
圈养雄性林麝泌香规律及其与繁殖力的关系研究
2018-06-06 05:59:02周杨韩红金蔡永华程建国付文龙孟秀祥庄宝策
特产研究 2018年2期
周杨,韩红金,蔡永华,程建国,付文龙,孟秀祥,,庄宝策
(1.中国人民大学环境学院,北京 100872;2.四川养麝研究所,四川 都江堰 611830;3.山西农业大学动物科技学院,山西 太谷 030801;4.吉林省永吉县一拉溪镇畜牧兽医站,吉林 永吉 132213)
麝属(Moschusspp.)动物因其雄性分泌麝香而驰名,属濒危资源动物,目前被列为《濒危野生动植物种国际贸易公约》(CITES)附录II物种,并为我国一级重点保护动物。驯养是麝类迁地保护和麝香资源可持续保育的有效方式[1,2]。我国于1958年率先开展麝类繁育及麝香生产,实现了麝的圈养繁殖和活体取香,目前,建有100余个麝类驯养场,存栏麝达1万余头。然而,在麝类驯养实践中,一直存在繁殖力低下、麝香产量低、可泌香年限下降等问题[3,4],严重制约了我国麝类保育、驯养和麝香生产的规模化、产业化发展。
从麝类驯养开始,麝类繁育和麝香生产一直是学者和驯养业关注的焦点。郑生武等[5]报道了野生马麝的麝香分泌机理及麝香腺囊的解剖特征。Zhang[6]、尹淑媛[7]发现,麝香分泌与麝种类、饲喂模式及麝场地理气候条件等多种因素相关联,与野生麝相比较,驯养麝泌香量趋于更少,驯养环境对雌麝繁殖力存在一定影响。雄麝年龄、疾病、体况、雄激素状态及饲料营养水平等均会对麝香分泌和繁殖力有不同程度的影响[8]。圈养林麝泌香量与其泌香期长短无明显关系[9]。据李红亮等[10]报道,甘肃兴隆山圈养马麝个体年平均泌香量约7.9g(n=732),远低于野捕麝泌香量。雄性马麝泌香量在年度间呈波动规律[11];更大年龄的及野捕的个体取香率明显更高,并且在年度间存在显著差异[12,13]。
林麝是麝类迁地保育和驯养的主要麝种[14],在麝香产业中具有重要地位,大量麝香来源于林麝驯养产业[15]。有关麝类动物麝香分泌与繁殖性能之间存在着重要关联,但缺乏相关详细报道。因此,通过对林麝麝香分泌及其繁殖等有关数据的记录和统计分析,开展对圈养雄性林麝泌香规律及其与繁殖力关系的研究,可为濒危林麝迁地保护、进一步提高圈养林麝的繁殖力以及提高圈养林麝泌香量提供重要理论依据和参考数据。
1 研究方法
1.1 研究地点及动物管理
本研究于2012~2013年间在四川养麝研究所所属的马尔康麝场进行(以下均称麝场)。麝场有存栏林麝800余头,麝场所处区域地理及气候特征详见相关报道[16]。麝场设置有9个饲养区,每区包括5个~8个麝圈,每圈均由1个活动场(约100m2)和4个~7个小室(2m2~4m2)组成,小室作为雌麝躲避和分娩的场所。根据每个圈舍内小室的数量,雌麝饲养数量为4头~6头,以每头雌麝独占有1个小室为限。
每个饲养区配备专职饲养员,每日饲喂动物2次(夏季:06:00~18:00;冬季:07:00~17:00),林麝饲料主要由精饲料和人工采集的树叶组成。林麝个体均佩戴塑料唯一编码耳标,以方便个体识别及建立谱系资料。
麝场所饲养林麝发情最早见于10月下旬,于发情期前约1个月(一般为10月1日)将雄麝引入母麝圈舍中[17],采用单公多母(1∶4~6)一配到底的自然交配方式进行配种。麝场取香季节一般为每年3月,取香时,人工保定雄麝,用专用取香勺从马麝香囊口伸入囊内,取出麝香,置于草纸,吸干浮液,用电子天平称重后放入专用瓶中密封保存[10]。
1.2 研究对象及方法
本研究涉及45头雄麝,年龄均为2岁龄以上。麝类动物分娩同步化极高[16],因而以0.5岁为单位对全部个体进行年龄组区分(亚成体:2.5岁~3.5岁;成体:4.5岁~9.5岁;老龄:10.5岁及以上);按取香时麝香囊内有无成熟麝香留存进行雄麝个体区分(“未泌香雄麝”:无留存成熟麝香;“泌香雄麝”:留存成熟麝香)。
测定麝场林麝繁殖性能指标以及采集人工取香后的麝香分泌数据。相关测定指标具体为:
有效取香率=留存成熟麝香雄麝数/参与取香雄麝总数×100%;
妊娠率=妊娠雌麝数/参配雌麝总数×100%;
产仔总数=雄性林麝所配雌麝产的活仔麝总数;
仔麝早期存活率=出生头3天内存活的仔麝数/所有出生活仔麝总数×100%;
仔麝断奶成活率=断奶仔麝数/出生活仔麝数×100%;
妊娠雌麝胎均产仔数=产仔总数/妊娠雌麝总数×100%;
双胎率=产双胎雌麝数/分娩雌麝数×100%。
1.3 数据统计分析
应用Kolmogorov-Smirnov Test对取香率数据进行正态性检验。应用t检验或Mann-Whitney U test探察年龄对泌香率的效应及泌香与雌麝繁殖性能之间的关系。运用增长模型拟合林麝年龄与泌香率的关系。本研究显著性水平设为=0.05,所有数据分析通过SPSS19.0进行。
2 结果与分析
2.1 林麝泌香率和年龄的关系
如表 1所示,麝场雄麝总体取香率为 86.7%(n=45),各龄雄麝泌香率存在极显著差异(t=12.901,P=0.001)。按年龄组区分,亚成体组雄麝泌香率为100%(n=2),成体组雄麝泌香率为88.9%(n=33),老龄组雄麝泌香率为71.4%(n=5)。剔除老龄麝泌香数据后(泌香峰值年龄组),泌香率为89.50%(n=35)。如图1所示,线性模型y=120.408-6.122 5a(R2=0.471,F5,40=4.44,P=0.089),可近似模拟雄麝泌香率在年龄上的分布。依据该模型结果,19岁~20岁龄雄性林麝泌香率将趋于0值。
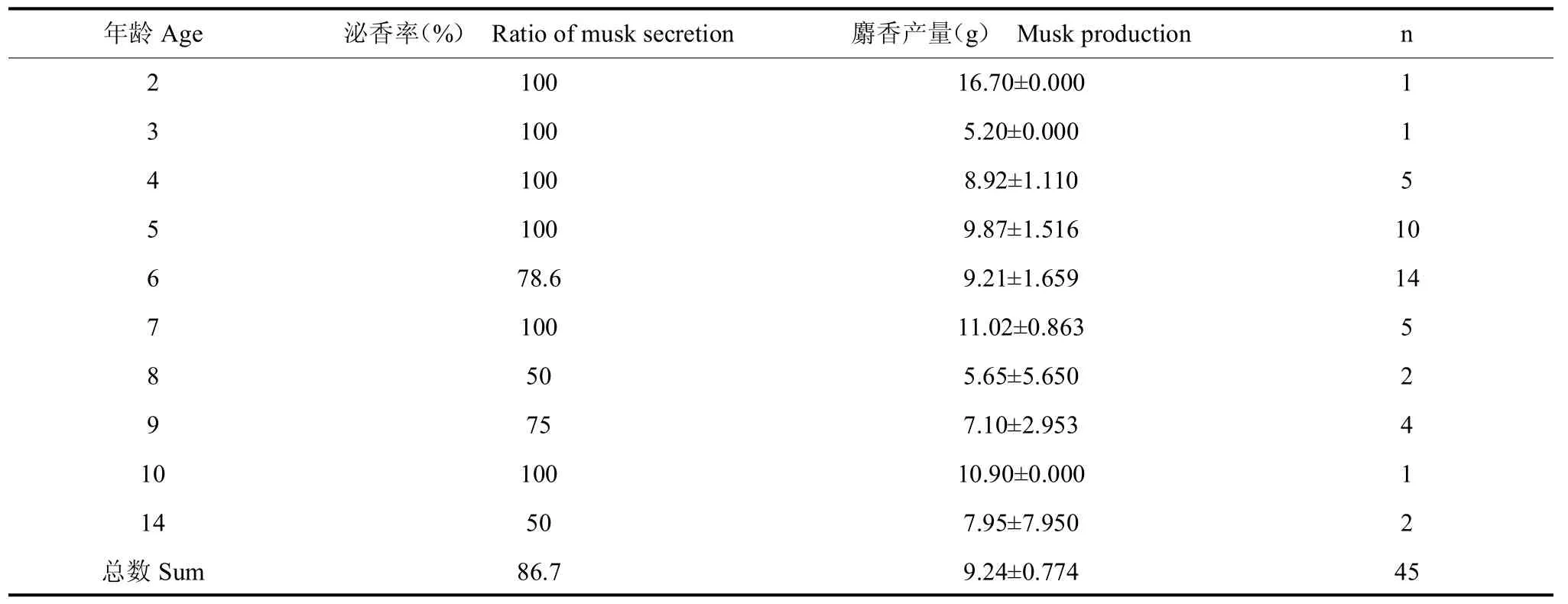
表1 各龄雄麝的泌香情况Tab.1 The musk secretion of captive male forest musk deer in age-class
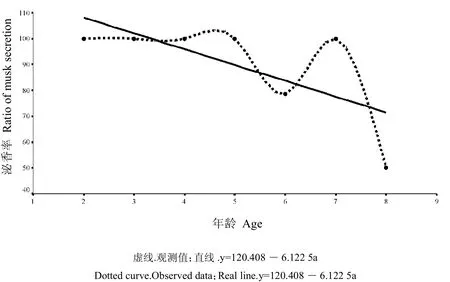
图1 雄麝泌香率在年龄的分布Fig.1 Fitted model between the musk secretion ratio and male's age-class
2.2 配种雄麝所配雌麝繁殖指数结果
对配种雄麝所配雌麝繁殖成效指数进行统计分析,如表2所示。

表2 各配种雌麝繁殖相关指数Tab.2 The reproductive index of captive female forest musk deer
2.3 配种雄麝麝香分泌与受配雌麝配怀率的关系
未泌香雄麝所配雌麝配怀率〔(80.00±4.615)%,n=6〕与泌香雄麝所配雌麝配怀率〔(74.74±3.846)%,n=39〕组间差异不显著(Kolmogorov-SmirnovTest,Z=1.004,P=0.266;t-test,t=0.523,P=0.603)。
2.4 配种雄麝麝香分泌与受配雌麝产仔数的关系
未泌香雄麝所配雌麝产仔总数(5.67±0.558,n=6)显著低于泌香雄麝所配雌麝产仔总数(7.15±0.416,n=39)(Kolmogorov-Smirnov Test,Z=0.87,P=0.436;t-test,t= 2.136,P=0.05)。
未泌香雄麝所配雌麝胎仔数(1.48±0.064,n=6)极显著低于泌香雄麝所配雌麝胎仔数(1.77±0.038,n=37)(Kolmogorov-Smirnov Test,Z=0.945,P=0.334;t-test,t= 2.966,P=0.005)。
未泌香雄麝所配雌麝所产仔麝的双胎率〔(47.50±6.437)%,n=6〕极显著低于泌香雄麝组〔(70.36±3.049)%,n=37〕(Kolmogorov-Smirnov Test,Z=0.772,P=0.554;t-test,t= 2.849,P=0.007)。
2.5 配种雄麝麝香分泌与受配雌麝产仔存活的关系
未泌香雄麝所配雌麝所产仔麝的早期死亡率〔(5.56±5.556)%,n=6〕低于泌香雄麝组〔(12.65±3.036)%,n=37〕,差异未达显著水平(Kolmogorov-Smirnov Test,Z=2.429,P=0.001;Mann-WhitneyUtest,Z=-0.970,P=0.332)。
未泌香雄麝所配雌麝所产仔麝的断奶存活率〔(69.86±8.548)%,n=6〕略低于泌香雄麝组〔(74.10±2.946)%,n=37〕,差异未达显著水平(Kolmogorov-Smirnov Test,Z=0.992,P=0.279;t-test,t=0.526,P=0.602)。
3 讨论
麝属动物的麝香分泌是一个复杂生理过程,受诸多因素影响[6],圈养麝的驯养模式与麝香分泌之间存在密切关联[17]。野生麝性独居[18]、领域性极强[19,20],而在麝类迁地保育模式中,多采用定时限饲以及群体圈养的方式,人工圈养环境对麝类动物产生综合胁迫效应,并进而导致其行为和激素水平发生相应变化。麝香分泌过程中,麝对环境胁迫的行为内分泌响应存在个体差异,部分雄麝响应圈养环境影响更为敏感,甚至会终止麝香分泌[13]。
本研究结果显示,马尔康麝场圈养雄性林麝年龄与有效取香率存在密切关联,有效泌香率随个体年龄增长而下降,亚成体取香率最高(100%),成体次之(88.9%),老龄雄麝最低(71.4%)。年龄对圈养林麝有效取香率的效应同其泌香生理特征直接相关,大部分2.5岁龄亚成体雄麝即可启动初次泌香[6],至成体麝时期,因其性成熟和体成熟已臻完善,雄麝内分泌机能旺盛,与麝香分泌相关的睾丸酮激素水平较高,麝香分泌逐渐进入盛期;老龄麝雄激素分泌逐渐减少,并因个体体况、疾病等因素的影响,麝香分泌功能明显下降,部分雄麝甚至停止泌香。程建国等[9]报道,雄性林麝2岁~8岁为麝香分泌的峰值年龄段,取香率可达96%,尹淑媛和戴卫国[7]的研究表明,圈养林麝泌香高峰年龄段为2岁~6岁,取香率可达95%。上述研究中取香率高于本研究中林麝泌香峰值年龄组(亚成体和成体麝)的泌香率(89.5%),原因在于本研究与上述研究中样本空间不同,目标雄麝均为配种后雄麝,配种对麝香泌香率产生一定影响。此外,本研究表明,依据麝香取香率增长模型,从统计上预测马尔康雄麝停止泌香的年龄为19岁~20岁,在麝香产业实践中,也有近20岁雄麝个体少量泌香的记录[1,21]。综合对马麝的研究,可以判定驯养麝具备商业价值的麝香分泌高峰年龄组为亚成体及成体,年龄区间为3岁~8 岁[10,12]。
本研究表明,未泌香雄麝所配雌麝的产仔总数、胎仔数及双胎率等繁殖性能指标均显著低于泌香雄麝组。雄麝麝香分泌生理过程与其发情交配紧密相关,受到同样的雄性性激素调控作用[22~24],外源性雄性激素(睾丸酮)的输入可重启已停止泌香的雄麝再次进行麝香分泌[7]。本研究虽未涉及有香与无香雄麝激素水平对照,但未泌香雄麝在繁殖交配季节的性激素水平推测应明显低于泌香雄麝,较低的性激素水平将影响其发情交配活动强度、效率及有效性(如求偶、打斗、追配、爬跨及配后守护等),从而影响繁殖成效和繁殖力,导致未泌香雄麝所配雌麝的繁殖性能(产仔总数、胎仔数及双胎率等)低于泌香雄麝所配雌麝。在本研究中,未泌香雄麝和泌香雄麝所配雌麝的配怀率并无明显差异,这可能受到未泌香雄麝样本(仅6头)较小的影响。
本研究中未泌香雄麝所配雌麝的配怀率和仔麝的早期存活率均高于泌香雄麝组,这与麝类驯养业的配种制度及麝香的生物活性存在关联。麝香可诱导中止妊娠,且随着妊娠时间增加,逐渐进入后期,诱导效应逐渐明显[25]。在麝类驯养的配种制度下,1头雄麝与4头~6头雌麝进行配对,同一个圈群内雌麝并不同期发情,当雄麝与某一发情雌麝配种后,雄麝并不移走,将继续与后期发情雌麝进行交配。圈养麝的发情交配活动可持续4个月以上[26],而麝香是典型的气体型外信息素[18,19,25],先期配怀成功的雌麝可能承受同一圈舍或邻近圈舍内雄麝麝香长期、高强度的诱导,极可能导致孕麝早期妊娠终止,因而泌香雄麝所配雌麝配怀率相对更低。部分雄麝配种结束后虽可能从雌麝圈导出,但为避免产生过多惊扰,一般仅将其隔离于配后雌麝邻近圈舍,泌香雄麝所分泌的麝香将持续对配怀雌麝后期妊娠产生高强度作用,增加其流产、死胎及死产等情况的发生,增加了仔麝的早期死亡。未泌香雄麝对所配雌麝中怀孕雌麝的诱导作用较弱或缺失,因而雌麝仔麝早期死亡率反而较低,更易存活。基于分析,麝类产业实践过程中,可对其配种制度和饲养管理进行优化,在明确雌麝配怀成功后,及时隔离配种雄麝与怀孕雌麝,饲养区域保持足够距离,以尽量减少雄麝麝香分泌对雌麝妊娠的负面诱导作用,提高雌麝繁殖生产力。
此外,麝类动物的繁殖和麝香分泌均具有强季节性,林麝发情交配季节为10月至翌年1月,而麝香分泌期处于5月~7月[6]。本研究中配种雄麝取香于发情交配结束后进行(3月左右),因而不能判断本研究中的未泌香雄麝是配种前即无麝香分泌,抑或是因配种而导致麝香被损失殆尽,使麝香囊内无成熟麝香留存。进一步的研究可开展相关的行为内分泌研究,及通过麝香分泌监测进行对照验证,相关结论具有重要意义,有助于探究麝类动物麝香分泌机理及优化麝类驯养生产实践和保育管理。
[1]Homes V.On the Scent:Conserving Musk Deer-the Uses of Musk and Europe's Role in its Trade[M].Brussels:TRAFFIC,Europe,1999.
[2]Yang Q,Meng X,Feng Z,et al.Conservation status and causes of decline on musk deer in China[J].Biological Conservation,2003,109:333-342.
[3]Meng X,Zhou C,Hu J,et al.The musk deer farming in China[J].Animal science,2006,82:1-6.
[4]Meng X,Liu D,Feng J,et al.Asian medicine:exploitation of wildlife[J].Science,2012,335:1168.
[5]郑生武,皮南林.马麝的麝香腺囊、麝香、数量与狩猎[J].兽类学报,1984,4(1):35-42.
[6]Zhang B L.Musk-deer,Their capture,domestication and care according to Chinese experience and methods[J].Unasylva,1983,35:16-24.
[7]尹淑媛,戴卫国.野麝家养研究综述[J].成都科技大学学报,1991,55(1):81-90.
[8]黄步军,盛和林,徐宏发.饲料营养对林麝麝香产量和品质的影响[J].动物学研究,1998,19(4):296-300.
[9]程建国,罗燕,乔美萍,等.影响林麝泌香量的因素[J].特产研究,2002,(3):15-18.
[10]李红亮,黎勇,周密,等.驯养马麝麝香产量及其影响因素[J].中国农业科学,2012,45(3):522-528.
[11]康发功,刘志霄,陈建国,等.甘肃兴隆山养麝场圈养高山麝的产香量[J].兽类学报,2008,28(2):221-224.
[12]孟庆辉,刘春兰,潘世成,等.甘肃兴隆山保护区驯养马麝麝香生产的有效取香率分析[J].西北农林科技大学学报:自然科学版,2011,39(12):53-59.
[13]Meng X X,Gong B C,Ma G.,et al.Quantified analyses of musk deer farming in China:a tool for sustainable musk production and ex situ conservation[J].Asian-Australian Journal of Animal Sciences,2011,24(10):1473-1482.
[14]Yang Q,Meng X,Feng Z,et al.Conservation status and causes of decline on musk deer in China[J].Biological Conservation,2003,109:333-342.
[15]Meng X,Zhou C,Hu J,et al.The musk deer farming in China[J].Animal science,2006,82:1-6.
[16]Meng X,Yang Q,Xia L,et al.Timing and synchrony of parturition in alpine musk deer[J].Folia Zoologica,2003,52(1):39-50.
[17]盛和林,刘志霄.中国麝科动物[M].上海:上海科学技术出版社,2007.
[18]Green M J B.Diet composition and quality in.Himalayan musk deer based on faecal analysis[J].Journal of Wildlife Management,1987,51:880-892.
[19]Green M J B.Scent-marking in the Himalayan musk deer[J].Journal of Zoology,1987,(1):721-737.
[20]杨奇森,冯祚建,王祖望,等.西藏东南部地区马麝家域的研究[J].兽类学报,1998,18(2):87-94.
[21]张保良.麝的驯养[M].北京:农业出版社,1979.
[22]任战军.人工养麝于取香技术[M].北京:金盾出版社,2003.
[23]王永生.麝香生产技术[M].北京:中国农业出版社,2004.
[24]Shrestha M N.Animal welfare in the musk deer[J].Applied Animal Behavior Science,1998,59:245-250.
[25]Sokolov V E.Chemical communication of some species of rodents,ungulates and carnivores[J].Acta Zoologica Fennica,1984,171:67-69.
[26]Meng X,Yang Q,Xia L,et al.The temporal estrous patterns of female alpine musk deer in captivity[J].Applied Animal Behavior Science,2003,82:75-85.
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23 13:46:54
今日农业(2021年11期)2021-08-13 08:53:40
辽金历史与考古(2021年0期)2021-07-29 01:06:54
今日农业(2020年14期)2020-12-14 19:47:34
天津中医药(2020年10期)2020-12-10 18:31:15
天津中医药(2020年5期)2020-06-01 12:16:06
科技传播(2019年22期)2020-01-14 03:06:54
民用飞机设计与研究(2019年4期)2019-05-21 07:21:24
天然产物研究与开发(2018年8期)2018-09-10 05:48:26
下一代英才(酷炫少年)(2017年10期)2017-11-22 02:16:08
