
褐毛铁线莲花部结构的扫描电镜观察
2018-05-30 04:52:59郭艳平王竞红张永胜
江苏农业科学 2018年9期
王 非, 郭艳平, 王竞红, 张永胜, 李 强, 郑 瑞
(东北林业大学园林学院,黑龙江哈尔滨 150040)
毛茛科(Ranunculaceae)是一个遍布全球的大科,有59属2 500种[1],毛茛科植物大多数是一到多年生草本,少有灌木和藤本,如铁线莲属(Clematis)[2]。铁线莲属植物所包含的花朵类型丰富,花萼形状、颜色千变万化,其雌雄蕊结构的形态特征是系统分类学中的重要依据。我国毛茛科专家王文采通过对铁线莲属植物的深入研究,依据萼片由水平开展到向上直展和雄蕊从无毛到有毛的演化趋势,将我国现有铁线莲属植物划分为9个组[3]。在孢粉学方面,国内宋之琛等已对少数种进行了光学显微镜的描述[4-5],吴海燕运用扫描电子显微镜(SEM)对毛茛科部分植物花形态和柱头式样的发生和发育过程进行了观察研究,结果表明,在铁线莲属中,花萼与苞片具有同源性,铁线莲属植物中出现的多于4枚萼片的现象是次生的[6]。
褐毛铁线莲(Clematisfusca)是毛茛科铁线莲属的多年直立草本或藤本植物。在东北地区,花期为春末至秋初。褐毛铁线莲开花相对较早,花型别致,花量充盈且花期较长,开花期间同一株褐毛铁线莲上的花可接替绽放,可一直保持有花可赏,它为东北地区的园林垂直绿化增添了一种新的选择可能性。但是,对于褐毛铁线莲花部显微结构的研究至今未见报道。因此,本研究以露地栽培的褐毛铁线莲为植物材料,实地观察褐毛铁线莲的花部雌雄蕊形态特征,同时对花粉、雌蕊及萼片进行扫描电镜观察,以期为其今后的繁育栽培提供相应的理论知识,进一步为铁线莲属植物种质资源评价提供理论依据,为今后铁线莲属其他植物在园林绿化上的应用起到一定的指导作用,并为相关研究奠定一定基础[7]。
1 材料与方法
本试验材料为从吉林省磐石市烟筒山移植的野生褐毛铁线莲,该植株已在哈尔滨市成功露地越冬,且生长良好。试验地点设在黑龙江省哈尔滨市香坊区东北林业大学园林学院苗圃内,于2015年6月中旬在苗圃样地内采集褐毛铁线莲的新鲜花朵,之后立即制样,对其花粉、雌蕊及萼片进行扫描电镜观察。同时在样地内选取典型代表植株进行实地观察,确定雄蕊及柱头的形态特征。每个试验均重复3次。
1.1 花朵雄蕊及柱头形态特征的观察
在样地内选取具有代表性的若干褐毛铁线莲植株进行标记,在盛花期采用放大镜、镊子等工具进行雌雄蕊的基本形态观察。对柱头的伸长、颜色变化、有无黏液的分泌、雄蕊的伸长、散粉顺序等指标进行观察。
1.2 花粉扫描电镜的观察
采集盛花期的新鲜花粉,均匀撒在制样杯的双面胶上,在SCD-005型溅射仪上溅射,之后放入FEI QUANTA200型扫描电镜中扫描,最后观察、拍照。使用该电镜软件(精度 0.01 μm)对20粒花粉进行测量,包括最大值、最小值(表示变化幅度)和平均值,以及极轴和赤道轴长度。花粉粒大小以极轴长(P)和赤道轴长(E)的乘积(P×E)来表示,用极轴长和赤道轴长的比值(P/E)表示花粉粒的形状,P/E>1.6为长椭球形,P/E在1.3~1.6之间为椭球形[8]。拍摄花粉极面照片进行观察,再在高倍显微镜(4 000倍)下观察花粉纹理结构。
1.3 柱头、花萼片扫描电镜观察
于盛花期取处于不同时期的新鲜花朵,将其柱头和花柱粘在制样杯的双面胶上,3次重复,之后通过扫描电镜直接观察、拍照及测量。花萼片取具有典型代表性的花朵,截取花萼片的正反两面直接粘于样品台上,喷金,之后置于电镜下观察。
2 结果与分析
2.1 褐毛铁线莲花朵形态特征
褐毛铁线莲花苞呈紫色或红紫色(图1-a、图1-b),初开时,花苞先由4枚花萼镊合相交处绽开1个小口(图1-c),之后雌雄蕊随着花开口角度的不断扩大,向外进行伸长生长(图1-d)。雌蕊先露出,1 d内整个雌蕊即可全部露出(图1-e),刚绽放出来的柱头呈浅绿色且聚拢在一起(图1-f),之后变为黄白色,柱头逐渐呈指状分离,弯向雄蕊方向(图1-g),最后变为褐色萎蔫状(图1-h)。通常情况下,最初整个雌蕊高于周围的雄蕊,之后随着外轮雄蕊高度超过雌蕊,花药进行开裂,1~2 d内先开始进行散粉。随后内侧雄蕊高度逐渐与雌蕊群等高,或超过雌蕊,然后进行散粉。在之后的2~3 d内花药相继开裂并授粉。在散粉进行的同时,花丝也逐渐伸展、直立,之后向心发育,环绕于雌蕊群之上。雌蕊生长到一定长度后,离生心皮由聚拢状态转而分离,顶端柱头向外弯曲,最后其形态呈菊花状分散(图1-h)。柱头顶部发育成小指形态,以期最大程度地接收花粉。

2.2 花粉特征
2.2.1 花粉的形状与大小 在扫描电镜下,褐毛铁线莲的花粉为单粒花粉,整个形状为椭球形(图2-a),花粉的极面观为三裂圆形,赤道面观为纺锤形,表面具有3条萌发沟,是铁线莲属中常见的三沟型花粉(图2-b)。沟长平均为(24.29±1.95)μm,变化幅度为18.67~28.75 μm,长至近两极,但是3条萌发沟在极面处不相交,它们将花粉划分为3个部分,萌发孔不易被观察到。沟宽平均为(0.81±0.28)μm,变化幅度为 0.36~1.33 μm,在沟的两端不似沟中部向内侧深陷,而是趋于平缓呈钝圆形,沟沿部整齐不加厚(图2-a)。褐毛铁线莲的花粉极轴平均长度为(28.90±1.37)μm,变化范围为 26.67~32.5 μm。赤道轴平均长度为(20.98±2.28)μm,变化范围为18.67~29.33 μm。花粉粒大小约为28.90 μm×20.98 μm,花粉极轴与赤道轴长的比值平均为1.39±0.12。
2.2.2 花粉表面纹饰 在扫描电镜下,褐毛铁线莲的花粉外壁表面分布着细密均匀的刺状凸起,且大小不一,萌发沟内也密生点状突起,密集度大于花粉表面,花粉表面较为粗糙(图2-a)。

2.3 柱头与花柱
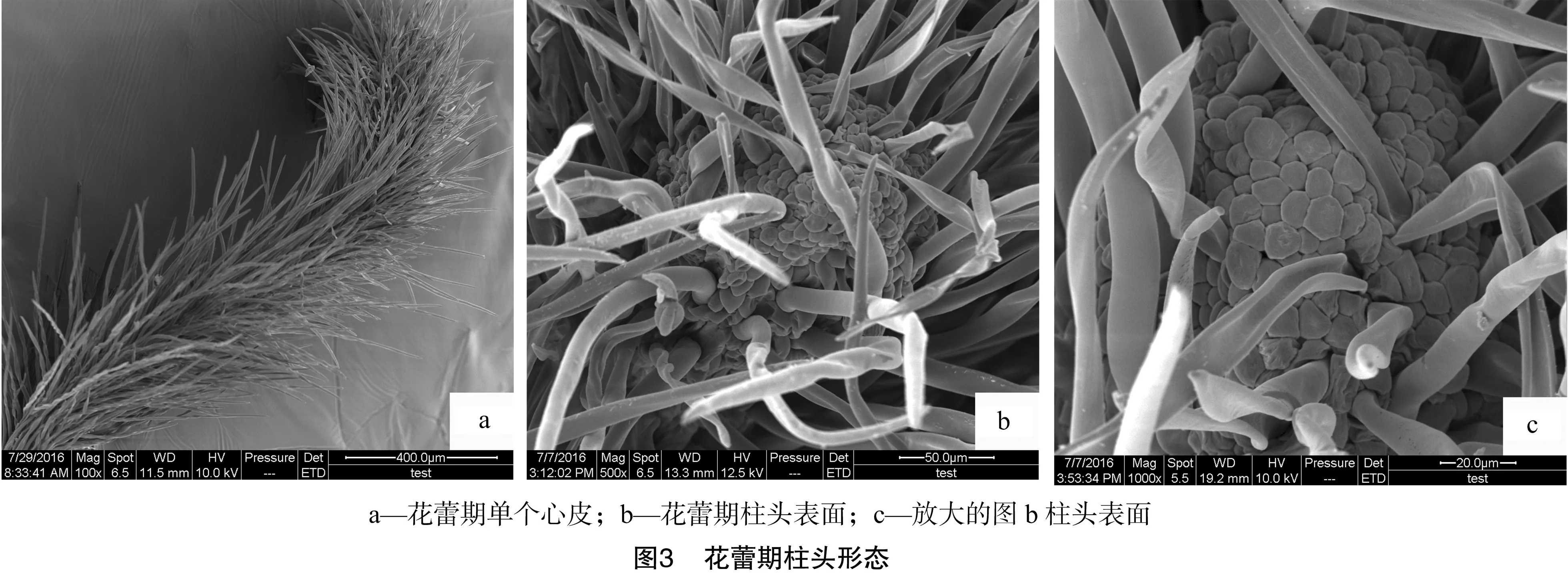
在扫描电镜下,褐毛铁线莲的柱头主要经历3个不同形态时期,分别为花蕾期、盛花期和末花期。在电镜下花柱整体为长圆柱形,顶端柱头呈尖圆形,后期弯向外侧。花柱及柱头上密生许多纤长的茸毛,长短不一,形成包围整个雌蕊的毛被,茸毛愈往顶部愈尖,有的茸毛底部扭曲,有的则直生(图 3-a、图3-b)。花柱顶部的茸毛(柱头部分)一般短于底部的茸毛,花柱内部的茸毛一般短于外围的茸毛。褐毛铁线莲受粉面较大,可由顶端沿着腹缝线向下延伸、渐次发育,最后呈长条带状分布于单个心皮向心的一侧。
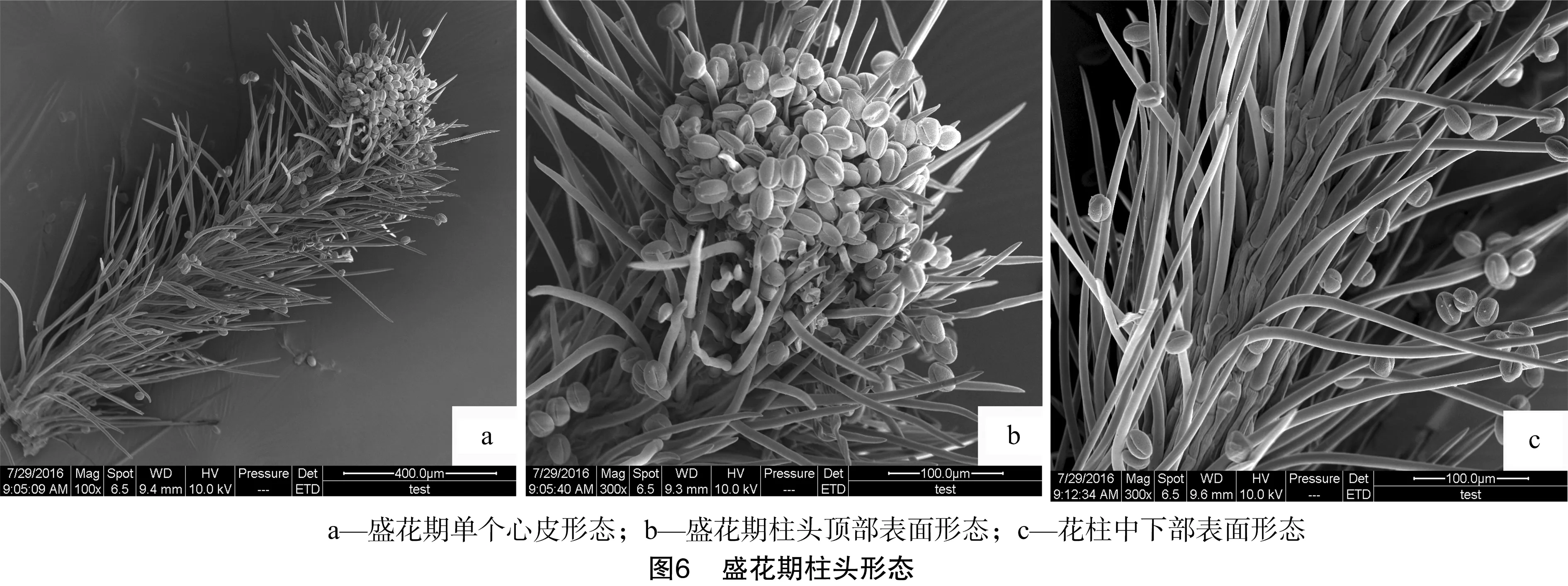
花蕾期柱头与花柱表面的细胞呈不规则形状(图3-c),在花蕾期花柱上已有乳突细胞形成(图3-b)。花刚开放时,柱头形态见图4,此时花苞先缓慢地绽出1个小口,雌蕊从内部随着花苞的绽放逐渐露出,这个时期在扫描电镜下的柱头与花柱仍未有花粉散落(图4-b)。随后花苞小口不断扩大,整个雌蕊结构全部露出,初花期到来,初花期柱头形态见图5。与此同时,外侧花药也开始散粉,先有少数花粉散落到柱头上,表明进入授粉状态。在盛花期,花粉主要聚集于整个花柱的顶部柱头上(图6-a),由顶部开始受粉面逐渐向中部延伸(图6-b)。花柱下部也有花粉散落(图6-c)。在末花期,柱头上仍有较多花粉,在扫描电镜下的柱头沿腹缝线向两侧张开,可明显看到中间部位的缝隙(图7),并且向花柱中下部延伸,受粉面逐渐延长扩大,花粉也随之向下分布。此时柱头已呈现萎蔫发黑的状态,但仍具备一定的受粉能力。
褐毛铁线莲的柱头主要集中在花柱顶部,受粉面由顶部逐次往下,这一点可由花粉的分布情况看出。褐毛铁线莲也存在腹缝线,由花柱顶部弯曲的一侧向中央位置及花柱下部开裂,但是由于褐毛的整个雌蕊结构密被的茸毛及较多的花粉,阻挡了该结构被发现,在扫描电镜下即使是最大的放大倍数也未能观察到该结构。但是,从花粉在柱头上的分布位置及扫描电镜下对末花期的整个花柱的观察中,可以清楚地得出褐毛铁线莲存在腹缝线结构的结论。



2.4 花萼片的扫描电镜观察结果
通过对花萼片的扫描电镜观察可知,电镜下萼片的茸毛及气孔器仅在背面分布,腹面相对较为光滑(图8-a、图8-b)。其背面的茸毛按照一定的方向进行排列,在具有明显脉序的地方分布尤为紧密(图8-b)。腹面的细胞呈不规则状,排列紧凑(图8-a)。气孔器则普遍存在于背面,其形状基本呈梭形,气孔缝周围由1圈厚实的类似唇形的细胞包围。气孔缝平均长度为18.19 μm,变化范围在11.11~28.89 μm;平均宽度为3.79 μm,变化范围在1.48~7.413 μm。气孔大小为18.19 μm×3.79 μm,平均厚度为4.72 μm,变化范围在2.78~7.41 μm。花萼片背面的两边缘处,分布着白色毡茸毛,肉眼可见其数量明显多于中央部位的茸毛,但是其长度短于中央部位的茸毛(图8-c)。
3 结论与讨论
对花形态特征的观察是深入认识植物繁殖器官生殖功能的首要条件[9],了解雌雄蕊的外观形态特征是观察花内部发生发育的重要途径。观察发现,褐毛铁线莲的雄蕊数量较多,雌蕊由多数离生心皮构成。在发育过程中,单个心皮的柱头顶端呈指状膨大,花柱及柱头均密生纤状茸毛。褐毛铁线莲柱头在成熟过程中,逐渐进行离心式发育,会随着4枚萼片开口角度的扩大,柱头逐渐伸长,达到一定程度后,柱头部分开始向外翻卷弯向雄蕊的方向,从外观上看,整个雌蕊部分呈菊花状。其雄蕊最初低于整个雌蕊结构,之后随着开花进程的推进,外轮雄蕊赶超过雌蕊,先进行第一轮的散粉,然后内侧的雄蕊逐渐成熟,进行接下来的逐次散粉,整个散粉过程呈阶段性进行。在实地观察中,用放大镜未观察到柱头表面有任何黏液类物质分泌,因此该柱头属于干柱头。干柱头在传粉时不产生分泌物,但柱头表面存在亲水性蛋白质薄膜,能从薄膜下角质层的断裂处吸收水分[10]。但是在其雄蕊基部有花蜜产生,且带有淡香味。通过对褐毛铁线莲雌雄蕊形态特征及发育过程的初步观察,可以为进一步的扫描电镜观察提供一定的形态依据,从而确定两者之间的相互关联性。
花粉是植物携带遗传信息的雄性生殖细胞,其形态具有较强的遗传保守性[11],是植物系统分类学、研究进化分析和物种起源的宝贵资源、是花部综合特征以及繁育系统研究中重要的组成部分[12-13]。褐毛铁线莲花粉的基本特征与吴海燕等研究的铁线莲属部分植物的花粉形态特征相似,均为三沟型花粉,极面观为三裂圆形,三孔沟,外壁密布刺状凸起[6,8],但是在花粉的大小、形状及凸起状态等方面存在差异。Santisuk研究了毛茛族124种植物的花粉,总结出毛茛族花粉有9种进化趋势,其中萌发孔形态的进化顺序是三沟—散沟—散孔;花粉外壁覆盖层是从平坦—轻度波状—强烈波状变化的[14]。由此可知,褐毛铁线莲的花粉属于较为原始的发育类型,该花粉表面较为粗糙,分布着均匀的刺状凸起,这样的结构符合虫媒花花粉的特征。因为花粉表面比较粗糙,易于昆虫足器等的黏附[15]。褐毛铁线莲花粉的形态特征有力地证明了它是以虫媒为主要传粉模式的类群。
植物为了接受花粉完成受精,进化出适应其花粉附着的柱头形态及受精机制[16]。柱头结构是花部构造中十分重要的组成部分,是接受花粉的场所。根据植物形态学的概念,构成雌蕊的单位是心皮。无论是合生心皮还是离生心皮,可以分化出下部能育的子房和上部不育的花柱,花柱上部扩展形成柱头[6]。褐毛铁线莲的雌蕊就是由多数离生心皮组成的,其柱头发育主要经历3个发育阶段,在花蕾期受粉面开始生成,在柱头向心面开始有乳突细胞出现,为受粉活动进行相关的准备,之后柱头的发育沿着腹缝线向两侧张开,进一步扩大受粉面。在末花期,可以清楚地看到腹缝线结构,同时可以观察到整个茸毛的形成在花蕾期就已基本完成,茸毛沿着整个雌蕊的生长变形而变化,从底部一直密生到柱头。初花期受粉开始进行,其受粉面的形成不是一次全部完成的,它是随着花期的推移,受粉面不断向花柱中部延伸,呈渐次发育的形式。这一点与吴海燕研究的铁线莲属6种植物的柱头发育过程[6]大体一致,这样的柱头发育方式,实际有效地延长了整个受粉时间,与此同时,外观表象上延长了花朵的观赏期。经比较后发现,褐毛铁线莲在顶端柱头上密生纤长茸毛,这与目前所观察到的其他种类的铁线莲存在一些差异,其他铁线莲的柱头顶端疏生或几乎无茸毛存在。而且褐毛铁线莲的花柱顶部(柱头部分)的茸毛一般短于底部的茸毛,花柱内部的茸毛一般短于外围的茸毛,这样的茸毛构造更加有利于花粉的捕捉和延长花粉在柱头上的滞留时间,在一定程度上为受粉的成功进行提供了保障。

经过实际观察得知,褐毛铁线莲的雄蕊群散粉时间不具有统一性,往往是雄蕊群外侧的花药先进行散粉,由外至内地进行阶段性散粉,这与柱头受粉面的渐次发育相适应,保证了受粉面与花粉的同步性。其花粉的分布总是在柱头及花柱向心的内侧分布较多,并且是从顶端开始逐渐向下分布,顶端的花粉数往往最多。受粉面的延伸使得花粉着落面增加,提供了更多的受粉机会。Cruden等对19种虫媒植物的研究表明,柱头面积的扩大提高了传粉效率[17]。并且雄蕊群内各个雄蕊的高度存在一定的差异,外层雄蕊高于内侧的雄蕊,这有效地对整个雌蕊结构形成了包围之势。雌蕊结构在发育时逐渐由聚拢到分散,受粉面随着分散的过程逐渐大面积地露出,这时内侧雄蕊的花粉可以落在延伸出来的受粉面上,参差错落的雄蕊结构更有利于受粉的进行,体现了其独特的捕获花粉的能力。
铁线莲属的植物没有花瓣结构的存在,往往是具有与花瓣相类似的萼片,其萼片较大且颜色鲜艳,呈花瓣状,行使花瓣的功能去吸引昆虫前来受粉。褐毛铁线莲的花萼属于典型的厚萼型,花蕾期萼片最初的颜色为绿色,之后逐渐变为紫色,花萼背面的茸毛也逐渐增多。前人对铁线莲属花萼片的研究多集中于其发生发育方面,关于其基本形态的研究报道则较少。因此本研究对褐毛铁线莲的花萼片进行了扫描电镜的形态观察,结果表明,在扫描电镜下成熟期的花萼背面遍布皮孔器和浓密的茸毛,背面的边缘被白色毡茸毛,腹面无皮孔和茸毛,表面比较光滑。背面相对粗糙,推测可能是便于昆虫的停留,利于更好地进行受粉,并且密实的茸毛结构在一定程度上降低了风速的流动,这样可以更好地减少水分的蒸腾。毛茛科植物中花瓣零星存在于不同类群中,一般而言,没有花瓣的属被认为是原始的[18]。由此可知,褐毛铁线莲萼片是相对比较原始的发育类型,保留了许多原始的特征。在实际观察中可知,褐毛铁线莲花朵口径较大,雌雄蕊暴露得也较为彻底,传粉者可以轻松地进行授粉活动。受粉面的延伸、雄蕊数量众多以及花萼的特殊结构为该种植物的繁殖提供了较为便利的条件。
:
[1]Tamura M. Ranunculaceae[M]// Kubitzki K,Rohwer J G,Bittrich V. The families and genera of vascular plants(Vol 2). New York:Springer-Verlag,1993:563-583.
[2]车晓芬. 毛茛科植物花分泌组织的比较形态学研究[D]. 西安:陕西师范大学,2012.
[3]史京华. 羽叶铁线莲杂交起源的初步研究[C]. 北京:中国科学院植物研究所,2003:5-6.
[4]宋之琛. 孢子花粉分析[M]. 北京:科学出版社,1995.
[5]中国科学院植物研究所形态室孢粉组.中国植物花粉形态[M]. 北京:科学出版社,1960.
[6]吴海燕. 毛茛科部分植物花器官发育及柱头式样对传粉的适应[D]. 兰州:西北师范大学,2008.
[7]及 利,张 洁,王会仁,等. 嫩江云杉造林特性及叶片形态特征研究[J]. 森林工程,2015,31(6):37-40.
[8]张彦妮,钱 灿. 12种百合属植物花粉形态扫描电镜观察[J]. 草业学报,2011,20(5):111-118.
[9]蔡正旺. 几种铁线莲属植物的花部综合征与访花昆虫多样性及甘青铁线莲传粉生物学研究[D]. 兰州:西北师范大学,2008.
[10]李耀龙. 连翘花粉、柱头、花柱和叶表皮的扫描电镜观察[J]. 宁夏师范学院学报,2010,31(3):48-52.
[11]丁春邦,李 强,李 燕,等. 重楼属9种5变种花粉活力与柱头可授性研究[J]. 草业学报,2009,18(4):61-66.
[12]赵先贵,肖 玲,毛富春. 中国植物花粉形态的研究进展[J]. 西北植物学报,1999,19(5):92-95.
[13]殷东生,魏晓慧. 濒危植物风箱果的研究现状及展望[J]. 森林工程,2016,32(2):19-22.
[14]Santisuk T A. Palynological study of the tribe Ranunculeae (Ranunculaceae)[J]. Opera Bot,1979,48:1-74.
[15]周凌瑜,刘群录,邵邻相. 虫媒花与风媒花花粉形态的比较[J]. 上海交通大学学报:农业科学版,2008,26(3):177-182.
[16]赵同欣,刘林德,张 莉,等. 南蛇藤花粉和柱头的生物学特性研究[J]. 植物研究,2014,34(3):309-316.
[17]Cruden R W,Miller-Ward S. Pollen-ovule ratios,pollen size,and the ratio of stigmatic area to the pollen-bearing area of the pollinator:a hypothesis[J]. Evolution,1981,35(5):964-974.
[18]Tamura M. Phylogenetical consideration of the Ranunculaceae[J]. Korean J Plant Taxon,1984,14:33-42.
猜你喜欢
西北植物学报(2023年9期)2023-10-17 13:34:00
经济林研究(2022年2期)2022-07-20 09:40:24
课外生活·趣知识(2022年6期)2022-07-14 09:50:42
现代农业科技(2022年4期)2022-03-08 11:04:12
园林科技(2021年3期)2022-01-19 03:17:32
中国果树(2020年2期)2020-07-25 02:14:16
数学小灵通(1-2年级)(2018年12期)2018-12-24 13:16:12
花卉(2017年7期)2017-11-15 08:53:30
红蜻蜓·低年级(2016年9期)2016-05-14 10:50:49
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06 07:49:00
