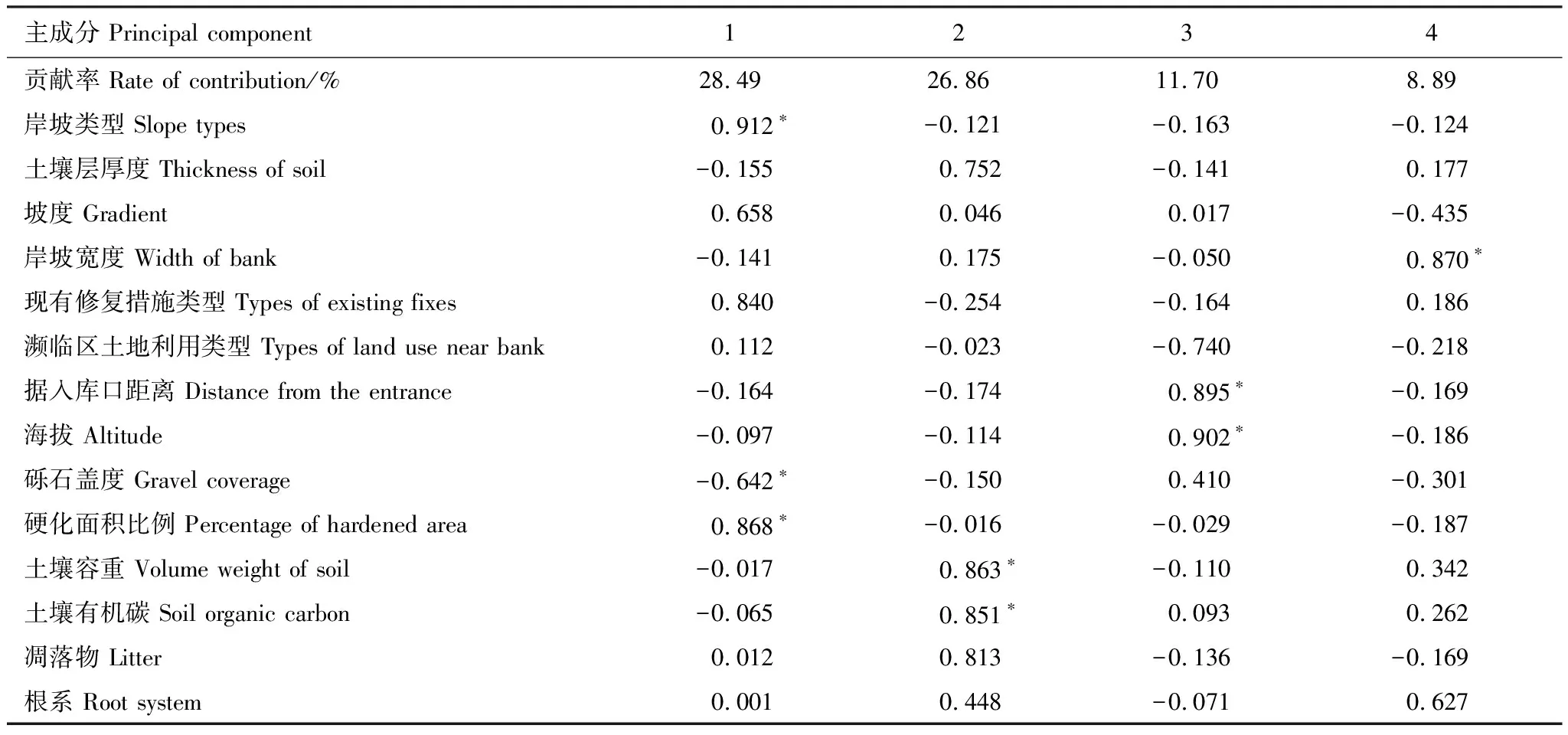
表1 4个主成分因子负荷量
*较大的因子载荷系数

图6 50个河岸带样地聚类分析图Fig.6 Cluster analysis of 50 riparian sampling
2.3 河岸带植物群落的最小面积
2.3.1 不同河岸带类型种-面积曲线的拟合
怀九河河岸带植物群落物种数随样方面积的增大而增加(表2)。自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带、人工岸坡观赏性乔灌草河岸带、人工岸坡疏乔灌草干砌石河岸带和人工岸坡浆砌石河岸带的最大样地面积分别为316.30、494.12、757.76、423.21、456.41、289.36m2,最大面积样地中植物种数分别为50、41、35、46、32、7种。
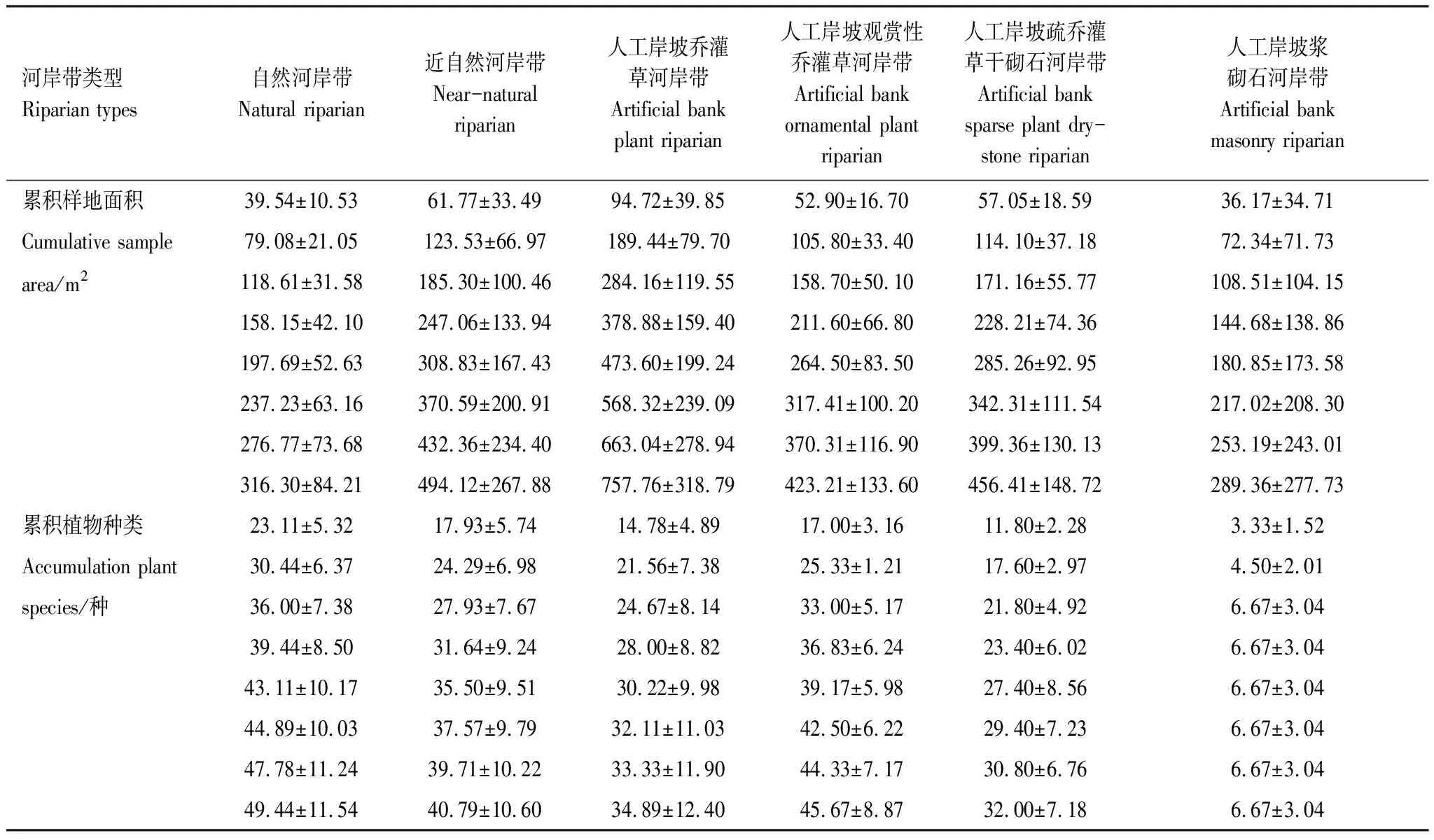
表2 怀九河不同类型河岸带植物样地物种数
6种不同类型河岸带植物群落样地面积与物种数均符合对数函数关系,除人工浆砌石河岸带R2=0.805外,其他几种类型R2值均大于0.989(图7)。
人工岸坡浆砌石河岸带所需样地面积最小,当取样面积达到109m2时,植物群落物种数的增加已保持同一水平;其次是自然河岸带,随样地面积的增加,自然河岸带植物种数迅速增加,当样地面积达到158m2时可包含群落80%植物种;人工岸坡观赏性乔灌草河岸带、近自然河岸带和人工岸坡疏乔灌干砌石河岸带均随样地面积的增加植物种数迅速增加,当样地面积分别达到212、300、285m2时可包含群落80%植物种;样地面积最大是人工岸坡乔灌草河岸带,当样地面积达到379m2时可包含群落80%植物种(图7)。
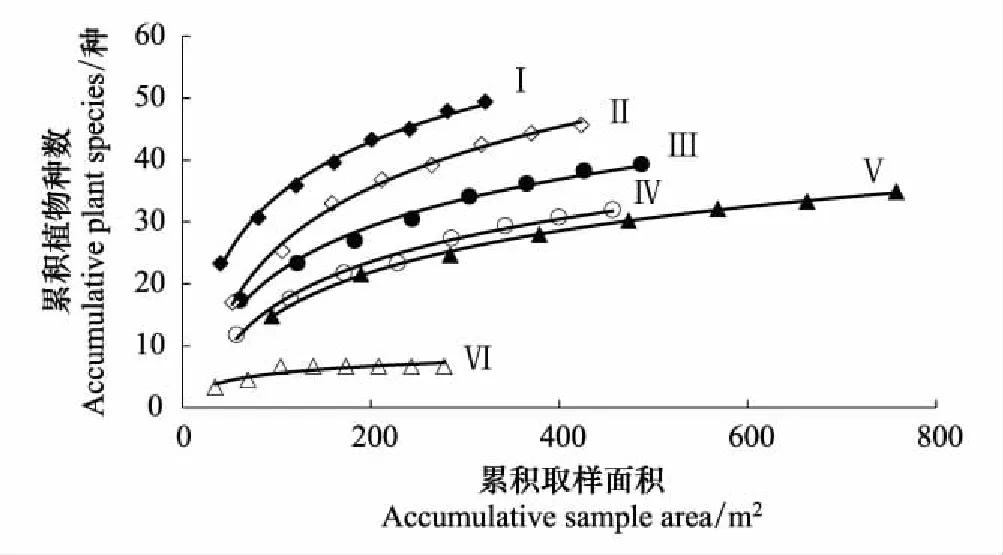
图7 北京市怀柔区怀九河不同类型河岸带植物群落种-面积关系Fig.7 Species-area relationship of different types riparian in Huaijiu River Beijing HuairouI:自然河岸带 y=12.823ln(x)-24.875, R2=0.9957, n=8, P<0.01; II:人工岸坡观赏性乔灌草河岸带 y=14.162ln(x)-39.496, R2 =0.9960, n=8, P<0.01; III:近自然河岸带 y=11.386ln(x)-30.121, R2=0.9886, n=14, P<0.01; IV:人工岸坡疏乔灌草干砌石河岸带 y=9.9367ln(x)-29.08, R2=0.9902, n=8, P<0.01; V:人工岸坡乔灌草河岸带 y=9.6463ln(x)-29.242, R2=0.9985, n=8, P<0.01; VI:人工岸坡浆砌石河岸带 y=1.6746ln(x) -2.2475, R2=0.8047, n=8, P<0.01
运用4种公式对植物群落的种面积关系进行拟合,得出系数a、b、c和R2值、AIC值(表3)。根据赤池信息量准则AIC判定模型适合度,优先考虑最小的AIC值模型,自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带和人工岸坡疏乔灌干砌石河岸带优先选取S=c-ae-bA,人工岸坡观赏性乔灌草优先选取S=aA/(1+bA),人工岸坡浆砌石河岸带优先选取S=c/(1+ae-bA)。
2.3.2 不同河岸带类型的最小样地面积
将4条种-面积曲线拟合的各参数带入公式(5)—(8),取比例因子P分别为0.7、0.8和0.9,计算得到各群落的最小取样面积如表4所示。不同类型河岸带最小样地面积差异较大且随研究精度的变大,每种河岸带类型的最小取样面积呈递增趋势。
比例因子P取0.7、0.8和0.9时,不同类型河岸带植物群落最小面积变化较大(表4)。人工岸坡浆砌石河岸带植物群落70%、80%和90%所需的最小样地面积分别为67、84、108m2;人工岸坡疏乔灌干砌石河岸带植物群落70%、80%和90%所需的最小样地面积分别为261、362、535m2,近自然河岸带、人工岸坡乔灌草河岸带70%、80%所需的最小样地面积类似,而人工岸坡观赏性乔灌草河岸带70%与近自然河岸带、人工岸坡乔灌草河岸带类似,80%和90%所需的最小样地面积迅速增加,人工岸坡浆砌石河岸带植物调查所需样地面积最小。
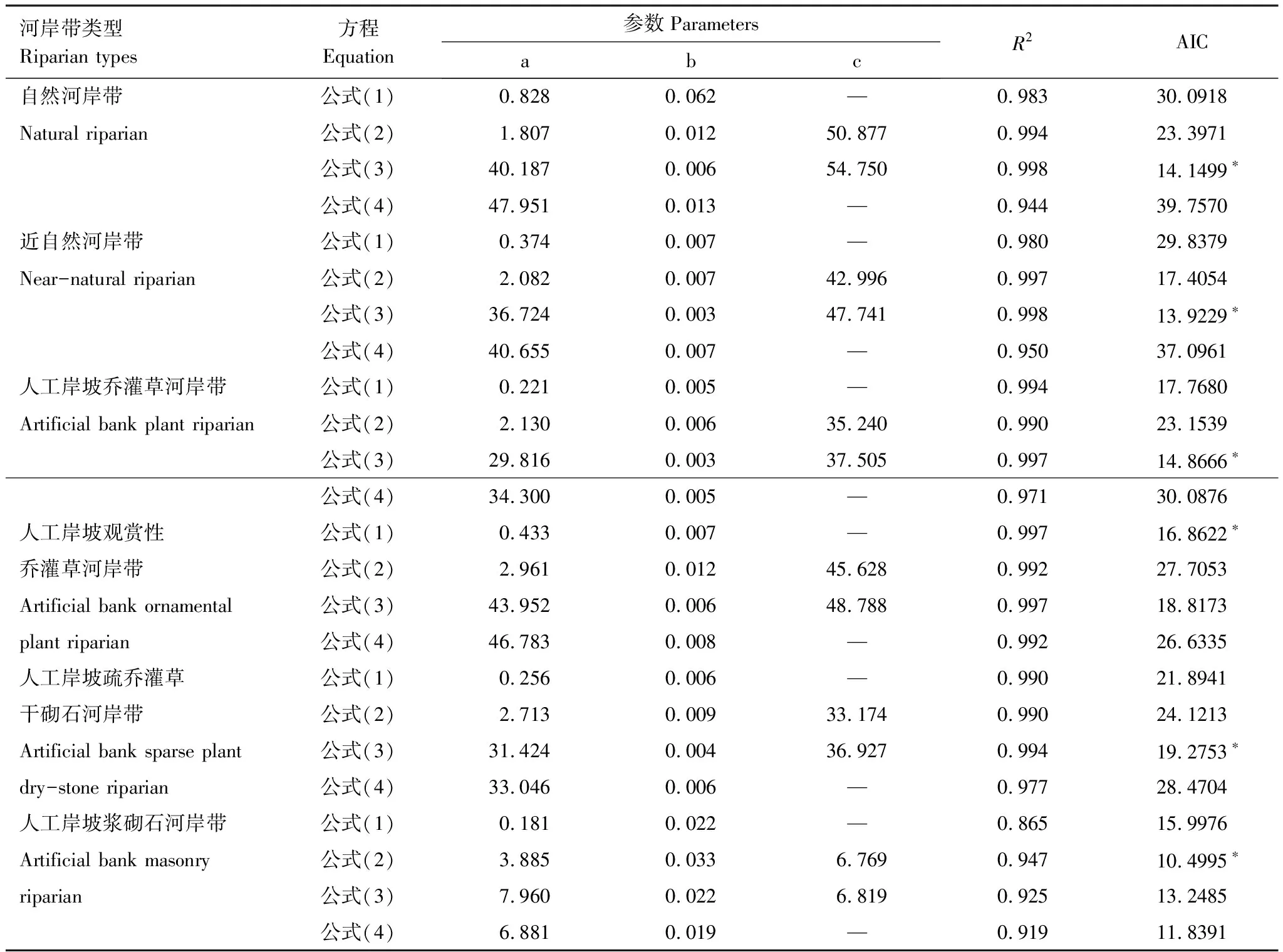
表3 种-面积曲线拟合结果
*表示有最小的AIC值
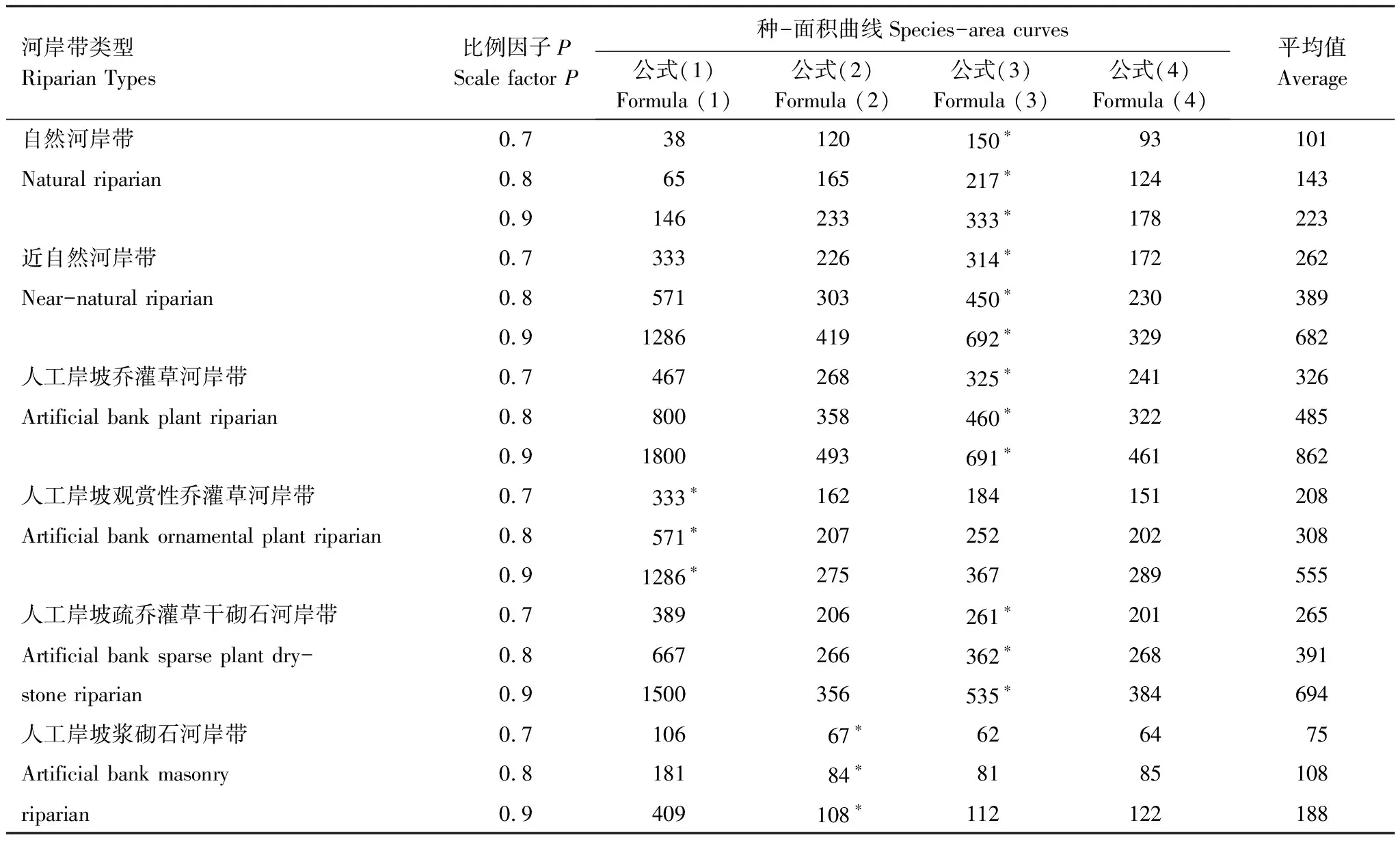
表4 种-面积曲线计算的最小面积
*表示每种类型河岸带植物调查最合理的最小样地面积
满足河岸带植物调查80%植物种,自然河岸带(217m2)、近自然河岸带(450m2)、人工岸坡乔灌草河岸带(460m2)、人工岸坡观赏性乔灌草河岸带(571m2)、人工岸坡疏乔灌干砌石河岸带(362m2)样地面积比人工岸坡浆砌石河岸带(84m2)分别高61.29%、81.33%、81.74%、85.29%、76.80%。
3 讨论
3.1 植物群落种-面积关系曲线样地布设方法
河岸带是一个特殊的陆地生态系统与水生生态系统的过渡区域,与森林植物群落、草地植物群落相比,河岸带植物群落不均匀性非常明显[26,41- 45]。近年来,大型机械的应用使得河岸带濒临区土地利用方式决定着河流河岸带的宽度。河岸带植被可以改变河岸微地貌,减少地表侵蚀,拦截泥沙,及时了解河岸带植物特征对于河流河岸带保护和生态修复意义重大[46- 48]。
最小面积是群落植物调查的基础,以往对最小面积的研究中,植物调查方法不一,不同的取样方法、样方大小、形状与数量等样方布设方式会影响曲线的拟合[49- 51]。牛俊勇等[2]运用相邻格子样方法研究了塔里木河下游荒漠河岸林群落的最小样地面积,结果表明:调查结果虽符合塔里木河下游的实际状况,但是物种数的变化不具规律性,应增强调查区域样地的频率和强度;学者利用大样地的数据优势探讨了吉林蛟河针阔混交林样地和阔叶红松林样地植物种-面积关系,研究表明:不同取样方法对种-面积关系影响显著,与巢式样方法相比,随机取样方法拟合效果更优,种-面积关系的构建与取样尺度密切相关[52- 53]。本研究充分考虑河流河岸带立地条件,基于河流河岸带坡长和坡度,采用逐步扩大样地面积的方法探讨了河流河岸带种群-面积关系,该方法较好地避免了河流河岸带岸坡不均匀造成样地布设困难的问题。
3.2 河岸带类型划分
河流河岸带是一个连续的、多变的整体,由于河流河岸带的大尺度性,以往对河流河岸带最小面积的研究主要集中于某几个断面或上、中、下游某一地区。牛俊勇等[2]沿塔里木河下游河道走向选取3个不同退化程度的典型断面进行研究;代力民等[6]在海拔为800、900、1000m的二道白河河岸带原始阔叶红松林群落分别设置一条样带进行研究;Zhou等[9]在中国青藏高原北部的阿尔卑斯山脉草地中选取5个断面进行研究;有学者指出:河岸带的类型非常丰富,从不同的角度可以划分为很多类型[54- 57]。本研究使用主成分分析提取的8项环境因子,通过聚类分析的方法将所调查50个样地分为6类,根据立地条件、植物层次和现有修复措施类型分别命名为自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带、人工岸坡观赏性乔灌草河岸带、人工岸坡疏乔灌草干砌石河岸带和人工岸坡浆砌石河岸带,系统研究了整条河流不同类型河岸带的种-面积关系,研究表明,在满足相同物种比重要求的条件下,不同河岸带类型所需调查样地面积存在明显差异。
3.3 种群-面积曲线模型的适用性
本文利用基于样地立地条件逐步扩大样地面积得到的种-面积关系数据,探讨了4种常用的种-面积关系曲线模型对最小面积的拟合结果。发现通过种-面积关系模型拟合而确定的最小样地面积与实际调查样地面积存在一定差距,本研究中通过种-面积关系拟合而得出的样地最小平均面积大于实际调查的样地最小面积,这可能是由于所选取模型和河岸带地区植物多样性较高引起的[7- 8,36,58]。4种饱和曲线对种-面积关系的拟合结果差异较大,这与王睿智和国庆喜[33]对小兴安岭阔叶红松林木本植物种-面积关系研究结果一致,与王金旺等[59]对浙江省温州市20个小型岛屿维管束植物丰富度拟合的种-面积关系结果一致。
自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带和人工岸坡疏乔灌草干砌石河岸带优先选取S=c-ae-bA,人工岸坡观赏性乔灌草优先选取S=aA/(1+bA),人工岸坡浆砌石河岸带优先选取S=c/(1+ae-bA),这说明不同类型河岸带对种-面积曲线的适用性不一,不同类型的河岸带应该选取不同的种-面积曲线拟合模型,本文仅选取4种常用的种-面积曲线且只有一种调查方法,在以后的研究中应该选取其他多种种-面积曲线,采用不同的样地调查方法进行对比,从而确定最适合的种-面积关系模型和最优的调查方法。
3.4 不同类型河岸带植物群落样地面积
大量研究表明:不同群落所需的最小样地面积不一[60- 61]。本文运用北京市怀柔区怀九河河岸带50个均匀样地探讨了不同河岸带类型下植物种-面积关系,其不同类型河岸带最小样地面积差异较大。满足河岸带植物调查80%植物种,人工岸坡浆砌石河岸带所需样地面积最小(84m2),其次是自然河岸带(217m2),人工岸坡观赏性乔灌草河岸带(571m2)所需样地面积最大,近自然河岸带、人工岸坡乔灌草河岸带、人工岸坡疏乔灌干砌石河岸带362—460m2。
人工岸坡浆砌石河岸带是由于此种修复措施破坏了原有植物生长的立地条件,浆砌石空隙中植物生长稀少;自然河岸带无修复措施,基本保持河岸带自然生境,虽然由于人地关系紧张导致的人工核桃林较多,这反而保护了河流河岸带免受人类干扰,使得较小的样地面积可以得到最多的植物种,这与牛俊勇等[2]对塔里木河下游荒漠河岸林群落最小面积的研究结果一致;人工岸坡观赏性乔灌草河岸带样地面积最大,这可能是由于为满足绿化的需要人为播撒绿化性草种,使得植物生长分布分散;包含80%植物种,人工岸坡疏乔灌干砌石河岸带所需样地面积362m2,干砌石的存在变相保护了河岸带植物的生长;近自然河岸带虽然遭受人类干扰,但是岸坡均为原始岸坡,近水区植物多样性高,随着河岸带保护意识的增加,人工岸坡乔灌草河岸带生态修复注重增加坡长,使得最小样地面积较大。
4 结论
本文通过基于河岸带立地条件逐步扩大样地面积的调查方法探讨北京山区河岸带植物群落种群-面积关系。研究表明:
(1)怀柔区怀九河河岸带植物种数为255种,隶属于70科185属。50个调查样地内乔木植物28种(10.98%)、灌木24种(11.76%)、藤本植物6种(2.35%)、一年生草本植物83种(32.55%)和多年生草本植物91种(35.69%)。河流河岸带上游植物种类(47.11±2.68)与数量(914.88±133.31)较多,中游和下游河段植物种类(32.29±5.13,31.94±3.58)和数量(640.89±126.06,527.20±71.80)相对较少。
(2)使用主成分分析提取的8项环境因子,通过聚类分析的方法将所调查50个样地分为6类,根据立地条件、植物层次和现有修复措施类型分别命名为自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带、人工岸坡观赏性乔灌草河岸带、人工岸坡疏乔灌草干砌石河岸带和人工岸坡浆砌石河岸带。
(3)根据赤池信息量准则AIC可知自然河岸带、近自然河岸带、人工岸坡乔灌草河岸带和人工岸坡疏乔灌干砌石河岸带优先选取S=c-ae-bA,人工岸坡观赏性乔灌草优先选取S=aA/(1+bA),人工岸坡浆砌石河岸带优先选取S=c/(1+ae-bA)。
(4)满足相同比例植物种调查,不同类型河岸带之间最小样地面积差异较大。满足河岸带植物调查80%植物种,人工岸坡浆砌石河岸带(84m2)与人工岸坡观赏性乔灌草河岸带(571m2)差异最大,自然河岸带(217m2)所需样地面积较小,人工岸坡疏乔灌草干砌石河岸带(362m2)、近自然河岸带(450m2)和人工岸坡乔灌草河岸带(460m2)所需样地面积较自然河岸带都有所增加。
致谢:香港大学地理系冉立山先生帮助写作,特此致谢。
参考文献(References):
[1] Cain S A. The species-area curve. The American Midland Naturalist, 1938, 19(3): 573- 581.
[2] 牛俊勇, 徐海量, 李吉玫, 杨红梅. 塔里木河下游荒漠河岸林群落最小取样面积. 生态学杂志, 2009, 28(6): 1182- 1187.
[3] Désilets P, Houle G. Effects of resource availability and heterogeneity on the slope of the species-area curve along a floodplain-upland gradient. Journal of Vegetation Science, 2005, 16(5): 487- 496.
[4] Báldi A. Habitat heterogeneity overrides the species-area relationship. Journal of Biogeography, 2008, 35(4): 675- 681.
[5] Barkman J J. A critical evaluation of minimum area concepts. Vegetatio, 1989, 85(1/2): 89- 104.
[6] 代力民, 王青春, 邓红兵, 陈高, 王庆礼. 二道白河河岸带植物群落最小面积与物种丰富度. 应用生态学报, 2002, 13(6): 641- 645.
[7] Mashayekhi M, MacPherson B, Gras R. Species-area relationship and a tentative interpretation of the function coefficients in an ecosystem simulation. Ecological Complexity, 2014, 19: 84- 95.
[8] Kong W J, Xia H J, Zhang Y. Minimum sampling area for the monitoring of herb diversity in riparian zone of temperate rivers, China. Ecological Research, 2016, 31(4): 547- 555.
[9] Zhou N, Wu J S, Shen Z X, Zhang X Z, Yang P W. Species-area relationship within and across functional groups at alpine grasslands on the northern Tibetan Plateau, China. Journal of Mountain Science, 2016, 13(2): 265- 275.
[10] Sreekar R, Huang G H L, Zhao J B, Pasion B O, Yasuda M, Zhang K, Peabotuwage I, Wang X M, Quan R C, Slik J W F, Corlett R T, Goodale E, Harrison R D. The use of species-area relationships to partition the effects of hunting and deforestation on bird extirpations in a fragmented landscape. Diversity and Distributions, 2015, 21(4): 441- 450.
[11] Novaglio C, Ferretti F, Smith A D M, Frusher S. Species-area relationships as indicators of human impacts on demersal fish communities. Diversity and Distributions, 2016, 22(11): 1186- 1198.
[12] Naiman R J, Decamps H. The ecology of interfaces: riparian zones. Annual Review of Ecology and Systematics, 1997, 28: 621- 658.
[13] Nilsson C, Berggren K. Alterations of riparian ecosystems caused by river regulation. BioScience, 2000, 50(9): 783- 792.
[14] 徐化成. 景观生态学. 北京: 中国林业出版社, 1996.
[15] Swanson F J, Fredriksen R L, McCorison F M. Material transfer in a western Oregon forested watershed//Edmonds R L, ed. Analysis of Coniferous Forest Ecosystems in the Western United States. Stroudsburg, Pennsylvania, USA: Hutchinson Ross Publishing Company, 1982.
[16] Sunil C, Somashekar R K, Nagaraja B C. Riparian vegetation assessment of Cauvery river basin of South India. Environmental Monitoring and Assessment, 2010, 170(1/4): 545- 553.
[17] 张建春. 河岸带功能及其管理. 水土保持学报, 2001, 15(6): 143- 146.
[18] Bennett P. Guidelines for Assessing and Monitoring Riverbank Health. NSW: Hawkesbury-Nepean Catchment Management Trust, 2000: 3- 4.
[19] Sweeney B W, Bott T L, Jackson J K, Kaplan L A, Newbold J D, Standley L J, Hession W C, Horwitz R J. Riparian deforestation, stream narrowing, and loss of stream ecosystem services. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(39): 14132- 14137.
[20] Bedison J E, Scatena F N, Mead J V. Influences on the spatial pattern of soil carbon and nitrogen in forested and non-forested riparian zones in the Atlantic Coastal Plain of the Delaware River Basin. Forest Ecology and Management, 2013, 302: 200- 209.
[21] Capon S J, Chambers L E, Nally R M, Naiman R J, Davies P, Marshall N, Pittock J, Reid M, Capon T, Douglas M, Catford J, Baldwin D S, Stewardson M, Roberts J, Parsons M, Williams S E. Riparian ecosystems in the 21st century: hotspots for climate change adaptation? Ecosystems, 2013, 16(3): 359- 381.
[22] Kuglerová L, Ågren A, Jansson R, Laudon H. Towards optimizing riparian buffer zones: ecological and biogeochemical implications for forest management. Forest Ecology and Management, 2014, 334: 74- 84.
[23] Thomas S M, Griffiths S W, Ormerod S J. Beyond cool: adapting upland streams for climate change using riparian woodlands. Global Change Biology, 2016, 22(1): 310- 324.
[24] 陈泓, 黎燕琼, 郑绍伟, 王玲, 何飞, 刘军, 慕长龙. 岷江上游干旱河谷灌丛群落种-面积曲线的拟合及最小面积确定. 生态学报, 2007, 27(5): 1818- 1825.
[25] Stella J C, Rodríguez-González P M, Dufour S, Bendix J. Riparian vegetation research in Mediterranean-climate regions: common patterns, ecological processes, and considerations for management. Hydrobiologia, 2013, 719(1): 291- 315.
[26] 赵清贺, 马丽娇, 刘倩, 丁圣彦, 汤茜, 卢训令. 黄河中下游典型河岸带植物物种多样性及其对环境的响应. 生态学杂志, 2015, 34(5): 1325- 1331.
[27] 刘志兰, 孙旭, 高君亮, 叶学华. 鄂尔多斯高原油蒿群落研究基本样方大小的确定. 水土保持研究, 2013, 20(3): 185- 189, 195- 195.
[28] 刘灿然, 马克平, 于顺利, 王巍. 北京东灵山地区植物群落多样性研究——种-面积曲线的拟合与评价. 植物生态学报, 1999, 23(6): 490- 500.
[29] 邓红兵, 吴刚, 郝占庆, 王庆礼, 代力民, 高林, 王建中. 马尾松-栎类天然混交林群落最小面积确定及方法比较. 生态学报, 1999, 19(4): 499- 503.
[30] 任学敏, 杨改河, 王得祥, 秦晓威, 刘振学, 赵双喜, 白宇. 太白山几类植物群落灌木及草本层的最小取样面积研究. 西北植物学报, 2011, 31(5): 1024- 1031.
[31] 要元媛, 闫明, 毕润成. 山西霍山植物群落种-面积曲线与物种多样性的关系. 生态学杂志, 2013, 32(1): 39- 44.
[32] 张蕊, 陈军强, 侯尧宸, 马丽娜, 丁路明, 龙瑞军, 尚占环. 亚高山草甸植物群落植物物种数与取样面积的关系. 生态学杂志, 2013, 32(9): 2268- 2274.
[33] 王睿智, 国庆喜. 小兴安岭阔叶红松林木本植物种-面积关系. 生态学报, 2016, 36(13): 4091- 4098.
[34] 李霞, 唐金, 万红梅. 塔里木河下游植物调查最小取样面积分析. 中国沙漠, 2011, 31(5): 1221- 1225.
[35] 高甲荣, 冯泽深, 高阳, 刘瑛. 河溪近自然评价——方法与应用. 北京: 中国水利水电出版社, 2010.
[36] 刘灿然, 马克平, 于顺利, 王巍. 北京东灵山地区植物群落多样性研究Ⅶ. 几种类型植物群落临界抽样面积的确定. 生态学报, 1998, 18(1): 15- 23.
[37] Scheiner S M. Six types of species-area curves. Global Ecology and Biogeography, 2003, 12(6): 441- 447.
[38] Anderson D R, Burnham K P, White G C. AIC model selection in overdispersed capture-recapture data. Ecology, 1994, 75(6): 1780- 1793.
[39] 贺士元, 邢其华, 尹祖堂. 北京植物志(修订版). 北京: 北京出版社, 1992.
[40] 中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 2004.
[41] Paine L K, Ribic C A. Comparison of riparian plant communities under four land management systems in southwestern Wisconsin. Agriculture, Ecosystems & Environment, 2002, 92(1): 93- 105.
[42] Suzuki W, Osumi K, Masaki T, Takahashi K, Daimaru H, Hoshizaki K. Disturbance regimes and community structures of a riparian and an adjacent terrace stand in the Kanumazawa Riparian Research Forest, northern Japan. Forest Ecology and Management, 2002, 157(1/3): 285- 301.
[43] Nagy R C, Porder S, Neill C, Brando P, Quintino R M, do Nascimento S A. Structure and composition of altered riparian forests in an agricultural Amazonian landscape. Ecological Applications, 2015, 25(6): 1725- 1738.
[44] 罗琰, 苏德荣, 吕世海, 布和, 贺晶, 谢晶杰. 辉河湿地河岸带植物物种多样性与土壤因子的关系. 湿地科学, 2016, 14(3): 396- 402.
[45] 修晨, 欧阳志云, 郑华. 北京永定河-海河干流河岸带植物的区系分析. 生态学报, 2014, 34(6): 1535- 1547.
[46] Dosskey M G, Vidon P, Gurwick N P, Allan C J, Duval T P, Lowrance R. The role of riparian vegetation in protecting and improving chemical water quality in streams. Journal of the American Water Resources Association, 2010, 46(2): 261- 277.
[47] Bullinger-Weber G, Le Bayon R C, Thébault A, Schlaepfer R, Guenat C. Carbon storage and soil organic matter stabilisation in near-natural, restored and embanked Swiss floodplains. Geoderma, 2014, 228- 229: 122- 131.
[48] Bätz N, Verrecchia E P, Lane S N. Organic matter processing and soil evolution in a braided river system. CATENA, 2015, 126: 86- 97.
[49] 余世孝, 宗国威, 陈兆莹, 臧润国, 杨应承. 随机与系统取样的生态学信息量比较. 植物生态学报, 1998, 22(5): 473- 480.
[50] Tjørve E. Shapes and functions of species-area curves: a review of possible models. Journal of Biogeography, 2003, 30(6): 827- 835.
[51] Ulrich W, Buszko J. Sampling design and the shape of species-area curves on the regional scale. Acta Oecologica, 2007, 31(1): 54- 59.
[52] 姜俊, 张春雨, 赵秀海. 吉林蛟河42 hm2针阔混交林样地植物种-面积关系. 植物生态学报, 2012, 36(1): 30- 38.
[53] 陈桂莲, 谭凌照, 范春雨, 张新娜, 张春雨, 赵秀海. 吉林蛟河阔叶红松林样地种-面积关系. 生态学报, 2017, 37(14): 4770- 4777.
[54] 夏继红, 胡玲. 生态河岸带功能区划的定性与定量研究. 水利学报, 2007, 38(S1): 542- 546.
[55] 吴建寨, 赵桂慎, 刘俊国, 姜广辉, 彭涛, 刘旭. 生态修复目标导向的河流生态功能分区初探. 环境科学学报, 2011, 31(9): 1843- 1850.
[56] 吴永祥, 王高旭, 伍永年, 丰华丽, 沈福新, 雷四华, 施睿. 河流功能区划方法及实例研究. 水科学进展, 2011, 22(6): 741- 749.
[57] Aguiar F C, Cerdeira J O, Martins M J, Ferreira M T. Riparian forests of Southwest Europe: are functional trait and species composition assemblages constrained by environment? Journal of Vegetation Science, 2013, 24(4): 628- 638.
[58] 岳隽, 王仰麟. 国内外河岸带研究的进展与展望. 地理科学进展, 2005, 24(5): 33- 40.
[59] 王金旺, 魏馨, 陈秋夏, 李效文, 杨升. 温州沿海小型海岛植物丰富度和β多样性及其影响因子. 生态学报, 2017, 37(2): 523- 540.
[60] Bonet A. Secondary succession of semi-arid Mediterranean old-fields in South-Eastern Spain: insights for conservation and restoration of degraded lands. Journal of Arid Environments, 2004, 56(2): 213- 233.
[61] Yu F H, Krüsi B, Schütz M, Schneller J J, Wildi O. Plant communities affect the species-area relationship onCarexsempervirenstussocks. Flora-Morphology, Distribution, Functional Ecology of Plants, 2008, 203(3): 197- 203.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
昆明医科大学学报(2022年2期)2022-03-29
现代园艺(2021年23期)2021-12-01
食品安全导刊(2021年20期)2021-08-30
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
河南科学(2020年3期)2020-06-02
小太阳画报(2019年4期)2019-06-11
散文诗(2018年20期)2018-05-06
少儿科学周刊·少年版(2015年11期)2015-12-17
