
辣椒疫霉菌诱导辣椒酵母双杂交cDNA文库的构建及鉴定
2018-05-29 00:29王玉姣陈姗姗孙柏华艾聪聪王中洋张修国
山东农业大学学报(自然科学版) 2018年3期
王玉姣,陈姗姗,孙柏华,艾聪聪,王中洋,张修国
辣椒疫霉菌诱导辣椒酵母双杂交cDNA文库的构建及鉴定
王玉姣,陈姗姗,孙柏华,艾聪聪,王中洋,张修国*
山东农业大学植物保护学院;山东省蔬菜病虫生物学省级重点实验室, 山东 泰安 271018
辣椒疫霉菌()引起的辣椒疫病是在世界范围内广泛分布的毁灭性病害之一,对我国辣椒生产造成严重的经济损失。为深入研究辣椒疫霉的致病机制,寻找辣椒疫霉在辣椒体内的互作蛋白,利用SMART同源重组交换技术,在酵母菌株Y187中构建了被辣椒疫霉侵染不同时期辣椒的cDNA文库。文库质量评定结果显示cDNA文库的滴度为1.13×106CFU/mL,文库的库容量为1.7×107CFU,文库的重组率为96%,插入片段平均长度在1Kbp左右。该文库的构建为研究辣椒疫霉的致病机制、辣椒的抗病机制及寻找药物靶标位点奠定基础。
辣椒疫霉菌; 辣椒; 酵母双杂交; cDNA文库
辣椒疫霉菌()引起的辣椒疫病是毁灭性土传病害,短期内暴发成灾[1],游动孢子主要侵染叶、果实和茎,引起软腐、倒伏,可减产50%以上,对蔬菜产业造成严重的经济损失。疫霉菌是一类营半活体营养的病原卵菌(Oomycetes)[2],能够在侵染过程中外泌效应因子到植物细胞中破坏植物的防卫反应促进自身侵染定殖[3]。效应因子是一类具有操控寄主先天免疫反应,加强病原菌在寄主内感染的蛋白质分子[4]。随着植物病原菌的深入研究,探索效应因子与寄主互作分子机制已经成为植物病理学家们研究的重要内容[4-6]。目前,越来越多的植物病原菌效应因子互作机制得到阐释,寄主中的互作靶标可以是核酸[7],也可以是单一的蛋白质或蛋白质复合物[8]。然而,至今还未见到对辣椒疫霉效应因子致病机制的相关报道。
酵母双杂系统是筛选互作蛋白最常用的方法[9],是利用酵母转录因子GAL4的特性来进行蛋白质相互作用分析的系统[10,11]。GAL4 DNA-BD(DNA结合域)与bait(诱饵)融合表达,GAL4 AD(转录激活域)与prey(猎物文库)融合表达。Bait与prey之间的相互作用可以使GAL4重新具有转录活性,从而可以激活下游报告基因的表达。利用选择培养基进行筛选即可获得具有报告基因活性的克隆,从而分析蛋白质之间的相互作用[12]。
筛选辣椒疫霉效应因子的宿主靶标蛋白,研究该蛋白在病菌侵染过程中如何调控植物的防卫反应,可采用构建辣椒cDNA文库筛选互作蛋白而实现。本研究采用SMART(Switching Mechanism at 5’End of the RNA Transcript)cDNA合成技术,通过同源重组方法构建了被辣椒疫霉侵染0 h、12 h、24 h、48 h、60 h及72 h的辣椒cDNA文库,为筛选辣椒疫霉效应因子互作蛋白,深入研究效应因子与宿主互作分子机制奠定基础。
1 材料与方法
1.1 材料来源
辣椒疫霉菌株SD33由本实验室分离鉴定,辣椒为本实验室种植的自交系06221,Make Your Own“Mate & PlateTM”Library System User Manual、cDNA文库构建试剂盒及SD/-Leu营养选择性培养基均购自Clontech公司,DH5α感受态细胞购自北京全式金生物技术有限公司。
1.2 文库鉴定引物
根据pGADT7-Rec序列设计1对扩增文库插入片段的鉴定引物。引物序列为:F:5′-TAATACGACTCACTATAGGGCGAGCGCCGCCATG-3′,R:5′-GTGAACTTGCGGGGTTTTTCA GTATCTACGATT-3′,引物由济南铂尚生物公司合成。
1.3 总RNA的提取
用液氮研磨被辣椒疫霉侵染0 h、12 h、24 h、48 h、60 h及72 h的辣椒叶片,采用Trizol法提取总RNA。用1% 的琼脂糖凝胶电泳检测提取的RNA质量。
1.4 cDNA文库的构建
1.4.1 双链cDNA的合成以5 μg总RNA为模版,以CDS Ⅲ(Oligo-dT)为引物,SMART MMLV Reverse Transcriptase反转录合成第一链cDNA;以第一链cDNA为模版,使用LD PCR(Long distance PCR)扩增合成dscDNA。利用CHROMA SPIN+TE-400 Columns分级分离纯化双链cDNA,在1.2%的琼脂糖凝胶上对PCR产物进行分析。
1.4.2 Y187菌株感受态细胞的制备挑取活化的Y187单克隆菌落,在YPDA液体培养基中振荡培养至OD600达到0.15~0.3;3800 rpm离心5 min,弃上清,加入100 mLYPDA重悬菌液并继续培养至OD600达到0.4~0.5,3800 rpm离心5 min,弃上清,无菌水重悬菌体后再次离心弃上清;加入1.5 mL1.1×TE/LiAc溶液重悬菌体高速离心15 s,弃上清,使用600 μL1.1×TE/LiAc重悬菌体,即为感受态细胞。
1.4.3 文库的构建和收集向500 µL Y187感受态中加入:6 µL pGADT7-Rec Cloning Vector,5 µg cDNA,3 µL SV40 Large-T PCR fragment (25 ng/µL),20 µL Herring Tests Carrier DNA,2.5 mL PEG/LiAc轻轻混匀;30 ℃孵育45 min,每15 min轻轻混匀1次;然后加入160 µL DMSO,42 ℃孵育20 min,每10 min混匀1次;3800 rpm离心5 min弃上清,加入3 mL YPD Plus重悬菌体,30 ℃振荡培养90 min;相同条件再次离心收集菌体,用15 mL 0.9% NaCl重悬菌体。将菌液涂布在15 cm SD/-Leu选择培养基上。每个板涂布150 µL菌液,30 ℃倒置培养3~4 d。文库收集:每个平板使用3 mL冻存液刮洗,将所有菌液混匀于无菌锥形瓶。将文库分装10管于1.5 mL离心管每管1 mL,剩余菌液分装于50 mL离心管,-80 ℃贮存。
1.4.4 文库的质量鉴定取500 µL共转化菌液按101到104稀释,分别涂布在SD/-Leu 15 cm固体培养基上,每个梯度涂布3个平板,30 ℃,倒置培养3~4 d。对生长的单菌落计数,计算文库转化效率和文库容量,文库滴度=平板上菌落数×稀释倍数/涂板体积。于SD/-Leu平板上随机挑取50个克隆进行PCR鉴定,确定文库的阳性率及插入片段大小的分布。
2 结果与分析
2.1 总 RNA的提取
提取被辣椒疫霉侵染不同时期的辣椒叶片总RNA,使用1%琼脂糖凝胶检测总RNA质量,如图1所示,28S和18S rRNA条带清晰,表明RNA完整性较好;经测定分析,A260/A280为1.98,浓度约为1.5 µg/µL,这表明RNA提取质量较好可用于下一步实验。
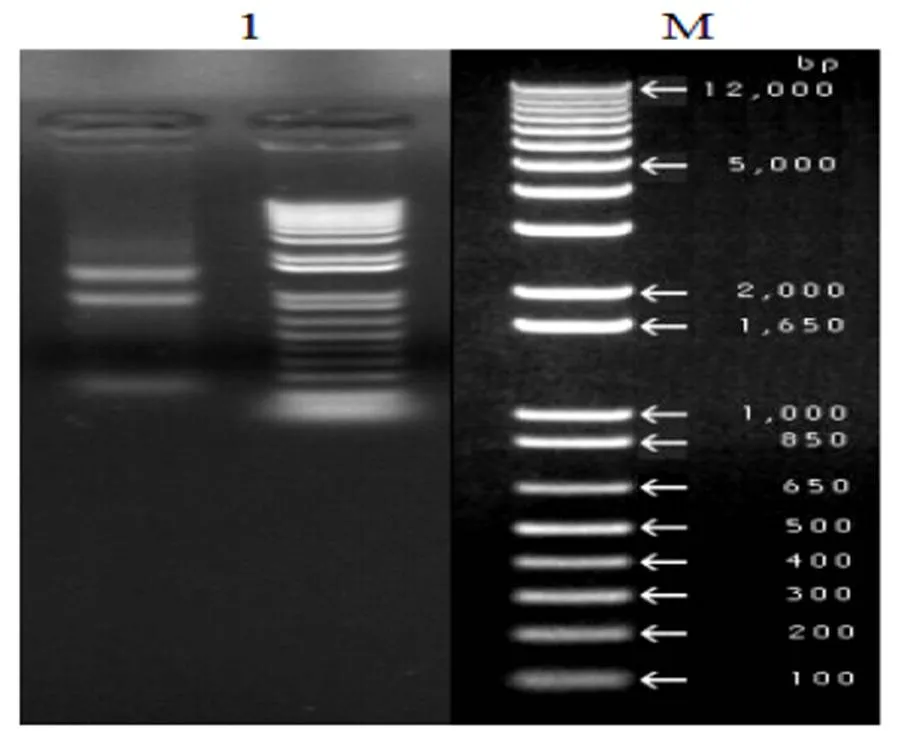
1:辣椒总RNA; M:DNA Marker DL12000
Fig.1 Detection of pepper total RNA by electrophoresis

1: dscDNA; M:DNA Marker DL12000
2.2 双链cDNA文库的琼脂糖凝胶电泳
以总RNA为模板,CDS Ⅲ为引物反转录合成第一链cDNA;以第一链cDNA为模板,使用LD PCR扩增合成 dscDNA,合成的dscDNA经1.2%琼脂糖凝胶电泳检测,如图2所示,大小集中分布在600~2000 bp范围内,且该区域有弥散状条带出现,表明双链cDNA在该区域的分布较为完整,丰度较好。
2.3 酵母双杂交cDNA文库的构建和评价
Mate & Plate文库构建是在酵母中进行的,借助生物体自身的同源重组功能将cDNA克隆到pGADT7-Rec中。首先,使用SMART cDNA合成技术合成含有与pGADT7-Rec载体末端同源的cDNA,然后,将cDNA和pGADT7-Rec转入Y187中,然后在SD/–Leu固体培养基上培养3~5 d。最后将所有克隆收集并分装成1 mL的文库用于双杂交筛选。
统计稀释10、102倍、103倍和104倍后长出的单克隆数目,求平均值后计算文库滴度,文库滴度=每板平均克隆数/涂布体积×稀释倍数(表1);总库容量Total CFU (cfu) =平均滴度(cfu/mL)×cDNA library总体积(mL)。该文库平均滴度为1.13×106CFU/mL,共计15 mL的转化后原始菌液,则总库容量为:1.13×15×106CFU=1.7×107CFU。从SD/-Leu平板上随机挑25个克隆进行摇菌提质粒后进行PCR检测,部分结果如图3所示,显示插入片段的大小集中在600~2000 bp之间,阳性菌落数为24个,重组率为96%。

表 1 文库滴度

M: DNA Marker DL2000; 1-24: 插入片段PCR产物
3 讨论
大量的研究表明,辣椒疫霉能够在侵染过程中外泌效应因子进入植物细胞中破坏植物的防卫反应促进自身侵染定殖[13,14],效应因子通过与宿主靶标蛋白互作,调控植物的信号通路及代谢活动从而实现自己的功能。因此,为了更好的控制由辣椒疫霉引起的辣椒疫病,我们需要对效应分子的靶标蛋白以及效应分子的致病机制进行更加深入的了解。
对基因功能的研究离不开对基因所编码蛋白的研究,蛋白质是基因功能的执行者和体现者,生命活动中大多数蛋白质需与其他蛋白结合才能发挥作用。目前,酵母双杂交是研究蛋白质互作最广泛的技术[10],具有筛选效率较高、可以检测到一些微弱的蛋白质互作以及在酵母细胞内验证互作而无需纯化蛋白质等优点,成为当前互作蛋白筛选的主要方法[8]。
本研究利用SMART技术合成dscDNA建立酵母双杂交文库,无需对样本总RNA中mRNA分离纯化,有效避免了纯化过程中mRNA的降解;重组cDNA片段集中在600-2000 bp之间,具有较好的多态性,满足后续要求;利用同源重组方法,将外源dscDNA定向与pGADT7-Rec载体重组,避免了繁琐的酶切连接等步骤和解决了低效率连接和克隆问题,从而保证了cDNA文库的完整性。本研究以多达7个时间点的经辣椒疫霉诱导的辣椒叶片作为建库材料,保证了文库包含的抗病相关基因数量的最大化。本研究所在课题组前期已构建部分效应因子的诱饵载体,可使用本文库进行效应因子互作蛋白筛选工作。虽然cDNA文库中辣椒基因在各种信号转导和代谢通路已有注释,但其在辣椒体内的实际功能还有待发掘。因此,本研究构建的经辣椒疫霉SD33诱导的辣椒酵母双杂交cDNA文库,不仅有助于辣椒基因功能注释,还将推动抗病基因的发掘和抗病机制的解析。
4 结论
本研究利用SMART技术构建了经辣椒疫霉SD33诱导的辣椒酵母双杂交cDNA文库。提取的总RNA条带清晰,未发生降解,并且没有蛋白质和其他小分子物质的污染;文库容量为1.7×107,滴度为1.13×106cfu/mL,重组cDNA片段大小集中在600~2000 bp之间,重组率96%。该文库质量较高,满足下步互作蛋白筛选及致病机制分析。
[1] 杨楠,张丽,丁鹏,等.辣椒疫霉效应分子RxLR121504功能特性的研究[J].菌物学报,2017,36(9):1233-1242
[2] Ah Fong AM, Judelson HS. New role for Cdc14 phosphatase: localization to basal bodies in the oomyceteand its evolutionary coinheritance with eukaryotic flagella[J]. PLoS ONE, 2011,6:e16725
[3] Yu Du, Mohamed HM, Paul RJB,.RXLR effector AVR1 interacts with exocyst component Sec5 to manipulate plant immunity [J]. Plant Physiology, 2015, 1975-1990.
[4] Goritschnig S, Krasileva KV, Dahlbeck D,. Computational prediction and molecular characterization of an oomycete effector and the cognateresistance gene[J].PLoS Genet, 2012,8(2):el002502
[5] King SR, McLellan H, Boevink PC,RxLR effector PexRD2 interacts with host MAPKKKε to suppress plant immune signaling [J]. Plant Cell2014,26(3):1345-59
[6] 李金玥.植物病原菌效应因子的毒性功能研究进展[J].科教导刊,2016(4):25-27
[7] Diane GO, Susan B, Joe W,. Host protein BSL1 associates withRxLR effector Avr2 and theimmune receptor R2 to mediate disease resistance [J]. Plant Cell, 2012, 24:3420-3434
[8] Auerbach D, Stagljar I. Yeast two-hybrid protein-protein interaction networks [M]//Proteomics and protein-protein interactions:biology, chemistry, bioinformatics, and drug design. Singapore:Springer Science & Business Media, Inc, 2005:19-31
[9] 杨君,张艳,王伟巧,等.黄萎病菌诱导海岛棉酵母双杂交cDNA文库构建及评价[J].生物技术通报,2014,29(12):105-110
[10] Bram S, Hélène T, Jan T,. Diversity in Genetic In Vivo Methods for Protein-Protein Interaction Studies: from the Yeast Two-Hybrid System to the Mammalian Split-Luciferase System Microbiol[J]. Mol. Biol. Rev. 2012,76:2331-382
[11] 赵慧,王遂,姜静,等.酵母双杂交筛选与小黑杨PsnWRKY70相互作用的蛋白质[J].北京林业大学学报,2016,38(2):44-51
[12] Oender K, Niedermayr P, Hintner H,. Relative quantitation of protein-protein interaction strength within the yeast two-hybrid system via fluorescence beta-galactosidase activity detection in a high-throughput and low-cost manner[J]. Assay and Drug Development Technologies, 2006,4(6):709-719
[13] Erbs G, Newman MA. The role of lipopolysaccharide and peptidoglycan, two glycosylated bacterial microbe-associated molecular patterns (MAMPs), in plant innate immunity[J].MolecuLar Plant Pathology, 2012,13(1):95-104
[14] Dou D, Kale SD, Wang X,. RXLR-mediated entry ofeffector Avr1b into soybean cell does not require pathogen-encoded machinery [J]. Plant Cell, 2008,20:1930-1947
Construction and Characterization of Yeast Two-Hybrid cDNA Library Derived fromInoculated with
WANG Yu-jiao,CHEN Shan-shan, SUN Bai-hua, AI Cong-cong, WANG Zhong-yang, ZHANG Xiu-guo*
271018,
is the casual agent ofPepper blight, which is one of the most destructive diseases that harms pepper industry around the world.Therefore, to study the pathogenic mechanism and screen proteins for, yeast two-hybrid cDNA libraries of pepper infected bywere constructed in yeast strain Y187 using homologous recombination-mediated SMART technology. The qualities of the cDNA libraries were evaluated as the followings: the titers were 1.13×106CFU/mL, the capacity of cDNA library was 1.7×107CFU/mL, the recombinant rate was 96%, and the protein average length encoded by the inserted cDNA is about 1Kbp. The construction of this library lays the foundation for studying the pathogenic mechanism of, the disease resistance mechanism of pepper and the search for drug target sites.
;; yeast two-hybrid; cDNA libraries
S436.418.1+2
A
1000-2324(2018)03-0379-04
2017-09-23
2017-10-23
国家大宗蔬菜产业技术体系(CARS-25-03B)
王玉姣(1990-),女,在读硕士研究生,研究方向:植物病原卵菌分子遗传学. E-mail:18354285903@163.com
Author for correspondence. E-mail:zhxg@sdau.edu.cn
猜你喜欢
当代水产(2022年1期)2022-04-26
昆明医科大学学报(2022年2期)2022-03-29
中国农业科学(2021年23期)2022-01-14
华侨大学学报(自然科学版)(2021年4期)2021-07-30
猪业科学(2021年3期)2021-05-21
中国医药科学(2021年7期)2021-05-11
幽默大师(2020年10期)2020-11-10
中国瓜菜(2020年8期)2020-09-26
中华诗词(2019年1期)2019-11-14
猪业科学(2018年4期)2018-05-19
