
Trillium tschonoskii maxim extract attenuates abnormal Tau phosphorylation
2018-05-31 09:02:38HongBinLuoNanShangWenZhiXieDeJianWenMinQuShengHuangShaShaFanWeiChenNanQiaoMouXiangYuLiuQinChenFengFengXieJunXuLi
中国神经再生研究(英文版) 2018年5期
Hong-Bin Luo , Nan Shang Wen-Zhi Xie De-Jian Wen Min Qu, Sheng Huang Sha-Sha Fan Wei Chen Nan-Qiao Mou Xiang-Yu Liu Qin Chen Feng-Feng Xie Jun-Xu Li
1 Hubei Key Laboratory of Biological Resource Protection and Utilization, Hubei University for Nationalities, Enshi, Hubei Province, China
2 Medical College, Hubei University for Nationalities, Enshi, Hubei Province, China
3 Institute of Neurological and Psychiatric Comorbidity, Hubei University for Nationalities, Enshi, Hubei Province, China
4 Hubei Provincial Key Laboratory for Applied Toxicology, Hubei Provincial Center for Disease Control and Prevention, Wuhan, Hubei Province,China
5 Department of Pharmacology and Toxicology, Jacobs School of Medicine and Biomedical Sciences, University at Buffalo, State University of New York, Buffalo, NY, USA
Introduction
Alzheimer’s disease (AD) is a significant neurodegener‐ative disease often seen in the aged population, and the typical symptoms include diffuse brain atrophy, cognitive impairment, deteriorating memory loss, and personality disorders (Querfurth and LaFerla, 2010). The disease is his‐topathologically characterized by neurofibrillary tangles of the Tau protein and senile plaques of amyloid beta peptides in certain brain regions, and in most serious cases leads to neuronal cell death (Blennow et al., 2010). Given the huge economic and public heath toll of AD, various potential therapeutic approaches have been attempted. However,currently, there is no effective treatment available for AD.Therefore, the development of new and effective treatments is direly needed.
Many factors may contribute to the development of AD.Large‐scale epidemiological studies have found that one of the strongest independent risk factors for AD is hyperho‐mocysteinemia (Ravaglia et al., 2005; Obeid and Herrmann,2006). Homocysteine (Hcy) is a sulfur‐containing amino acid, which is a crucial intermediate product of the remethi‐onine cycle (Bi et al., 2014). Animal studies show that Hcy treatment results in AD pathogenesis in the hippocampus and other brain regions in rats (Flicker et al., 2004; Pache‐co‐Quinto et al., 2006), and Hcy treatment is increasingly used as a rodent model of AD.
Trillium Tschonoskiimaxim (TTM) is the rhizome and root ofLiliaceae Trillium, a traditional Chinese medicinal plant, which contains various steroidal saponins and al‐kaloids from Liliaceae, and can be found in most areas of central and western China (Li et al., 2005). In traditional folk medicine, TTM has been used for the treatment of dizziness, neurasthenia, hypertension, bleeding, traumatic injury, headache, and AD (Qiu et al., 2016; Wu et al., 2016;Yan et al., 2016). However, the clinical efficacy of TTM ex‐tract for these diseases has not been established by carefully controlled clinical studies, and the underlying mechanisms are largely unknown. This study attempted to evaluate the effects of TTM extract against the memory impairment in‐duced by Hcy in a rodent model of AD.
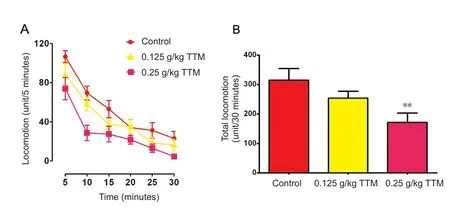
Figure 1 Effects of TTM extract on the spatial memory deficit assessed by open field test in Alzheimer’s disease model rats.
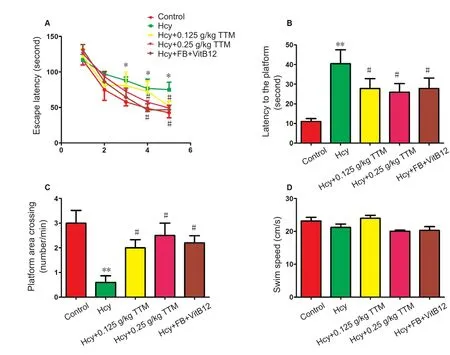
Figure 2 Protective effects of TTM extract on spatial memory impairment induced by Hcy in Alzheimer’s disease model rats.
Materials and Methods
Experimental animals
A total of 60 male wild‐type Sprague‐Dawley rats aged 3 months old and weighing 200—225 g were supplied by the Experimental Animal Central of Hubei Province, China(license No. 42000600003985). All rats were allowed free access to food and water. The experimental procedure fol‐lowed the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023,revised 1986). Rats were kept in cages under a 12‐hour light/dark cycle with the light on from 7:00 a.m. to 7:00 p.m.
AD model establishment and TTM extract intervention
A total of 50 rats were randomly assigned into five different treatment groups (n= 10 per group). Rats were injected by vena caudalis with DL‐Hcy (400 μg/kg; Sigma Chemical Co.,St. Louis, MO, USA; four groups) to induce AD models or saline (control group) daily for 14 days.
TTM was purchased from Xinghe Pharmaceuticals(Bozhou, China). TTM extract was prepared in our laboratory. Brie fly, 12.5 g TTM was boiled twice with water, 1 hour each, and the resultant solutions were combined, filtered with a 200‐mesh sieve, and centrifuged at 3 000 r/min for 15 minutes. The supernatant was heat‐concentrated to 100 mL at 100 °C, which led to the final drug solution of 0.125 g/mL.Rats were treated with the following, daily for 7 days: saline(control group, Hcy group), 0.125 g/kg TTM extract (Hcy+ 0.125 g/kg TTM group), 0.25 g/kg TTM extract (Hcy +0.25 g/kg TTM group), or folate (4 mg/kg; Yabang Aipusen Co., Yancheng, Jiangsu, China) plus vitamin B12 (VitB12;250 μg/kg; Yunpeng Co., Linfen, Shanxi, China) (Hcy + FB+ VitB12 group) by intragastric administration. The Hcy metabolism process requires VitB6, VitB12 and folate as a positive control group (Finkelstein, 1998).
A separate experiment that used three groups of rats consisted of control, 0.125 g/kg TTM, and 0.25 g/kg TTM. Rats received intragastrical administration of TTM solution 30 minutes before locomotion testing (n= 10 per group). The locomotion testing was conducted to examine the acute effects of gavage TTM extract administration on locomotor activity using the locomotion measurement apparatus as de‐scribed below.
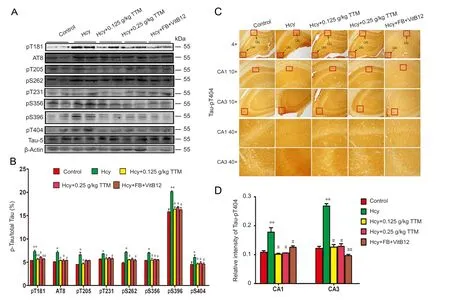
Figure 3 Phosphorylation levels as measured by different epitopes of Tau in hippocampus of Alzheimer’s disease model rats.
Behavioral experiments
Open field test
After gavage administration of saline or TTM extract for 7 days, locomotion was assessed using the open field test,as previously described (Zhu et al., 2014). Brie fly, the loco‐motor activity of rats was measured using a commercially available apparatus (YLS‐1B, Shandong Academy of Medi‐cal Sciences, China). This apparatus consists of a controller unit and four circular black acrylic locomotion units (30 cm in diameter and 30 cm in height). The photo beam sensor located in the center of the cover of the locomotion units re‐cords the spontaneous locomotor activity. Locomotor activity was measured during a 30‐minute test period with each count indicating one beam break by the animal.
Morris water maze test
The Morris water maze test is a behavioral experiment that assesses hippocampal‐dependent learning and memory abilities. The standard Morris water maze procedure was used with minor modifications (Morris, 1984). Brie fly, rats were trained to find a hidden platform in the water maze for 5 consecutive days, 4 trials per day with a 30‐second inter‐val. The rats were not allowed to search for the platform for more than 60 seconds, after which they were guided to the platform and remained there for 30 seconds. The swimming pathways and latencies of each rat to find the hidden plat‐form were recorded by Noldus video tracking system (Etho‐vision, Noldus Information Technology, Holland) during each test. One day after the last TTM extract treatment, rats were placed in the water maze to find the hidden platform.One hour later, the platform was removed, and the rats were placed in the opposite quadrant for 1‐minute test periods.
Western blot assay
After behavioral testing, rats were euthanized and their hip‐pocampi were removed from the brain. Hippocampal tissue was homogenized in a buffer containing tris (hydroxymeth‐yl) aminomethane (Tris; Sigma Chemical Co.)‐HCl (pH 7.6)10 mM, Na3VO41 mM, NaF 50 mM, benzamidine 1 mM,edeticacid 1 mM, and phenylmethylsulfonyl fluoride (Sigma Chemical Co.) 1 mM. Three volumes of the homogenized tissue were added to one volume of an extracting buffer containing Tris‐HCl (pH 7.6) 200 mM, 8% sodium dodecyl sulfate (Sigma Chemical Co.), and 40% glycerol, and extracts were boiled in a water bath for 10 minutes. The lysates were sonicated brie fly and centrifuged at 12,000 ×gfor 5 minutes.Protein concentration of the supernatants was measured by the bicinchoninic acid Protein Assay Kit (Beijing Kan‐gweishiji Biotech Ltd., Beijing, China). The proteins were separated by sodium dodecyl sulfate‐polyacrylamide gel electrophoresis on 10% gel and transferred to a nitrocellulose membrane (GE Healthcare Bio‐Sciences, Pittsburgh, PA,USA). After blocking in 3% nonfat milk for 1 hour at 25°C,the membranes were incubated with primary antibodies at 4°C overnight. The blots were then incubated with an‐ti‐mouse or anti‐rabbit IgG conjugated to IRDye™(800CW)(1:5000; LI‐COR Biosciences, Lincoln, NE, USA) for 1 hour at 25°C and visualized with the Odyssey Infrared Imaging System (LI‐COR Biosciences). Primary antibodies used in western blotting studies included the following: mouse an‐ti‐rat Tau‐5 (1:1000), mouse anti‐rat phosphorylated PP2AC(P‐PP2Ac) (1:300) and mouse anti‐rat methylated‐PP2Ac(M‐PP2Ac) (1:300) monoclonal antibodies from Abcam(Cambridge, United Kingdom); mouse anti‐rat β‐actin monoclonal antibody (1:1000) from Proteintech (Chicago,IL, USA); mouse anti‐rat AT8 monoclonal antibody (1:300)from Thermal Fisher Scientific (Waltham, MA, USA); rabbit anti‐pT181 (1:300), pT205 (1:300), pT231 (1:1000), pS262(1:300), pS356 (1:300), pS396 (1:1000), pS404 (1:1000) poly‐clonal antibodies from Signalway Antibody LLC (College Park, MD, USA); mouse anti‐rat PP2Ac (1:300) and DM PP2Ac (1:300) monoclonal antibodies from Cell Signaling(Danvers, MA, USA); and mouse anti‐rat methylesterase(PME) monoclonal antibody (1:300) from Millipore (Burlington, MA, USA). These molecules are all critical signaling molecules involved in Tau phosphorylation. Results were analyzed using Image‐Pro Plus software (Media Cybernet‐ics, Shanghai, China).
Immunohistochemistry
For immunohistochemical studies, rats were sacrificed by overdose with chloral hydrate (1 g/kg) after behavioral test‐ing and perfused through the aorta with 100 mL 0.9% NaCl followed by 400 mL phosphate buffer containing 4% para‐formaldehyde. Brains were removed and post fixed in perfu‐sate overnight and then cut into sections (20 μm) with a vi‐bratome (S100, TPI; Leica, Nussloch, Germany). Fixed AD and the age‐matched control group brain cubes containing the hippocampal formation were cut into sections (40 μm)with a freezing microtome (Kryostat 1720; Leitz, Wetzler,Germany) after anhydration in 25% sucrose. The sections of rat brain and AD hippocampi were collected consecutively in phosphate‐buffered saline for immunohistochemical staining. Free floating sections were blocked with 0.3% H2O2in absolute methanol for 30 minutes and nonspecific sites were blocked with bovine serum albumin for 30 minutes at room temperature. Sections were then incubated overnight at 4°C with the same primary antibodies. Immunoreaction was developed using Histostain™‐SP kits (Thermal Fish‐er Scientific) and visualized with diaminobenizidine. For each primary antibody, three to five consecutive sections from each brain were used. The images were observed using a microscope (BX60; Olympus, Tokyo, Japan). The immunoreactivity of anti‐pS404 (1:100; rabbit polyclonal antibodies; Signalway Antibody LLC) was probed using Rhodamine Red‐X conjugated goat anti‐rabbit IgG (1:100;Thermal Fisher Scientific). The images were observed using a laser scanning confocal microscope (FV500, Olympus).Semi‐quantitative analysis was performed using Image‐Pro Plus software.
Golgi staining
The rats were anesthetized with chloral hydrate and per‐fused with 400 mL normal saline containing 0.5% sodium nitrite, followed by 400 mL 4% paraformaldehyde solution.The brains were further perfused with 400 mL dying solution made of 5% chloral hydrate, 5% potassium dichromate,and 4% paraformaldehyde. The brains were then incubated in dying solution for 3 days in the dark and transferred to silver solution containing 1% silver nitrate (Shanghai Fine Chemicals Institute, Shanghai, China) for 3 days in the dark;30‐μm brain sections of hippocampal tissue were cut using a vibratome (Leica, Wetzlar, Germany). All the dendritic spines on the first and third branches of the apical dendrites of five pyramidal neurons in the hippocampus CA1 area were pho‐tographed with an oil‐immersed objective. The total dendritic spines per branch were counted and the mean and standard deviation were found. Results were analyzed using Image J software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Data are expressed as the mean ± SEM. All results were analyzed using GraphPad Prism 5.0 statistical software(GraphPad Software, La Jolla, CA, USA) by one‐way analysis of variance or two‐way analysis of variance (Morris water maze) followed by a least significant differencepost hoctest.A level ofP< 0.05 was considered statistically significant.
Results
TTM extract ameliorated the development of spatial memory deficit in Hcy-induced AD rat model
As shown in Figure 1, intragastric administration with TTM extract (0.125, 0.25 g/kg) acutely reduced the general loco‐motor activity in a dose‐dependent manner assessed by open field test. The dose of 0.25 g/kg significantly reduced the locomotor activity (P= 0.0045), suggesting that TTM may have a sedative effect at a large dose. These data provided evidence for the subsequent experiments on dose selection,such that no doses higher than 0.25 g/kg were used. During the five acquisition training sessions, rats reliably learned to locate the platform, however they performed differently on the latency to reach the platform (Figure 2A). Hcytreated rats had significantly increased escape latency, which was significantly prevented by chronic TTM extract treatment or a combined folate and VitB12 treatment. Significant main effects of drug treatment [F(4, 172) = 39.17,P< 0.001] and sessions [F(4, 172) = 7.62,P< 0.001] were identified according to two‐way analysis of variance.Post hocanalysis found that escape latency significantly increased in the Hcy‐treat‐ed rats during the third to fifth acquisition sessions (P<0.05). Importantly, 0.25 g/kg TTM extract treatment and the combined folate‐VitB12 treatment significantly reversed the increased escape latency during the fourth and fifth ac‐quisition sessions (P< 0.05). The platform was removed 1 hour later and a probe trial was initiated. Rats that received Hcy treatment spent a significantly longer time to reach the platform area and showed a fewer number of crossings over the platform area compared with control rats (P< 0.01 for latency andP< 0.01 for number of crossing). TTM extract and folate‐VitB12‐treated rats were significantly faster to reach the platform position, and crossed more times over the platform, compared with Hcy‐treated rats (F[4,49] =4.38,P< 0.01;F[4, 49] = 5.17,P< 0.01, respectively) (Figure 2B, C). Importantly, there was no significant difference in the swim speed among the groups (Figure 2D).
TTM extract reduced Tau phosphorylation in Hcy-induced AD rat model
A previous study has shown that high plasma Hcy induces Tau hyperphosphorylation that mimics AD (Zhang et al.,2008), and the aberrant hyperphosphorylation level of Tau causes dendritic loss and neurodegeneration (Wang et al.,2010). Therefore, we tested the effect of TTM extract on Tau phosphorylation in Hcy‐treated rats. The levels of phosphor‐ylated Tau at Ser202 (AT8), Thr181 (pT181), Ser262 (pS262),Thr205 (pT205), Ser356 (pS356), Thr231 (pT231), Ser396(pS396), and Ser404 (pS404) were significantly increased in the hippocampal tissue of Hcy‐treated rats (P < 0.05; Figure 3A, B). Similarly, the immunoreactive level of Tau1, an indicator of Tau (pT404) phosphorylation, in Hcy‐treated rats was significantly stronger than that of control rats (P< 0.01;Figure 3C, D). Treatment with two doses of TTM extract,or combined folate‐VitB12, significantly attenuated the Hcy‐induced Tau hyperphosphorylation (Figure 3) with the exception of Thr205 (pT205) and Thr231 (pT231), for which only 0.125 g/kg dose of TTM extract achieved statistical significance. These data suggest that TTM extract attenuates Hcy‐induced Tau hyperphosphorylation, which has similar efficacy as the combined treatment of folate and VitB12.
Specific protein kinases and protein phosphatases partici‐pate in the regulation of Tau phosphorylation (Wang et al.,2010). Hcy can increase Tau phosphorylation through inactivating PP2A (Zhang et al., 2008; Chai et al., 2013). Consis‐tent with the literature, we found that the phosphorylation of PP2Ac at Tyr307, which inhibits PP2Ac activity, increased in the hippocampus of Hcy‐treated rats (Figure 4A, B). It should be noted that the total PP2Ac was not altered among different treatment conditions. Treatment with TTM extract and the folate‐VitB12 combination remarkably reduced the Hcy‐induced PP2Ac phosphorylation in the hippocampus.In addition, because elevated plasma Hcy decreases meth‐ylated PP2Ac (activated form) and increases demethylated PP2Ac (inactivated form) (Zhang et al., 2008), which is crucial for modulating PP2Ac activity, we also examined the effects of TTM extract on methylated and demethyl‐ated PP2Ac. As expected, methylated PP2Ac significantly decreased, and demethylated PP2Ac significantly increased in Hcy‐treated rats (P< 0.01; Figure 4A, B). Importantly,treatment of TTM extract and the combination of folate and VitB12 normalized the changes in methylated and de‐methylated PP2Ac levels in the hippocampus of Hcy‐treated rats. A specific PME is responsible for the methylation of PP2AC (Lee et al., 1996), and Hcy treatment obviously in‐creases the level of PME (Zhang et al., 2008). Therefore, we examined whether TTM extract could alter the expression ofthis enzyme. Consistent with the literature, Hcy‐treated rats showed a noticeably increased level of PME (Figure 4C, D).TTM extract treatment reduced the increased level of PME in Hcy‐treated rats (Figure 4C, D). The combined use of folate and VitB12 also normalized the Hcy‐induced increase of PME activity (Figure 4C, D).
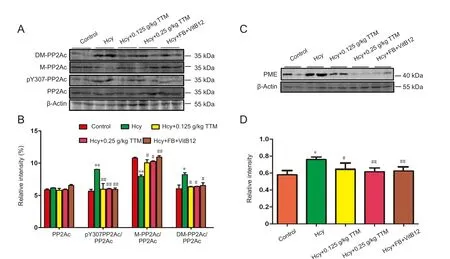
Figure 4 Effect of TTM extract on the activity of PP2A in the hippocampus of Alzheimer’s disease model rats.
TTM extract increased the number of dendritic spines in hippocampal CA1 area of Hcy-treated rats
It is believed that dendritic spines, particularly those located on pyramidal cells, are essential postsynaptic elements for cortical excitatory synapses and are critical structural elements underlying learning, memory, and cognition (Roberts et al., 2010). Importantly, long‐term memory is partially mediated by the emergence of new dendritic spines to enhance a particular neural pathway (Hofer et al., 2009). To deter‐mine whether Hcy treatment affects neural morphological structures and to investigate the underlying mechanisms of the role of TTM extract in memory retention, we performed Golgi staining to detect the morphological changes of den‐drites and the density of spines in pyramidal neurons of rat hippocampus. Hcy reduced the total number of dendritic spines in hippocampal neurons (P< 0.01). Treatment with TTM extract preserved the dendritic morphologies and the spine density, for example, thin dendritic spines (P< 0.05).The same effect was also achieved by supplementing rats with folate‐VitB12 combination (P< 0.01; Figure 5).
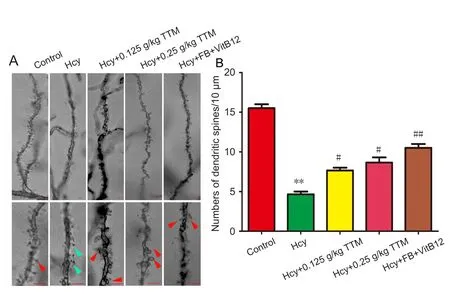
Figure 5 Representative dendritic spine in hippocampal CA1 of the rat brain.
Discussion
The primary findings of the present study were that TTM extract prevented Hcy‐induced spatial memory impairment and memory retention, with a similar efficacy to the combination of folate and VitB12. In the study exploring the underlying mechanisms, it was found that Hcy led to in‐creased Tau phosphorylation, inactivated protein phosphatase PP2Ac activity, and reduced the number of dendritic spines in hippocampal neurons. Importantly, TTM extract prevented all of the biochemical changes induced by Hcy regarding Tau pathophysiology and neuronal morphology.Combined, these results suggest that the traditional Chinese medicine TTM extract may be able to prevent Hcy‐related memory impairment, possibly through its protective effects on neurons against Hcy‐induced aberrant Tau phosphorylation and dendritic spine functions.
TTM is a medicinal plant that has been used to treat var‐ious neuropsychiatric disorders for many years in China.Recent progress on understanding the pharmacological effects and related mechanisms has just begun to emerge.For example, TTM extract has been shown to attenuate the enhancement of superoxide dismutase and glutathione peroxidase activity, and the decrease of malondialdehyde in the hippocampus of a rat model of AD induced by okadaic acid (Huang et al., 2008). These results demonstrate strong antioxidant activity of TTM extract. In addition, furotrilliu‐moside extracted from TTM extract can inhibit lipopolysac‐charide‐induced inflammation by upregulating phosphory‐lated phosphoinositide‐3‐kinase and Akt, inhibiting nuclear factor‐κB translocationviathe prevention of inhibitory IκBα phosphorylation and degradation, and suppressing the signaling pathway of mitogen‐activated protein kinase (Yan et al., 2016). However, the understanding of the mechanisms underlying the potential therapeutic efficacy of TTM extract in AD remains very limited. To further verify its therapeutic effects and understand the potential neurobiological mechanisms, we established a rat AD modelviaHcy treatment and examined the protective effect of TTM extract and the underlying mechanisms.
Consistent with the literature, Hcy treatment led to obvi‐ous memory impairment and impaired memory retention in the Morris water maze assay at doses that did not remark‐ably alter the spontaneous activity. Importantly, the high dose that did dramatically reduce locomotion (0.25 mg/kg)did not produce sustained suppression, as the locomotor activity of the rats was maintained at control level on the next day (data not shown). The finding that TTM extract mark‐edly restored the performance of rats in this assay confirmed the potential therapeutic efficacy of TTM extract in this AD model, suggesting that TTM extract may be effective in treating certain forms of AD. In an effort to understand the underlying mechanisms of TTM extract, we examined one of the key neuronal mechanisms that contribute to the development of AD, Tau hyperphosphorylation.
Under normal conditions, the balance of Tau phosphorylationin vivois a fine‐tuned process by two different mech‐anisms: kinases, including glycogen synthase kinase‐3β and cyclin‐dependent kinase‐5, can phosphorylate Tau, whereas phosphatases, such as PP2A, can dephosphorylate Tau. In AD, the balance is perturbed, and Tau is hyperphosphory‐lated and aggregates to form neurofibrillary tangles (Goedert,1993). It has been shown that Hcy treatment primarily affects the activity of phosphatases (e.g., PP2Ac inactivation),whereas the activity of kinases, such as glycogen synthase ki‐nase‐3β, is unchanged (Zhang et al., 2008). Consistent with the literature, this study showed that Tau phosphorylation was increased together with an increase of the inactivated form of PP2Ac due to increased phosphorylation at the Tyr‐307 site. Importantly, TTM extract treatment markedly reduced the level of Tau phosphorylation and reduced the inactivated form of PP2Ac, suggesting that regulating Tau phosphorylation might be the mechanism that contributed to the improved memory retention in Hcy‐treated rats.
Hyperhomocysteinemia and low folate/VitB12 status have been long known to independently elevate homocysteine levels, which impairs the methylation potential in the brain(Selhub et al., 2010; Bottiglieri, 2013). The catalytic activity of PP2Ac is actively modulated by its methylation: methylat‐ed active form and demethylated inactive form. During this process, the PP2A‐specific methylesterase PME‐1 binds to PP2A, subsequently removing the methyl group and inac‐tivating PP2A (Sontag and Sontag, 2014). Hcy‐treated rats show increased PME‐1 levels, decreased methylated PP2Ac and increased demethylated PP2Ac (Zhang et al., 2008),which was also demonstrated in the present study. Remark‐ably, TTM extract treatment markedly reversed the changes of PP2A activity induced by Hcy. The efficacy of TTM ex‐tract was equivalent to the supplemental treatment of folate/VitB12.
Finally, dendritic spines play a critical role in the process of learning, memory, and cognition (Roberts et al., 2010).In Hcy‐treated animals, Hcy decreases the average dendritic length and reduces the number of dendritic branches in cortical neurons (Xia et al., 2014). Hcy also decreases the spine density in the dendritic branches of hippocampal CA1 pyramidal neurons (Xia et al., 2014). Here, we found that Hcy‐treated rats had a markedly reduced number of den‐dritic spines in the hippocampus, which is consistent with a previous study (Xia et al., 2014). Importantly, TTM ex‐tract dramatically reversed the reduction to a level that was similar to the effect of folate/VitB12 treatment. This result suggests that TTM extract normalizes the altered dendritic morphology in Hcy‐treated rats.
In conclusion, the traditional Chinese medicine TTM ex‐tract improved memory deficits in rats treated with Hcy, and the effect was at least partially mediated through the nor‐malization of the Hcy‐induced increase of Tau phosphorylation and spine morphological changes. These results suggest that TTM extract may have some value against certain forms of AD and deserves further investigation. These results also pave the way for refining the primary biologically active ingredients in TTM for further studies, which may lead to effective treatments against AD.
Acknowledgments:We are very grateful to Professor De-pei Yuan from Medical College, Hubei University for Nationalities, Enshi, Hubei Province, China for his technical help in this study.
Author contributions:HBL designed this study and was responsible for paper authorization. HBL and JXL wrote the paper and provided technical support. NS and WZX performed experiments. DJW provided technical support. MQ and SH participated in critical revision of the paper for intellectual content. SSF, WC, NQM, XYL, QC and FFX were responsible for data collection, analysis and interpretation. All authors approved the final version of the paper.
Conflicts of interest:The authors declare no competing financial interests.Financial support:This work was supported in part by grants from the National Natural Science Foundation of China, No. 81260172,81660223; the Opening Foundation of Hubei Key Laboratory of Biological Resource Protection and Utilization of China, No. PKLHB1318; the Science and Technology Innovation Team Project of Hubei University for Nationalities of China, No. MY2011T005; the Doctoral Fund of Hubei University for Nationalities of China, No. MY2012B015; the Natural Science Foundation of Hubei Province of China, No. 2017CFB451. The conception, design, execution, and analysis of experiments, as well as the preparation of and decision to publish this manuscript, were made independent of any funding organization.
Institutional review board statement:The study protocol was approved by the Experimental Animal Ethics Committee of Hubei University for Nationalities of China (approval No.HB-01409). The experimental procedure followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1985).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Bi JJ, Zhang SY, Luo WY, Guan XQ (2014) Relationships of plasma ho‐mocysteine levels and other clinical indexes with the polymorphisms of MTHFR gene C677T among the young people in Shenzhen area.Zhongguo Zuzhi Gongcheng Yanjiu 18:6871‐6877.
Blennow K, Hampel H, Weiner M, Zetterberg H (2010) Cerebrospinal fluid and plasma biomarkers in Alzheimer disease. Nat Rev Neurol 6:131‐144.
Bottiglieri T (2013) Folate, vitamin B(1)(2), and S‐adenosylmethionine.Psychiatr Clin North Am 36:1‐13.
Chai GS, Jiang X, Ni ZF, Ma ZW, Xie AJ, Cheng XS, Wang Q, Wang JZ, Liu GP (2013) Betaine attenuates Alzheimer‐like pathological changes and memory deficits induced by homocysteine. J Neuro‐chem 124:388‐396.
Finkelstein JD (1998) The metabolism of homocysteine: pathways and regulation. Eur J Pediatr 157 Suppl 2:S40‐44.
Flicker L, Martins RN, Thomas J, Acres J, Taddei K, Norman P, Jam‐rozik K, Almeida OP (2004) Homocysteine, Alzheimer genes and proteins, and measures of cognition and depression in older men. J Alzheimers Dis 6:329‐336.
Goedert M (1993) Tau protein and the neurofibrillary pathology of Alzheimer’s disease. Trends Neurosci 16:460‐465.
Hofer SB, Mrsic‐Flogel TD, Bonhoeffer T, Hubener M (2009) Expe‐rience leaves a lasting structural trace in cortical circuits. Nature 457:313‐317.
Huang HC, Zhong RL, Cao P, Wang CR, Yang DG (2008) Effects of Goutengsan on model of Alzheimer dementia in rats by AlCl3.Zhongguo Zhong Yao Za Zhi 33:553‐556.
Lee J, Chen Y, Tolstykh T, Stock J (1996) A specific protein carboxyl methylesterase that demethylates phosphoprotein phosphatase 2A in bovine brain. Proc Natl Acad Sci U S A 93:6043‐6047.
Li Q, Xiao M, Guo L, Wang L, Tang L, Xu Y, Yan F, Chen F (2005)Genetic diversity and genetic structure of an endangered species,Trillium tschonoskii. Biochem Genet 43:445‐458.
Morris R (1984) Developments of a water‐maze procedure for studying spatial learning in the rat. J Neurosci Methods 11:47‐60.
Obeid R, Herrmann W (2006) Mechanisms of homocysteine neurotoxicity in neurodegenerative diseases with special reference to dementia. FEBS Lett 580:2994‐3005.
Pacheco‐Quinto J, Rodriguez de Turco EB, DeRosa S, Howard A,Cruz‐Sanchez F, Sambamurti K, Refolo L, Petanceska S, Pappolla MA (2006) Hyperhomocysteinemic Alzheimer’s mouse model of amyloidosis shows increased brain amyloid beta peptide levels. Neu‐robiol Dis 22:651‐656.
Qiu Y, Li RP, Liu S, Tan ZX, Chen LQ, Liu H, Wu H (2016) The pro‐tective effects of ethanol extract of Trillium tschonoskii Maxim. on hypoxiaischemia brain damage in neonatal rats. Zhongguo Yao Li Xue Bao 32:1548‐1552.
Querfurth HW, LaFerla FM (2010) Alzheimer’s disease. N Engl J Med 362:329‐344.
Ravaglia G, Forti P, Maioli F, Martelli M, Servadei L, Brunetti N, Por‐cellini E, Licastro F (2005) Homocysteine and folate as risk factors for dementia and Alzheimer disease. Am J Clin Nutr 82:636‐643.
Roberts TF, Tschida KA, Klein ME, Mooney R (2010) Rapid spine stabilization and synaptic enhancement at the onset of behavioural learning. Nature 463:948‐952.
Selhub J, Troen A, Rosenberg IH (2010) B vitamins and the aging brain. Nutr Rev 68 Suppl 2:S112‐118.
Sontag JM, Sontag E (2014) Protein phosphatase 2A dysfunction in Alzheimer’s disease. Front Mol Neurosci 7:16.
Wang X, Blanchard J, Kohlbrenner E, Clement N, Linden RM, Radu A, Grundke‐Iqbal I, Iqbal K (2010) The carboxy‐terminal fragment of inhibitor‐2 of protein phosphatase‐2A induces Alzheimer disease pathology and cognitive impairment. FASEB J 24:4420‐4432.
Wu H, Qiu Y, Shu Z, Zhang X, Li R, Liu S, Chen L, Liu H, Chen N (2016)Protective effect of Trillium tschonoskii saponin on CCl4‐induced acute liver injury of rats through apoptosis inhibition. Can J Physiol Pharmacol 94:1291‐1297.
Xia Y, Liu R, Chen R, Tian Q, Zeng K, Hu J, Liu X, Wang Q, Wang P,Wang XC, Wang JZ (2014) Novel multipotent AChEI‐CCB attenu‐ates hyperhomocysteinemia‐induced memory deficits and Neuropa‐thologies in rats. J Alzheimers Dis 42:1029‐1039.
Yan T, Yu X, Sun X, Meng D, Jia JM (2016) A new steroidal saponin,furotrilliumoside from Trillium tschonoskii inhibits lipopolysaccha‐ride‐induced inflammation in Raw264.7 cells by targeting PI3K/Akt,MARK and Nrf2/HO‐1 pathways. Fitoterapia 115:37‐45.
Zhang CE, Tian Q, Wei W, Peng JH, Liu GP, Zhou XW, Wang Q,Wang DW, Wang JZ (2008) Homocysteine induces tau phosphor‐ylation by inactivating protein phosphatase 2A in rat hippocampus.Neurobiol Aging 29:1654‐1665.
Zhu Q, Sun Y, Yun X, Ou Y, Zhang W, Li JX (2014) Antinociceptive effects of curcumin in a rat model of postoperative pain. Sci Rep 4:4932.
- 中国神经再生研究(英文版)的其它文章
- Weak phonation due to unknown injury of the corticobulbar tract in a patient with mild traumatic brain injury: a diffusion tensor tractography study
- Exosomes: a novel therapeutic target for Alzheimer’s disease?
- Inhibition of retinal ganglion cell apoptosis:regulation of mitochondrial function by PACAP
- Association between Alzheimer’s disease pathogenesis and early demyelination and oligodendrocyte dysfunction
- Nogo receptor expression in microglia/macrophages during experimental autoimmune encephalomyelitis progression
- Endothelial progenitor cell-conditioned medium promotes angiogenesis and is neuroprotective after spinal cord injury