
BMR基因型高粱不育系主要农艺性状的配合力效应
2018-05-28 01:54:50游永亮赵海明武瑞鑫刘贵波
草业科学 2018年5期
李 源,游永亮,赵海明,武瑞鑫,刘贵波
(河北省农林科学院旱作农业研究所 河北省农作物抗旱研究重点实验室,河北 衡水 053000)
高丹草(Sorghumbicolor×S.sudanense)是高粱(S.bicolor)与苏丹草(S.sudanense)的远缘杂交种,其选育技术借鉴的是普通高粱杂交种“三系”配套原理,母本主要利用高粱不育系(或将普通高粱不育系改良为饲草型),父本将高粱恢复系改为苏丹草。近年来,高丹草作为一种优质饲草,受到了国内外学者的普遍关注[1-3],但花后期青贮利用时,因木质化程度增强,导致饲用品质下降,给生产利用带来一定困难[4-5]。
褐色中脉(brown midrib,BMR)基因型突变体是指植株叶片中脉、茎表皮以及髓部有褐色,是根据其表型命名的;一般在植株生长到6个扩展叶后开始表现,花期后随着叶片老化褐色逐渐消失[6]。研究表明,具有这种基因型的材料能使茎、叶中难以消化的木质素含量降低40%左右,能明显提高消化率[7-9]。因此,培育BMR基因型高丹草新品种,将会大幅度改善高丹草的饲用品质,应用前景广阔。
BMR基因型高丹草新品种的选育途径同普通高丹草,只是将高粱不育系转育成BMR基因型高粱不育系,苏丹草改良成BMR基因型苏丹草,然后再进行远缘杂交,核心是选择杂种优势强大的组合,而选择组合的关键在于双亲配合力的大小[10]。关于BMR基因型高丹草配合力的研究较少,但在高粱配合力的报道上较多,张晓娟等[11]对3个新选高粱不育系与7个恢复系组配的21个杂交组合的配合力分析得出,在杂交种选配时,首先应考虑双亲穗粒重的配合力,其次考虑双亲株高的配合力;王立新等[12]对5个高粱不育系和5个恢复系组配的25个杂交组合的配合力分析得出,产量性状、千粒重、茎粗、穗粒重这些性状应选择一般配合力高的亲本进行轮回选择和直接利用;李巍等[13]对5个甜高粱不育系和5个甜高粱恢复系组配25个杂交组合的配合力分析得出,产量和糖锤度一般配合力较高,有较大的利用价值;吕鑫等[14]对5个高粱不育系和6个饲草高粱恢复系组配的30个杂交组合的配合力分析得出,生物产量、株高、茎粗、分蘖和干重5个农艺性状的遗传是受加性和非加性效应双重影响,其中加性效应占主导地位;尹学伟等[15]对6个糯高粱不育系和8个糯高粱恢复系组配的48个杂交组合的配合力分析得出,抽穗期、穗粒重、千粒重、育性等性状受加性基因效应控制,株高、穗长、穗粒数和产量性状受非加性基因影响较大。上述有关高粱配合力的研究多采用不完全双列杂交设计,主要集中在普通粒用高粱、甜高粱、饲草高粱以及糯高粱上,但所选材料亲本的数量、农艺性状指标也各不相同,得出的结果也不尽一致;可以看出,在高粱配合力大小的评价方法未见统一标准,而有关本研究新引及自育的19份BMR基因型高粱不育系配合力的大小也未见报道。基于此,本研究采用不完全双列杂交试验设计,以19份引进和自育的BMR基因型高粱不育系为母本(待测系),以自主选育的3份BMR基因型苏丹草为父本(测验种),从与饲草产量相关的株高、主茎叶片数、单株分蘖数、主茎直径、中部叶片长度、中部叶片宽度、主茎穗长共7个农艺性状上对其组配的57个F1杂交组合进行配合力效应分析,旨在探讨这些农艺性状在杂交后代的表现规律,挖掘配合力强的BMR基因型高粱不育系和杂交组合;为杂交育种双亲选择、杂交组合F1代田间评价遴选标准的确定提供理论依据。
1 材料与方法
1.1 试验材料
试验亲本材料共22份,分为BMR基因型高粱不育系、BMR基因型苏丹草两组。BMR基因型高粱不育系共19份,其中15份由美国引进,分别为T98A、AMP12、AMP15、AMP17、AMP19、AMP21、AMP447、AMP448、AMP449、AMP450、AMP451、AMP452、AMP453、AMP454、AMP455;其余4份由河北省农林科学院旱作农业研究所采用连续回交法选育而成,分别为H33A、H41A、H117A、H129A。BMR基因型苏丹草共3份:ZS-29、ZS-102、15C-ZY487,由河北省农林科学院旱作农业研究所采用系谱法选育而成。BMR基因型高粱不育系套袋自交是不结实的,其繁殖方式是在隔离条件下通过其保持系授粉来繁殖不育系;苏丹草为常异花授粉作物,BMR基因型苏丹草繁殖方式同普通苏丹草,是在隔离条件下开放授粉获得后代。供试材料名称、来源以及主要农艺性状的基本信息如表1所列。
1.2 试验设计
试验按NCⅡ不完全双列杂交设计,以19份BMR基因型高粱不育系为母本(待测系),以3份BMR基因型苏丹草为父本(测验种),于2015年冬在河北省农林科学院海南试验站(海南三亚,109° E,18° N)人工组配杂交组合:以待测的每个BMR基因型高粱不育系分别与3个不同的BMR基因型苏丹草进行组间杂交,即每个不育系于抽穗后开花前,先用硫酸纸袋套5~7个单株主茎穗,待不育系开花后再分别取不同的父本花粉进行授粉,每隔2 d授粉一次,共配置F1组合57个。
2016年5月23日,分别将这22个亲本材料、57个杂交组合以及对照品种皖草2号一起种植在河北省农林科学院旱作农业节水试验站(河北衡水,115° E, 37° N)。采用随机区组排列,3次重复,每份材料播种2行,行长6.0 m,行距40 cm,株距15 cm,播种深度3~5 cm。试验条播种植,造墒播种,灌水量为600 m3·hm-2,播前底施复合肥750 kg·hm-2,播后苗前采用38%莠去津(阿特拉津,Atrazine)悬浮剂均匀喷施地表防除杂草,用药量为1 800~2 250 g·hm-2。
1.3 指标测定
当每小区50%以上植株达到乳熟期时,在每小区分别选择长势整齐一致的10个单株,测量株高、主茎直径、主茎穗长等农艺性状指标。株高:测量从地面到穗(或植株新叶)顶部的绝对高度;单株分蘖数:统计每个单株的分蘖数;主茎叶片数:统计每个单株主茎的叶片数;主茎直径:用游标卡尺测量距地面30 cm处主节间的直径;用直尺测量每个单株中间部位叶片从叶舌到叶尖的最长距离以及叶片的最宽距离,分别记为中部叶片长度、中部叶片宽度;同时用直尺测量主茎穗的长度,记为主茎穗长;开花天数:统计每份材料从播种日到50%以上植株达到初花期的天数;穗部性状的调查分穗型、穗形、壳色、粒色,穗型主要分圆筒型、纺锤型、圆锥型等,穗形分散、中散、紧、中紧等,壳色、粒色主要有黑、白、黄、黑红色等。
1.4 数据处理
运用Excel 2007软件对不同性状下的数据进行平均值计算,杂种优势计算公式如下[16]:
中亲优势(MPH)=(F1-MP)/MP×100%。
式中:MP为双亲的中亲值,MP=(P1+P2)/2;P1、P2分别为BMR基因型苏丹草、BMR基因型高粱不育系主要农艺性状表现值。
超亲优势(HPH)=(F1-HP)/HP×100%。
式中:HP为高值亲本表现值,其中主茎叶片数、主茎直径、中部叶片长度、中部叶片宽度、主茎穗长为BMR基因型高粱不育系表现值,株高、分蘖数为BMR基因型苏丹草表现值。
竞争优势(CKH)=(F1-CK)/CK×100%。
式中:CK为对照皖草2号主要农艺性状的表现值。
利用DPS 3.01专业版软件进行配合力方差统计分析;利用不完全双列杂交设计模块计算一般配合力(GCA)、特殊配合力(SCA)及配合力效应[17]。
2 结果与分析
2.1 各组合主要农艺性状杂种优势分析
2.1.1生育期及穗部性状调查分析 对19份母本BMR基因型高粱不育系、3份父本BMR基因型苏丹草以及组配的57个F1杂交组合的生育期及穗部性状分析得出(表2),不同杂交组合下的开花天数明显长于父、母本的开花天数,生育期偏向晚熟;F1杂交组合的穗型偏向于父本、母本的中间型,多以纺锤型为主;穗形多以散、中散为主,偏向父本穗形;壳色多以黑壳、红壳为主,偏向为父本颜色;粒色多偏向于双亲的中间型,多以黄色、白色、黑红色为主。
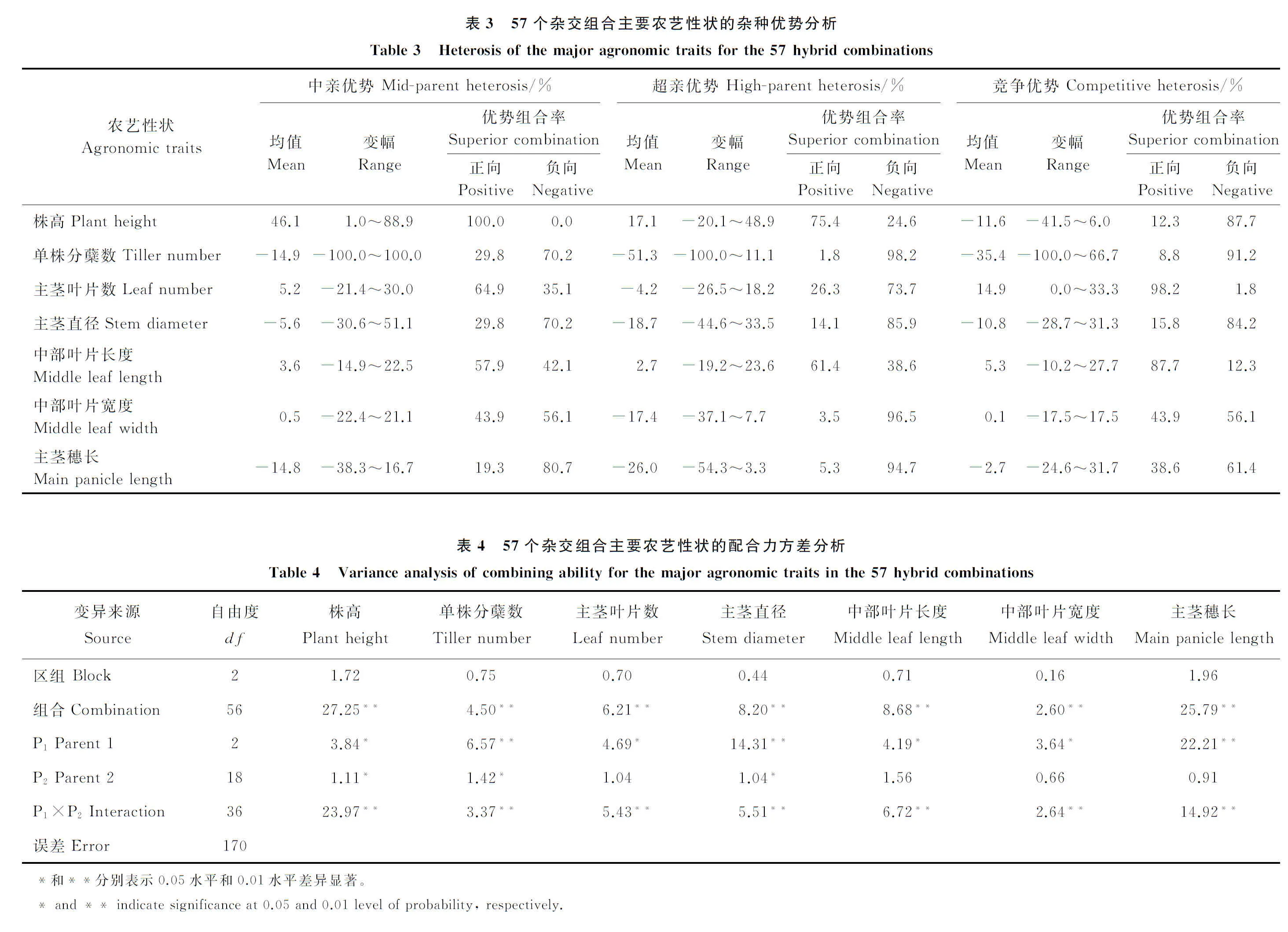
2.1.2农艺性状杂种优势分析 57个F1杂交组合相同性状不同杂交组合下的中亲优势、超亲优势、竞争优势变幅较大(表3),表明不同杂交组合在这些性状上的表现差异明显。对中亲优势分析得出,株高、主茎叶片数、中部叶片长度、中部叶片宽度4个性状具有正向中亲优势,以株高的正向优势组合率最大,为100%;单株分蘖数、主茎直径、主茎穗长表现为负向中亲优势,以主茎穗长的负向优势组合率最大,为80.7%。对超亲优势分析得出,株高、叶片中部长度表现为正向超亲优势,以株高的正向超亲优势组合率最大,为75.4%;单株分蘖数、主茎叶片数、主茎直径、中部叶片宽度、主茎穗长表现为负向超亲优势,以单株分蘖数、主茎直径、主茎穗长的负向超亲优势较高,分别为-51.3%、-18.7%、-26.0%,但这些性状下也具有一定的正向组合率,分别为1.8%、14.1%、5.3%。对竞争优势分析得出,主茎叶片数、中部叶片长度、中部叶片宽度表现为正向竞争优势,且变幅较大,以主茎叶片数的正向竞争优势最大,达到98.2%。而株高、单株分蘖数、主茎直径、主茎穗长表现为负向竞争优势。
2.2 各性状配合力效应分析
2.2.1配合力的方差分析 在进行配合力分析之前,首先需要对各性状的配合力进行方差分析,只有基因型引起的方差表现为显著时,才能进行配合力效应分析;反之,则不需要进行配合力效应分析[14]。本研究中,配合力方差分析表明(表4),57个杂交组合的7个主要农艺性状在区组间差异均不显著(P>0.05),在组合间呈极显著差异(P<0.01),说明不完全双列杂交的57个组合在7个农艺性状上存在真实性的遗传差异,这种遗传的差异性是加性基因和非加性基因共同作用的结果[12]。因此,有必要将组合间方差分解成父本P1(BMR基因型苏丹草)、母本P2(BMR基因型高粱不育系)和父母本互作(P1×P2)的配合力进行方差分析。在P1(BMR基因型苏丹草)中,不同亲本间各性状的一般配合力差异显著(P<0.05),其中单株分蘖数、主茎直径、主茎穗长呈极显著差异(P<0.01);在P2(BMR基因型高粱不育系)中,株高、单株分蘖数、主茎直径性状呈显著差异(P<0.05),而主茎叶片数、中部叶片长度、中部叶片宽度、主茎穗长性状差异不显著(P>0.05);父母本互作(P1×P2)条件下,各性状的特殊配合力呈极显著差异(P<0.01)。

2.2.2一般配合力效应分析 相同BMR基因型高粱不育系在不同农艺性状间的一般配合为(GCA)不同,相同农艺性状不同不育系间的GCA差异也较大(表5)。株高的GCA值以T98A、AMP21、H129A表现较高,单株分蘖数GCA值以AMP12、T98A、AMP19表现较高,主茎叶片数GCA值以AMP19、AMP454、AMP455和H117A表现较高,主茎直径GCA值以AMP454、AMP453、AMP448表现较高,中部叶片长度GCA值以H129A、AMP15、AMP454表现较高,中部叶片宽GCA值以AMP452、AMP453、T98A表现较高,主茎穗长GCA值以AMP454、AMP455、AMP19表现较高。
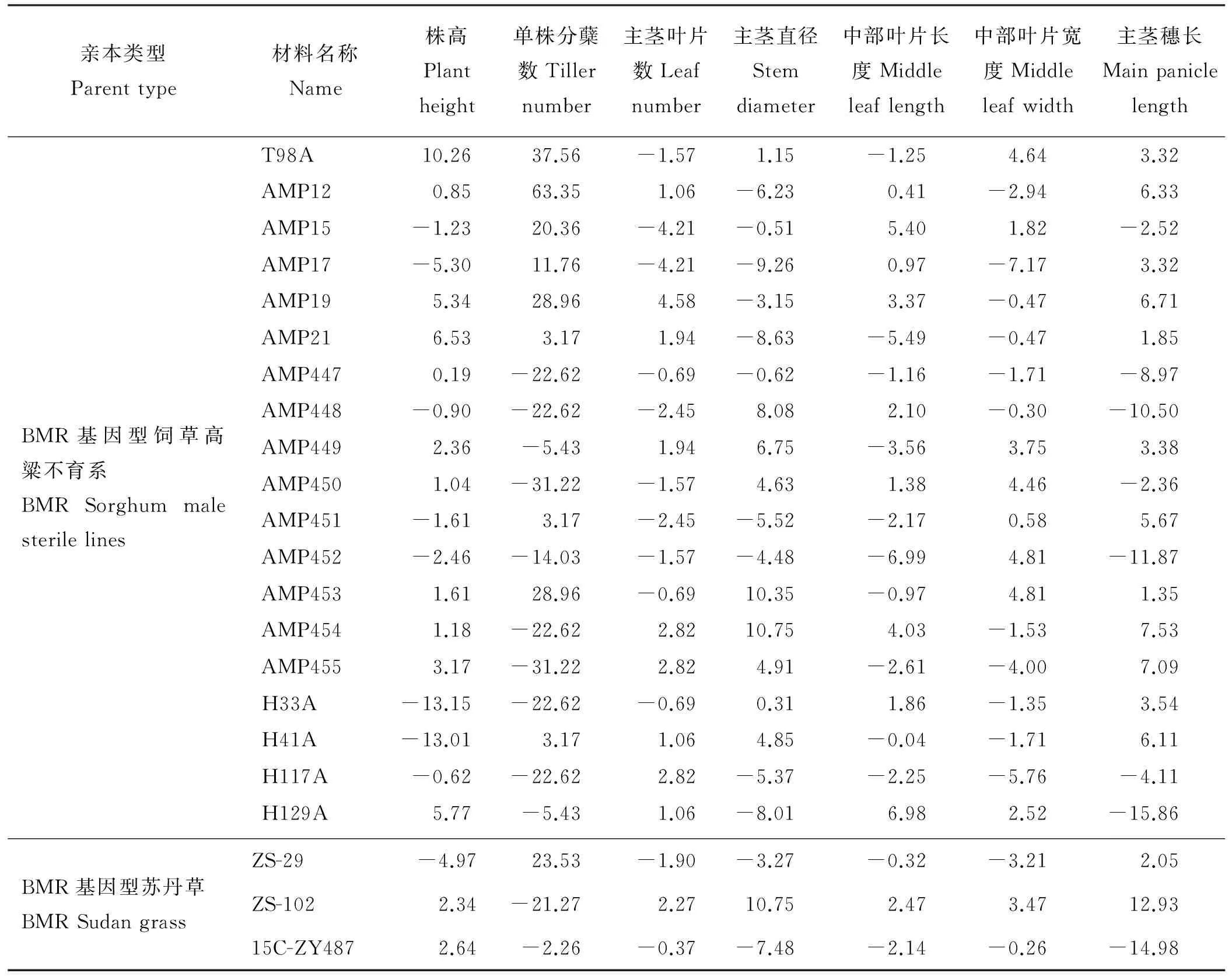
表5 BMR基因型高粱不育系、BMR基因型苏丹草主要农艺性状的一般配合力(GCA)效应值Table 5 General combining ability for the major agronomic characters of BMR sorghum male sterile lines and BMR sudangrass
BMR基因型苏丹草的GCA在不同农艺性状间也存在一定程度变异(表5),其中ZS-102的农艺性状除单株分蘖数外,GCA值均表现出较高正值,表明其一般配合力较高,而15C-ZY487的农艺性状除株高外,GCA值均表现出较高的负值。
2.2.3特殊配合力效应分析 相同杂交组合不同农艺性状间的特殊配合力(SCA)不同,相同农艺性状不同杂交组合的特殊配合力差异较大(表6)。在株高上,SCA值表现较大的组合为H41A×15C-ZY487、H33A×15C-ZY487、AMP454×ZS-102;在单株分蘖数上,SCA值表现较大的组合为AMP12×ZS-29、AMP19×ZS-29、AMP21×15C-ZY487;在主茎叶片数上,SCA值表现较大的组合为AMP19×ZS-102、AMP454×ZS-102、AMP17×ZS-102;在主茎直径上,SCA值表现较大的组合为AMP454×ZS-102、H33A×ZS-102、H117A×ZS-29;在叶片中部长度和宽度上,以AMP454×ZS-102、AMP447×ZS-29、AMP450×15C-ZY487的SCA值较高;在主茎穗长上,以AMP21×15C-ZY487、H33A×ZS-102、AMP449×ZS-29的SCA值较大。
2.3 各性状遗传参数分析
遗传参数分析得出(表7),除中部叶片宽度外,其他6个农艺性状总亲本的遗传方差均大于环境方差,说明遗传因素是这些性状变异的主要影响因子;主茎直径、主茎穗长的一般配合力基因型方差占总方差的比重(Vg)和特殊配合力基因型方差占总方差的比重(Vs)相差不大,表明这两个性状的遗传受基因的加性、非加性效应共同作用,其中主茎穗长以加性效应为主,主茎直径以非加性效应为主;株高、单株分蘖数、主茎叶片数、中部叶片长度、中部叶片宽度的Vs远大于Vg,说明这几个性状的遗传受基因的非加性效应影响较大。
遗传力分析得出(表7),7个主要农艺性状的广义遗传力顺序依次为主茎穗长>株高>主茎直径>中部叶片长度>主茎叶片数>单株分蘖数>中部叶片宽度,狭义遗传力顺利依次为主茎穗长>主茎直径>中部叶片长度>单株分蘖数>株高>主茎叶片数>中部叶片宽度。主茎穗长的广义遗传力和狭义遗传力均较大,分别为91.06%和49.61%,说明其受加性基因影响显著;株高、单株分蘖数、主茎叶片数、主茎直径以及叶片中部长度等性状的广义遗传力均在50%以上,而狭义遗传力均小于35%,且差值较大,表明这几个农艺性状非加性遗传的作用比较突出;而中部叶片宽度的广义遗传力和狭义遗传力均最小,分别为40.1%和7.33%,说明其受遗传决定能力较弱,受环境影响较大。
3 讨论与结论
以往高丹草育种研究只侧重农艺性状的选择和产量的提高,常常忽视品质性状的改良;BMR基因型突出特性是显著降低了木质素的含量而增加了饲草消化率,因此,培育BMR基因型高丹草新品种能大幅度改善高丹草的饲用品质。BMR基因型高丹草新品种选育的关键在于选择杂种优势强大的组合,而杂种优势的强弱主要取决于亲本的一般配合力和特殊配合力的效应值。但本研究结果得出,亲本一般配合力高的组合的特殊配合力并不一定高,杂交优势最强的组合其合其双亲不一定都有最好的一般配合力,亲本一般配合力的高低与其组合特殊配合力的高低并没有必然的相关性。在主茎直径性状中,BMR基因型高粱不育系AMP454和BMR基因型苏丹草ZS-102的一般配合力最高,其特殊配合力效应值也最高;但在株高性状中,BMR基因型高粱不育系T98A和BMR基因型苏丹草15C-ZY487的一般配合力最高,其组合的特殊配合力却不是最高;主茎穗长性状中,特殊配合力效应值最高的组合不是AMP454×ZS-102,而是AMP21×15C-ZY487,二者的一般配合力均不是最高;这与段有厚等[18]、高海燕等[19]研究结果相一致。由此表明,杂交种优良亲本的选育在考虑一般配合力的同时,也要兼顾特殊配合力的作用,这是因为一般配合力受基因的加性效应决定的,遗传较稳定,而特殊配合力受基因的非加性效应影响显著,受环境影响较大[20];这就需要在实际育种工作中,通过对杂交组合进行多年多点的小区品比试验表现,以此挖掘特殊配合力较强的杂交组合。
在双亲一般配合力的基础上选配特殊配合力高的组合,已成为杂种优势利用的重要技术原则[21]。本研究通过分析不同BMR基因型高粱不育系的一般配合力效应得出,BMR基因型高粱不育系中,AMP454在主茎叶片数、主茎直径、叶片中部长度、主茎穗长等性状上的一般配合力较高,T98A在株高、单株分蘖数、叶片中部宽度等性状上的一般配合力较高,而AMP19在单株分蘖数、主茎叶片数、主茎穗长上的一般配合力较高,综合分析以AMP454、AMP19、T98A的一般配合力较高,可作为理想的不育系材料。在此基础上,进一步通过分析特殊配合力效应得出,杂交组合AMP19×ZS-102、AMP454×ZS-102在与饲草产量相关的株高、单株分蘖数、主茎叶片数、主茎直径以及中部叶片长度、中部叶片宽度等性状上SCA值表现较高,可为目标组合进行进一步的鉴定、评价提供理论依据。然而,在配合力测定方法上还未见统一的标准,在配合力研究中,因供试材料的亲本数量、农艺性状指标选择不同,得出的结果也不尽一致;本研究为测定19份BMR基因型高粱不育系的配合力,选用了3份自育的不同类型BMR苏丹草种质作为测验种,从7个农艺性状指标上进行了配合力效应分析;在BMR基因型高丹草亲本群体数量确定、农艺指标选择上是否合理,还需进一步研究。同时得出,构建一套科学合理的适宜于配合力效应分析的标准体系,对挖掘配合力强的亲本材料和杂交组合,以及明确农艺性状指标在杂交后代的表现规律是非常有必要的。

表6 各杂交组合主要农艺性状的特殊配合力(SCA)效应值Table 6 Specific combining ability for the major agronomic characters of every hybrid combination
续表6

组合名称 Combination 株高Plantheight单株分蘖数Tillernumber主茎叶片数Leafnumber主茎直径Stemdiameter中部叶片长度Middleleaflength中部叶片宽度Middleleafwidth主茎穗长MainpaniclelengthAMP15×15C-ZY4870.2010.862.131.732.79-6.96-4.96AMP17×15C-ZY4873.28-32.132.1310.38-3.315.19-5.73AMP19×15C-ZY487-9.21-49.32-1.39-4.49-2.39-5.20-7.64AMP21×15C-ZY487-1.7453.851.25-0.322.81-7.8419.84AMP447×15C-ZY487-4.772.26-1.39-1.29-6.01-11.903.61AMP448×15C-ZY4872.712.26-2.2713.54-1.3411.0113.67AMP449×15C-ZY487-0.13-14.93-1.399.656.064.315.53AMP450×15C-ZY487-0.9336.65-3.153.204.9511.019.13AMP451×15C-ZY487-2.26-23.530.374.02-3.23-7.32-6.93AMP452×15C-ZY4871.01-32.13-0.51-7.15-5.584.317.49AMP453×15C-ZY487-8.74-49.32-1.39-5.37-0.501.140.17AMP454×15C-ZY487-0.792.26-7.54-8.53-6.33-0.45-10.10AMP455×15C-ZY487-3.4910.860.372.660.27-6.96-10.81H33A×15C-ZY48712.832.263.89-9.70-3.33-1.68-8.08H41A×15C-ZY48723.1928.052.13-5.135.47-0.803.61H117A×15C-ZY487-1.5528.050.37-7.251.997.48-1.41H129A×15C-ZY4879.5236.654.763.931.516.0710.01

表7 57个杂交组合亲本主要农艺性状的群体遗传参数估计Table 7 Heredity parameter value of the major traits for the 57 hybrid combinations
遗传力表明某一性状受到遗传控制的程度,广义遗传力是指遗传方差占表现型方差的百分数,反映遗传性状选择效果优劣;狭义遗传力指群体内加性遗传方差占表型方差的比率, 能更准确地反映选择效果优劣;而遗传方差中只有加性方差能够稳定遗传[18]。本研究得出,7个主要农艺性状的广义遗传力和狭义遗传力排序不完全一致,表明各性状的基因型是有差异的。主茎穗长的广义遗传力和狭义遗传力均较大,说明由亲本直接传递给杂种的能力较强,可在早代进行选择;株高、单株分蘖数、主茎叶片数、主茎直径以及叶片中部长度等性状的狭义遗传力(h2N)均小于35%,而广义遗传力(h2B)均在50%以上,表明显性效应和上位性效应较大;在早代选择可能会影响选择效果,应结合其所配组合特殊配合力的方差表现在高代(4~5代)进行选择,这样显性上位效应干扰大大降低,所选材料的性状才会相对稳定[21];而中部叶片宽度的广义遗传力和狭义遗传力均小于50%,说明其受土壤水肥条件及环境影响较大。然而,本研究结果与吕鑫等[14]在主茎直径、株高、单株分蘖数等性状,与尹学伟等[15]在主茎穗长上的遗传有一定出入,可能与本研究所选择的亲本均是BMR基因型材料有关。关于BMR基因型材料的配合力效应还需进一步研究。
本研究中的高粱不育系和苏丹草的BMR基因型种类有BMR-6、BMR-12两种类型上,在配合力效应分析上只分析了不同BMR基因型高粱不育系与不同BMR基因型苏丹草的配合力效应,目的是测定所提供的BMR基因型高粱不育系的一般配合力,并没有考虑所组配的杂交种是否也具有褐色中脉这一特征。而田间观察表明,只有当两个亲本材料杂交时,同为BMR-6或同为BMR-12时,才表现出褐色中脉特征,若两个亲本一为BMR-6型,一为BMR-12型,尽管双亲均表现褐色中脉特征,但杂交种却不表现这一特性。因此,要选育、创造BMR基因型高丹草新品种,其前提是双亲都必须为同一基因型种类,在此基础上再进一步分析相同BMR基因型高粱不育系与BMR基因型苏丹草的配合力效应,才具有更科学的意义。同时,配合力的遗传为数量性状的遗传,易受环境因素影响,而且本研究只分析了一年的试验数据,事实上,在综合考虑上述因素的基础上,再进行一年的重复试验或许会得到更客观的结果。
参考文献References:
[1] 李源,游永亮,赵海明,刘贵波,武瑞鑫,杨建忠.种植密度对高丹草农艺性状及饲用品质的影响.草业科学,2017,34(8):1686-1693.
Li Y,You Y L,Zhao H M,Liu G B,Wu R X,Yang J Z.Effect of plant density on agronomic traits and forage quality forSorghumbicolor×S.sudanense.Pratacultural Science,2017,34(8):1686-1693.(in Chinese)
[2] 何振富,贺露,贺春贵,王斐,王国栋,刘陇生.不同种植和收割方式下夏播光敏型高丹草的效益分析.草业科学,2017,34(11):2325-2334.
He Z F,He L,He C G,Wang F,Wang G D,Liu L S.Economic benefit analysis of different summer-sowing patterns and cutting frequencies of three photoperiod-sensitive sorghum-sudangrass hybrids.Pratacultural Science,2017,34(11):2325-2334.(in Chinese)
[3] Lu X P,Yun J F,Gao C P,Surya A.Quantitative trait loci analysis of economically important traits inSorghumbicolor×S.sudanensehybrid.Canadian Journal of Plant Science,2011,91(1):81-90.
[4] Li Y,Mao P S,Zhang W X,Wang X G,You Y L,Zhao H M,Zhai L J,Liu G B.Dynamic expression of the nutritive values in forage sorghum populations associated with white,green and brown midrid genotypes.Field Crops Research,2015,184:112-122.
[5] Oliver A L,Grant R J,Pedersen J F,O’Rear J.Comparison of brown midrib-6 and -18 forage sorghum with conventional sorghum and corn silage in diets of lactating dairy cows.Journal of Dairy Science,2004,87(3):637-644.
[6] Porter K S,Axtell J D,Lechtenberg V L,Colenbrander V F.Phenotype,fiber composition,and in vitro dry matter disappearance of chemically induced brown midrib (bmr) mutants of sorghum.Crop Science,1978,18(2):205-208.
[7] Oliver A L,Pedersen J F,Grant R J,Klopfenstein T J.Comparative effects of the sorghum bmr-6 and bmr-12 genes:I. Forage sorghum yield and quality.Crop Science,2005,45(6):2234-2239.
[8] Ledgerwood D N,DePeters E J,Robinson P H,Taylor S J,Heguy J M.Assessment of a brown midrib (BMR) mutant gene on the nutritive value of sudangrass using in vitro and in vivo techniques.Animal Feed Science and Technology,2009,150(3/4):207-222.
[9] Grant R J,Haddad S G,Moore K J,Pedersen J F.Brown midrib sorghum silage for midlactation dairy cows.Journal of Dairy Science,1995,78(9):1970-1980.
[10] 卢庆善,孙毅.杂交高粱遗传改良.北京:中国农业科学技术出版社,2005:145-176.
Lu Q S,Sun Y.Genetic Improvement of Hybrid Sorghum.Beijing:China Agricultural Science and Technology Press,2005:145-176.(in Chinese)
[11] 张晓娟,张一中,周福平,邵强,柳青山.高粱新选不育系主要农艺经济性状的配合力分析.中国农学通报,2012,28(18):71-75.
Zhang X J,Zhang Y Z,Zhou F P,Shao Q,Liu Q S.Analysis on the combining ability of main agronomic and economic traits for new sorghum male sterility lines.Chinese Agricultural Science Bulletin,2012,28(18):71-75.(in Chinese)
[12] 王立新,成慧娟,马尚耀,葛占宇,严福忠,张姼,隋虹杰.粒用高粱新选亲本配合力分析.作物杂志,2014(6):56-60.
Wang L X,Cheng H J,Ma S Y,Ge Z Y,Yan F Z,Zhang S,Sui H J.The grain sorghum breeding of parental combining ability analysis.Crops,2014(6):56-60.(in Chinese)
[13] 李巍,王艳秋,邹剑秋,张飞.甜高粱亲本系主要农艺性状配合力分析.中国农业大学学报,2012,17(6):98-102.
Li W,Wang Y Q,Zou J Q,Zhang F.Analysis of the combining ability for major agronomic traits in the breeding lines of sweet sorghum (Sorghumbicolor).Journal of China Agricultural University,2012,17(6):98-102.(in Chinese)
[14] 吕鑫,平俊爱,张福耀,杜志宏,李慧明,杨婷婷,牛皓,姚琳.新选饲草高粱恢复系农艺性状配合力效应分析.草业科学,2016,33(7):1361-1366.
Lyu X,Ping J A,Zhang F Y,Du Z H,Li H M,Yang T T,Niu H,Yao L.Effect analysis on the combining ability of main agronomic traits for new breeding restorer lines derived from forage sorghum.Pratacultural Science,2016,33(7):1361-1366.(in Chinese)
[15] 尹学伟,王培华,张晓春,李泽碧,张志良,唐生佑,马强,谭平.14个糯高粱亲本主要农艺性状配合力及遗传力分析.西南农业学报,2014,27(4):1363-1368.
Yin X W,Wang P H,Zhang X C,Li Z B,Zhang Z L,Tang S Y,Ma Q,Tan P.Analysis of 14 parents glutinous sorghum’s main agronomic characteristics combining ability and heritability.Southwest China Journal of Agricultural Sciences,2014,27(4):1363-1368.(in Chinese)
[16] 戴茂华,刘丽英,吴振良.转Bt基因抗虫棉品种间杂种优势及配合力研究.华北农学报,2013,28(增刊):74-78.
Dai M H,Liu L Y,Wu Z L.Study on heterosis and combining ability of transgenic Bt cotton.Acta Agriculturae Boreali-Sinica,2013,28(supplement):74-78.(in Chinese)
[17] 刘来福,毛盛贤,黄远樟.作物数量遗传学.北京:中国农业科学出版社,1984.
Liu L F,Mao S X,Huang Y Z.Crops Quantitative Genetic.Beijing:Chinese Agricultural Press,1984.(in Chinese)
[18] 段有厚,卢峰,邹剑秋.早熟矮秆高粱亲本系配合力和遗传力分析.辽宁农业科学,2017(2):9-12.
Duan Y H,Lu F,Zou J Q.Analysis on the combining ability and heritability of early-maturing and dwarf traits for sorghum parental lines.Liaoning Agricultural Sciences,2017(2):9-12.(in Chinese)
[19] 高海燕,程庆军,田承华,高鹏,张俊珍,郭睿,范国华.新选高粱亲本系的配合力及遗传力分析.农学学报,2016,6(5):6-10.
Gao H Y,Cheng Q J,Tian C H,Gao P,Zhang J Z,Guo R,Fan G H.Combining ability and heritability of new sorghum parental lines.Journal of Agriculture,2016,6(5):6-10.(in Chinese)
[20] 苟才明,余世权,黄宁,荣廷昭,张吉海,徐克成,李志龙.17个玉米地方种质选系的配合力分析.华北农学报,2015,30(2):175-182.
Gou C M,Yu S Q,Huang N,Rong T Z,Zhang J H,Xu K C,Li Z L.Analysis on combining ability of 17 inbred lines from maize landrace.Acta Agriculturae Boreali-Sinica,2015,30(2):175-182.(in Chinese)
[21] 李陈建,付彦博,万江春,王玉祥,张博.30份苏丹草种质资源农艺性状的遗传多样性分析.草业科学,2015,32(1):85-93.
Li C J,Fu Y B,Wan J C,Wang Y X,Zhang B.Genetic diversity of agronomic characteristics of 30 Sorghum sudanense germplasm.Pratacultural Science,2015,32(1):85-93.(in Chinese)
猜你喜欢
西北农业学报(2024年7期)2024-07-12 20:26:59
农业科技通讯(2023年1期)2023-02-12 07:09:28
安徽农业科学(2022年19期)2022-10-29 08:55:54
安徽农业大学学报(2022年3期)2022-10-25 12:32:36
养猪(2022年4期)2022-08-17 07:07:02
种子(2021年7期)2021-08-19 01:46:58
猪业科学(2018年5期)2018-07-17 05:56:14
中国马铃薯(2017年1期)2017-03-02 09:15:46
河北农业科学(2016年2期)2016-09-09 01:41:31
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
