
浙江古田山不同森林类型两栖动物群落结构比较*
2018-06-01 06:54陈声文钱海源泮琦斌吴驰星
浙江师范大学学报(自然科学版) 2018年2期
陈声文, 钱海源, 王 宇, 泮琦斌, 吴驰星, 邵 晨
(1.古田山国家级自然保护区管理局,浙江 开化 324300;2.浙江师范大学 化学与生命科学学院,浙江 金华 321004)
0 前言
两栖动物是森林生态系统中重要的组成部分,对森林生态系统的物质循环和能量流动起到至关重要的作用.两栖动物也是脊椎动物从水生过渡到陆生的重要过渡类群,其生活史周期离不开水、皮肤通透性高等特点使其对栖息环境的变化特别敏感,因此,两栖动物是森林生态环境质量监控的理想指示物种,两栖动物的群落结构特征还是评价该地区生态系统质量的重要参数.因此,对两栖动物群落结构特征的研究是生态系统物种多样性和生态恢复研究的重要内容.近年来,国内外学者在两栖动物的区域物种调查、多样性分析等方面取得了较为丰硕的研究成果[1-3].
此外,由于栖息地环境改变带来的影响日益突出,因而有关两栖动物生态位分析及其与生态系统恢复和利用方面的研究不断涌现,对保护区的资源特征分析及物种保护也愈加重视[4-5].早在20世纪中期,就陆续有学者对古田山自然保护区两栖动物进行调查,但结合保护区森林类型对两栖动物群落特征的分析至今未见专门报道.
本研究通过对古田山自然保护区内不同森林类型中两栖动物物种多样性和优势度进行研究,掌握不同类型森林栖息地中两栖动物对环境的适应状态,从而预测两栖动物群落发展趋势,可为森林生境下两栖动物群落特征研究方面提供基础资料,也为两栖动物物种保护研究提供新的思路,还能为自然保护区的生态系统健康评价及科学管理提供理论依据.
1 材料与方法
1.1 研究地概况
浙江古田山国家级自然保护区(29°10′19.4″~29°17′41.4″ N,118°03′49.7″~118°11′12.2″ E)位于浙江省西部的开化县苏庄境内,总面积约8 107 hm2.属亚热带季风气候区,气候温暖湿润,四季分明.年平均温度为15.3 ℃,最高温度为38.1 ℃(7月),最低为-6.8 ℃(1月) .古田山雨量充沛,年均降雨量1 963.7 mm,年降雨天数大约为140 d,相对湿度为92.4%,山系水流经苏庄镇折入江西省德兴市的乐安江,随后流入鄱阳湖,再汇入长江[6].古田山母岩以花岗岩为主,主要土壤类型为红壤、黄红壤和红黄壤,局部地区有山地草甸土,土壤pH为5.5~6.5,呈酸性[7].植被类型主要有常绿阔叶林、常绿落叶阔叶混交林、针叶阔叶混交林和针叶林[8].
1.2 样地选择及两栖动物调查
在对古田山森林进行实地踏查的基础上,依据古田山的植被组成和古田山国家级自然保护区网格化监测体系将古田山的森林类型划分为阔叶林、马尾松林、杉木林、油茶林和茶叶林等5种.但由于茶叶林分布面积狭小,且多位于受人类干扰较大的村民聚居区,因此,本研究并未在茶叶林开展相关的调查工作.
2014年7—8月和2015年7—8月于上述阔叶林、马尾松林、杉木林和油茶林4种森林类型中分别选取具有一定海拔梯度(300~1 000 m)的样带(1 000 m×5 m)各8条以调查两栖动物的群落组成及其特征.由于两栖动物运动能力较弱,且多群居于静水坑或栖于溪沟边,因此,在本调查中所设置的样带长度和宽度足以充分发现两栖动物.于夜间18点到凌晨2点采用遇见法或鸣声法记录在样带中出现的全部两栖动物,并用手持全球定位仪(GPS)记录各调查样本所在的地理坐标,物种的鉴定依据《中国动物志:两栖纲》[9]和《中国两栖动物及其分布彩色图鉴》[10].
1.3 数据分析
用Shannon-Wiener物种多样性指数、Pielou均匀度指数、Simpson优势度指数和Margalef丰富度指数表示两栖动物群落结构特征.各指数的计算公式为:
Shannon-Wiener多样性指数:H′=-∑Pi×lnPi;
Simpson优势度指数:Cs=1-∑Ni×(Ni-1)×N-1×(N-1)-1;
Pielou均匀度指数:J=H′×(lnS)-1;
Margalef丰富度指数:D=(S-1)×(lnN)-1.
其中:Ni为第i个类群的个体数量;N为所有类群的个体数量;Pi为第i个物种占总物种数的比例;S表示物种数.
不同森林类型中的两栖动物群落相似性采用Jaccard指数[11-12],Sorenson指数[11-12]和Morisita-Horn指数[13-14]进行分析.Jaccard指数和Sorenson指数是群落相似性分析中最常用的二元相似性系数,它们重点关注于群落中物种的出现和未出现数据,因此这两种方法非常相似,且相对易于计算和较能直观了解;但如大多数物种仅出现在某个森林类型的群落中一样,Sorenson指数则相较于Jaccard指数具有更高的敏感性;Morisita-Horn指数不仅要考虑出现和未出现数据,还需要考虑群落内物种的丰富度,因此,该方法不会受到样本量的影响,同时对大多数物种的数量也非常的敏感.因此,结合这3种方法来分析将能更全面地理解不同森林类型中两栖动物的群落相似性.各指数的计算公式为:
Jaccard指数:CJ=j×(a+b-j)-1;
Sorenson指数:CS=2j×(a+b)-1.
式中:j为两个群落中共有的两栖动物物种数;a,b分别代表2个群落的物种数.
式中:nai,nbi分别是群落A和群落B中第i个物种的个体数;Na和Nb分别是群落A和群落B的总个体数.
群落相似性系数计算值在0.751~1.000为极相似;在0.501~0.750为中等相似;在0.251~0.500为中等不相似;在0.000~0.250为极不相似.
用Excel 2010软件对两栖动物的群落结构和群落相似性进行统计分析.用SPSS 16.0软件对古田山不同森林类型的两栖动物群落结构差异显著性进行检验.首先,使用Kolmogorov-Smirnov test对两栖动物群落的物种多样性、优势度、均匀度和丰富度进行正态分布检验.若数据不服从正态分布,则将数据进行自然对数(ln)转换后再进行后续统计分析.然后应用单因素方差分析(One-Way ANOVA)的Duncan′s检验进行不同森林类型间的多重比较.显著性水平α=0.05.
2 结 果
2.1 不同森林类型的两栖动物群落组成
调查共记录到两栖动物2目6科12属13种709个个体.其中,在马尾松林和杉木林中都记录到最多的11个物种,马尾松林中华南湍蛙(Amolopsricketti)、花臭蛙(Odorranaschmackeri)和东方蝾螈(Cynopsorientalis)是优势类群(个体数占总数>10%),黑斑侧褶蛙(Pelophylaxnigromaculatus)和饰纹姬蛙(Microhylafissipes)没有分布;杉木林中华南湍蛙和淡肩角蟾(Megophrysboettgeri)是优势类群,黑斑侧褶蛙、棘胸蛙(Quasipaaspinosa)、秉志肥螈(Pachytritongranulosus)和东方蝾螈罕见(个体数占总数<1%)或没有分布;在阔叶林中记录到最多的个体数243个,华南湍蛙、淡肩角蟾、花臭蛙和秉志肥螈是阔叶林中的优势类群,而弹琴蛙(Nidiranaadenopleura)、黑斑侧褶蛙、饰纹姬蛙、东方蝾螈和中华蟾蜍(Bufogargarizanz)则在阔叶林中罕见或没有分布;油茶林则记录到最少的个体数136个,其中华南湍蛙和花臭蛙是优势类群,而福建大头蛙(Limnonectesfujianensis)、饰纹姬蛙、秉志肥螈和东方蝾螈则罕见或没有分布.此外,泽陆蛙(Fejervaryamultistriata)和大绿臭蛙(Odorranagraminea)在4种森林类型中均有一定数量的个体,华南湍蛙在4种森林类型栖息地中出现的频度最高(见表1).

表1 不同森林类型中两栖动物的类群与数量组成
注:多度为在该类型栖息地中所调查到的该物种总的个体数;频度为该物种在该类型栖息地8条样带中的出现频率.
2.2 不同森林类型两栖动物群落多样性特征
两栖动物的个体数与物种数在不同森林类型之间差异均不显著(P>0.05).各林型中,阔叶林生境中两栖动物物种的多样性指数、优势度指数和均匀度指数的值均为最高,而杉木林在丰富度指数的值最大(见表2).在不同森林类型中,两栖动物的多样性指数、优势度指数、均匀度指数和丰富度指数差异不显著(P>0.05).
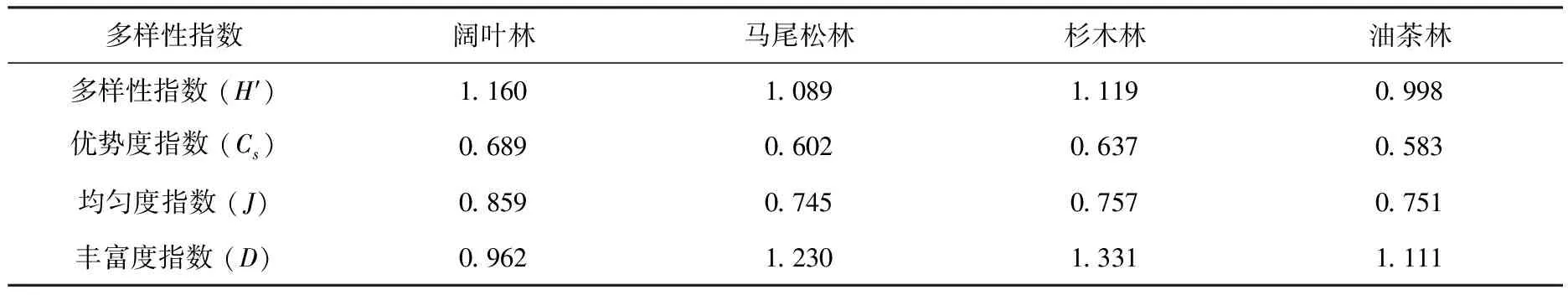
表2 浙江古田山不同森林类型两栖动物多样性
2.3 不同森林类型两栖动物群落的相似性
在不同的森林类型中,3个相似性指数均显示两栖动物在杉木林和油茶林中的相似度最高;Jaccard指数和Sorenson指数显示两栖动物群落在硬阔林和杉木林之间的相似性最低,而Morisita-Horn指数则显示在硬阔林和马尾松林之间两栖动物群落的相似性最低(见表3).
3 讨 论

表3 浙江古田山不同森林类型两栖动物群落相似性指数
本研究在古田山自然保护区的4种森林类型32条样带中共调查到13个两栖类物种,占浙江全省两栖动物资源总数2目9科46种[15]的28.3%,占古田山自然保护区两栖动物资源总数2目7科17种[1]的76.5%.从整个保护区中两栖动物的分布情况来看,物种的分布在不同的林型中并不均衡,其中有8个物种分布到所有的4种森林类型中,但在不同森林类型中的两栖动物物种在数量上存在差异,这表明两栖动物对于不同的森林类型栖息地具有一定的选择性,与当地独特的植被、食物组成等自然环境有密切的关系.
华南湍蛙在不同森林类型中出现的频率和数量均为最多,占所有两栖类物种数量记录总体的近半数,说明该物种是古田山自然保护区中两栖动物组成的优势种,也是常见种.其次,花臭蛙和淡肩角蟾在4种林型生境的调查中数量相当,仅次于华南湍蛙,但花臭蛙在阔叶林、马尾松林和油茶林3种森林类型中均为优势种,而淡肩角蟾则在阔叶林和杉木林中有较广分布.此外,花臭蛙和淡肩角蟾仅在阔叶林中有较多的重叠,虽二者同属流溪型生活的物种,但花臭蛙和淡肩角蟾在个体大小上的明显差异可能是造成二者生态位分化的原因,因此,二者能同时分布到阔叶林.而在其他林型,虽然二者数量均较多,但并不重叠,所以也可以很好地共存.
多样性指数可以反映物种群落内的丰富度和均匀度,其与生物和非生物因子间有着密切的关系[16].阔叶林和杉木林中两栖动物的Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数都高于其他2种林型,说明在这2种林型生境中两栖类物种的分布较均匀.Margalef丰富度指数显示杉木林中物种的丰富度最高,马尾松林次之.调查过程中发现,杉木林和马尾松林样地边缘地带分布着较多的农田和洼地,这与饰纹姬蛙、黑斑侧褶蛙、东方蝾螈等两栖动物的生活环境相符[17],因此,在这2种林型的物种丰富度指数较高.两栖动物的生存和繁殖都不能彻底摆脱对水环境的依赖,表3的结果显示,古田山自然保护区各林型生境中各项指数差异不显著,物种组成较为相似,这说明了两栖动物群落分布受沟田、溪流分布的影响较大,而与森林类型可能并无直接关系.
动物群落相似性取决于群落本身的种类组成、数量结构和群落间的共有种组成[18].古田山自然保护区4种林型生境中两栖动物群落结构,在组成上存在一定的共同性,Jaccard指数和Sorenson指数主要是通过4种林型生境中两栖动物物种种类相似度情况进行群落相似度分析,二者的结果呈现的趋势一致,杉木林与油茶林中两栖类物种的相似度最高,这2种林型生境中植被结构单一,物种种类大部分重合,唯一不同的是杉木林地的农田中发现饰纹姬蛙物种分布.两栖动物群落物种相似度指数最低的是硬阔林与杉木林,说明2种林型中的物种相似性最小,在阔叶林林边溪沟中发现有一定数量的秉志肥螈群落,杉木林中未见分布,其他林型生境中也较少见.杉木林中有3种物种在阔叶林中没有发现,分别是弹琴蛙、黑斑侧褶蛙和饰纹姬蛙,推测不同两栖类物种对森林生境具有一定的选择性.Morisita-Horn指数主要是基于不同林型生境中群落个体数进行的相似度分析,与前面相似度指数分析结果不同的是,硬阔林与马尾松林中群落相似度最低,该2种林型中群落物种个体总数虽是最多的2个生境,但从表1的数据中可以看出2种林型中群落物种数据多寡不一,各群落物种个体数量差异明显.据此可以推测两栖类动物喜欢选择水分充足、植被多样、人为干扰较小的栖息环境,2种生境中两栖物种分布的差异可能还与食物类型、竞争关系等复杂因素有关[19].
由于两栖动物主要分布在溪谷、水田等地的峭壁上或石体隐蔽处,本次调查主要采用遇见法和鸣声法,可以见闻相补,为夜间野外调查记录工作增加了可靠性和准确性.古田山保护区内森林生境分为纯林和混交林,调查结果显示,不同森林型中两栖动物物种多样性差异并不显著,为满足生物多样性保护的需求及林木生态系统的科学管理,今后在对两栖动物的管理和保护中可重点关注硬阔-马尾松混交林或硬阔-杉木混交林.经调查发现的浙江省古田山自然保护区内13种两栖动物全部列入《国家保护的有益的或有重要经济、科学研究价值的陆生野生动物名录》,因此具有重要的保护价值.两栖动物处于生物链的中间地位,其数量的多少将直接影响着生物链物质的循环和能量的传递过程及整个生态系统的健康发展,地方政府需在发展旅游业的同时,严禁随意捕杀两栖动物,如棘胸蛙、泽陆蛙和中华蟾蜍等具有食用和药用价值的物种,加大对保护区内生物多样性的保护力度.
参考文献:
[1]方小斌,周奕琳,杨璐伊,等.浙江省古田山国家级自然保护区两栖爬行动物资源现状[J].四川动物,2013,32(1):125-130.
[2]王同亮,程林,兰文军,等.江西武夷山国家级自然保护区两栖动物多样性及海拔分布特点[J].生态学杂志,2015,34(7):2009-2014.
[3]田凯,汪正祥,雷耘,等.湖北崩尖子自然保护区两栖爬行动物多样性研究[J].四川动物,2016,35(3):452-458.
[4]Roloff G J,Grazia T E,Millenbah K F,et al.Factors associated with amphibian detection and occupancy in Southern Michigan forests[J].Journal of Herpetology,2011,45(1):15-22.
[5]Steen D A,Mcgee A E R,Hermann S M,et al.Effects of forest management on amphibians and reptiles:Generalist species obscure trends among native forest associates[J].Open Environmental Sciences,2010,4:24-30.
[6]开化林业志编写组.开化林业志[M].杭州:浙江人民出版社,1988:41-343.
[7]楼炉焕,金水虎.浙江古田山自然保护区种子植物区系分析[J].北京林业大学学报,2000,22(5):33-39.
[8]于明坚,胡正华,余建平,等.浙江古田山自然保护区森林植被类型[J].浙江大学学报:农业与生命科学版,2001,27(4):375-380.
[9]费梁,胡淑琴,叶昌媛,等.中国动物志:两栖纲[M].北京:科学出版社,2009.
[10]费梁,叶昌媛,江建平.中国两栖动物及其分布彩色图鉴[M].成都:四川科学技术出版社,2012.
[11]马克平,刘灿然,刘玉明.生物群落多样性的测度方法[J].生物多样性,1995,3(1):38-43.
[12]覃林.统计生态学[M].北京:中国林业出版社,2009:63-75.
[13]何先进,吴鹏飞,崔丽巍,等.坡度对农田土壤动物群落结构及多样性的影响[J].生态学报,2012(12):3701-3713.
[14]Dent D H,Wright S J.The future of tropical species in secondary forests:A quantitative review[J].Biological Conservation,2009,142(12):2833-2843.
[15]江建平,谢锋,臧春鑫,等.中国两栖动物受胁现状评估[J].生物多样性,2016,24(5):588-597.
[16]陈灵芝,马克平.生物多样性科学:原理与实践[M].上海:上海科学技术出版社,2001.
[17]吴迪,岳峰,罗祖奎,等.上海大莲湖湖泊湿地两栖动物群落分布及生境选择模式[J].复旦学报: 自然科学版,2011,50(3):268-273.
[18]杨富亿,吕宪国,娄彦景,等.松嫩湖群鱼类群落相似性[J].生态学报,2015,35(4):1022-1036.
[19]庾太林,张良建,刘晓辉,等.广西北部湾海陆交错带两栖动物对生境的选择[J].四川动物,2013,32(6):883-888.
猜你喜欢
落叶果树(2021年6期)2021-02-12
农民致富之友(2020年8期)2020-05-11
红土地(2019年11期)2019-12-04
现代园艺(2018年1期)2018-03-15
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
军营文化天地(2017年6期)2017-06-28
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
军营文化天地(2016年3期)2016-05-26
体育科技(2016年2期)2016-02-28
