
景观格局与植物多样性的关系及其空间尺度效应
——以浑善达克沙地为例
2018-05-16 08:42王庆慧卿凤婷
生态学报 2018年7期
范 敏,彭 羽,王庆慧,米 凯,卿凤婷
中央民族大学,生命与环境科学学院,北京 100081
景观格局是指景观要素在景观空间内的配置和组合形式[1],最近的一些研究表明景观格局对植物群落的配置和植物多样性具有重要的影响[2]。景观格局不仅通过破碎化、栖息地变化、边缘效应等过程直接影响植物多样性[3],还通过影响生态学过程,比如:矿物质和营养的分配、微生物活动等间接影响植物多样性[4],因此景观格局是植物多样性重要的影响因素。景观格局存在着空间尺度效应[5],尺度效应是影响景观格局与植物多样性关系的重要因素[6- 7],对荒漠植被的研究发现,在较小尺度下(<100m),物种多样性对幅度尺度依赖性较强[8];对欧洲南部农牧交错带研究发现,500m尺度下,景观格局是植物α多样性的重要影响因素[9- 10],在研究景观格局与植物多样性关系时不考虑尺度效应,很容易得出错误的结论[11]。
景观格局指数高度浓缩了景观格局特征,是目前景观分析中广泛应用的指数[7]。其量化方式主要有斑块大小、形状、对比度、蔓延度、多样性、破碎度等指数[3]。根据MacArthur等提出的岛屿生物地理学理论,认为区域物种数量随面积的增加呈现幂函数增长,即斑块面积越大,其物种数目越多[12],对城市生态系统研究显示,较大的绿化面积具有较高的维管植物多样性[13];对半干旱草原生态系统研究显示,植物多样性与斑块面积具有显著正相关关系[10],故认为大面积的斑块有助于保护植物多样性。斑块的形状是植物多样性的重要影响因素,一般认为狭窄的或是不规则的斑块有助于提高植物多样性[1-2],但随着研究对象和尺度的不同,结果差异显著。Moser对农业景观的研究发现,植物物种丰富度与景观形状的不规则程度呈现正相关的关系[14];对日本针叶林研究显示,斑块形状与边缘物种多样性呈正相关关系,与内部种具负相关关系[15]。景观多样性对生物多样性的影响已经受到了学者的广泛关注,尤其是在农业生态系统的研究中[1]。虽然目前对景观格局与植物多样性相关性研究较多,但主要集中在森林、草原、农业及城市生态系统中[16],对沙地景观格局与植物多样性相关性研究尚缺乏。基于前人研究结果,本文提出如下科学问题:沙地环境条件下,景观格局对植物α和β多样性指数存在何种影响,这种影响是否具有空间尺度效应?为了回答这些问题,揭示沙地景观格局对植物多样性的影响,于2016年在浑善达克沙地腹地开展了此项研究。
浑善达克沙地是距京津地区最近的沙地,位于北京上风向,是京津地区主要的沙尘源头。沙地植物多样性不仅是沙地生态屏障的重要基础,还是当地居民生存与发展的物质基础,具有重要的保护价值[17]。目前对浑善达克沙地植物多样性影响因素的研究,主要集中在人为干扰和气候因素,景观格局与植物多样性的关系尚不十分清楚。因此,本文通过研究浑善达克沙地景观格局对植物多样性的影响,揭示沙地中景观格局生态影响效应,为沙地生态恢复和生物多样性保护提供科学合理的依据和参考。
1 研究区概况
研究区位于浑善达克沙地腹地内蒙古自治区正蓝旗(41°46′—43°69′N; 114°55′—116°38′E),正蓝旗共包含3个镇、8个苏木和3个国营牧场,2016年草场面积占到了72.3%,植被种类相对丰富。该区属于中温带半干旱大陆性季风气候,主要由固定沙丘、半固定沙丘、流动沙丘和丘间低地、湿地等景观组成。沙地乔木较为单一,主要为榆树,分布面积少而不均匀,多以单株散生或以疏林形式分布,与大量分布的温带草原形成沙地榆树疏林景观。
2 研究方法
2.1 植物群落调查及数据采集
本研究在2016年7月到9月在浑善达克沙地腹地内蒙古正蓝旗进行实地调研,在固定沙丘、半固定沙丘、流动沙丘及低地4种生境代表性区域共选取28个样地,每个样地设置15个1m×1m的草本样方,共计420个样方,进行GPS定位,统计每一样方内草本植物的种类、数量、高度和盖度等数据,计算植物多样性指数。
(1)Shannon-Wiener多样性指数
式中,H为Shannon-Wiener多样性指数,Pi表示第i种个体数占总数的比例,Pi=Ni/N,其中Ni为第i种的个数,N为总个体数。
(2)Simpson优势度指数
式中,D表示Simpson优势度指数,Pi表示第i种个体数占总数的比例。
(3)Pielou均匀度指数
E=H/lnS
式中,H为Shannon-Wiener多样性指数,S表示研究样地中物种总数。
(4)Whittaker指数
βw=S/ma-1
式中,S表示研究样地中物种总数,ma表示为各样方中物种数。
2.2 景观数据采集
研究数据选取2016年7月正蓝旗浑善达克沙地Landsat8遥感影像,数据处理和空间分析采用ERDAS 2013和ARCGIS 10.0软件,根据实地调研情况及研究目的,经过辐射校正、几何精校正,采用面向对象的监督分类法,将研究区景观划分为:低地、固定沙丘、半固定沙丘、流动沙丘、水域及建设用地(道路和居民区)5类景观类型,分类精度均达到85%以上,满足下一步研究条件。分别以样点为中心,建立不同尺度(半径为100、200、300、400、500、600、700、800、900、1000m)缓冲区。运用景观分析软件Patch Analyst计算各个缓冲区内的景观格局指数。本研究选取了景观水平上表征斑块面积、形状、排列方式、邻近度、破碎化及多样性特征的共33个景观格局指数。
2.3 数据处理
在每个尺度,分别将33个景观格局指数与植物物种的α和β多样性指数进行双变量相关性分析,筛选出不同尺度下与植物多样性指数显著相关的景观格局指数。为了探究景观格局与植物多样性关系的空间尺度效应,进一步以景观格局指数为自变量,植物多样性指数为响应变量,利用逐步回归分析方法,将350个野外样方调查数据和不同尺度下的景观格局指数进行回归,筛选出最优回归模型。最后,利用最优回归模型,将70个未参与建模的样地景观格局指数代入进行植物多样性的拟合,并与实际指数进行验证。数据分析在统计分析软件SPSS 23.0和Windows Excel 2010中完成。
3 研究结果
3.1 研究区内植物物种组成分析
根据野外调查数据分析,在不同的景观中,共有植物109种,隶属于菊科(Asteraceae)、藜科(Chenopodiaceae)、禾本科(Gramineae)、十字花科(Cruciferae)、伞形科(Umbelliferae)等23个科,62个属;其中菊科、藜科、禾本科和豆科为优势种,在所有景观中分布较广;但像白婆婆纳、圆枝卷柏在沙地中出现频率较低,由于这些偶见种的出现,提高了沙地植被β多样性。
3.2 景观格局指数与植物多样性指数的关系
相关性分析结果显示(表1),不同尺度下植物物种多样性对景观格局指数响应差异很大。景观格局指数与植物优势度指数和均匀度指数相关性较小,其主要对Shannon多样性指数和β多样性指数影响较大。在任何尺度下,斑块面积和核心面积以及景观形状指数均是植物多样性指数的重要影响因素;边缘密度在大于300m尺度上与植物α多样性和β多样性具显著相关性,斑块数量在500—1000m尺度下对Shannon多样性贡献较大,但NumP始终是β多样性重要的影响因素,且与Whittaker指数的正相关性随着尺度的增大逐渐增大;景观多样性是影响植物物种丰富度的重要因素,景观丰富度指数(PR)在较大尺度下(600—1000m)对植物α多样性作用显著,而SHEI则与物种β多样性显著相关;较大的斑块间距离会降低植物种子传播效率,导致植物多样性降低,故景观中斑块配置方式是影响物种丰富度的重要因子,其主要影响植物H多样性和β多样性,且其影响作用随着尺度的变化出现差异。
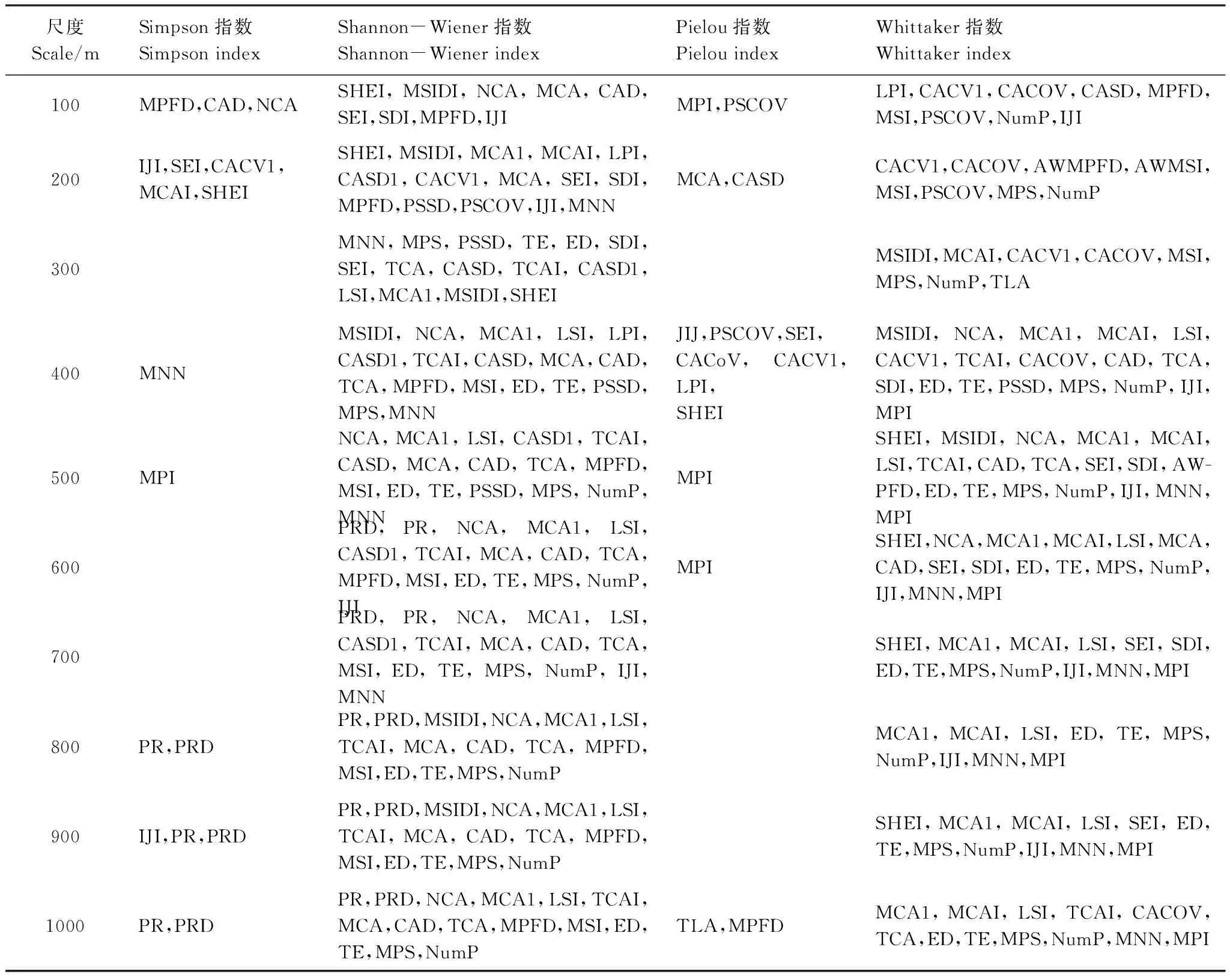
表1 浑善达克沙地不同空间尺度与植物多样性指数显著相关的景观格局指数
MPFD:平均斑块分维数,Mean Patch Fractal Dimension;CAD:核心区密度,Core Area Density;NCA:核心区数量,Number of Core Area;SHEI:香浓均匀度指数,Shannon′s Evenness Index;MSIDI:改良的辛普森多样性指数,Modified Simpson′s Diversity Index;MCA:平均核心区面积,Mean Core Area;MPI:平均接近指数,Mean Proximity Index;PSCoV:斑块面积变异系数,Patch Size Coefficient of Variation;SEI:辛普森均匀度指数,Simpson′s Evenness Index;SDI:香浓多样性指数,Shannon′s Diversity Index;IJI:散布与并列指数,Interspersion Juxtaposition Index;LPI:最大斑块指数,Largest Patch Index;CACV1:斑块核心区面积变异系数,Patch Core Area Coefficient of Variation;CACOV:核心区变异系数,Core Area Coefficient of Variance;CASD:核心区面积标准差,Core Area Standard Deviation;MSI:平均形状指数,Mean Shape Index;MPS:平均斑块大小,Mean Patch Size;NumP:斑块数量Number of Patch;MCA1:斑块平均核心区面积,Mean Core Area per Patch;CASD1:斑块核心区面积标准差,Patch Core Area Standard Deviation;PSSD:斑块面积标准差,Patch Size Standard Deviation;MNN:平均最邻近距离,Mean Nearest Neighbor;AWMPFD:面积加权平均斑块分维数,Area-Weighted Mean Patch Fractal Dimension;AWMSI:面积加权平均形状指数,Area-Weighted Mean Shape Index;TE:总边缘,Total Edge;ED:边缘密度,Edge Density;TCAI:总核心面积指数,Total Core Area Index;LSI:景观形状指数,Landscape Shape Index;TCA:总核心面积,Total Core Area;TLA:景观面积,Total landscape Area;PR:斑块丰富度,Patch Richness;PRD:斑块丰富度密度,Patch Richness Density
3.3 景观格局对植物多样性的逐步回归结果
利用逐步回归的方法进行景观格局指数的筛选并建立最优模型(P<0.01),结果见表2。由回归模型可以看出,景观格局指数对物种多样性的解释量随着尺度的变化而变化。100m尺度下,景观多样性和形状指数对植物多样性影响显著;200m尺度下,MCA对α指数贡献较大,而形状指数对β指数作用显著;当尺度达到400m和500m时,景观面积和形状指数与物种多样性显著相关;随着尺度的增大,景观多样性指数对α和β多样性指数影响作用增大。不同的尺度下决定系数(R2值)具有差异。综合来看,景观格局与Shannon指数和β指数的相关性较大,但存在尺度差异。在100m尺度下,景观格局对Shannon指数的影响(R2=0.138)最大;景观格局在900m尺度下与β指数的影响(R2=0.2)较100m(R2=0.197)、300m(R2=0.118)、400m(R2=0.140)、500m(R2=0.153)、600m(R2=0.107)、700m(R2=0.107)和1000m(R2=0.152)更为密切。
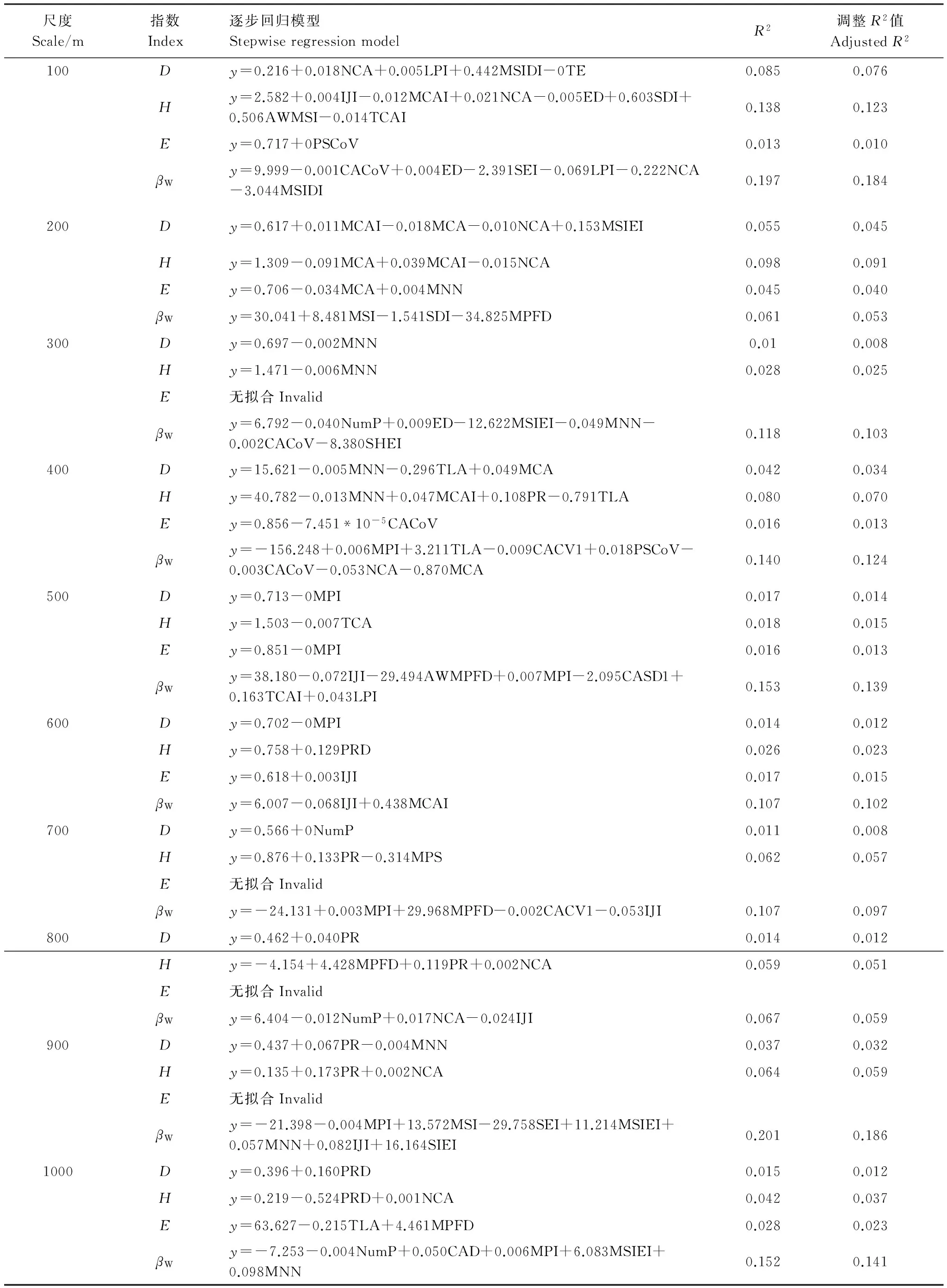
表2 浑善达克沙地不同尺度的景观格局与植物多样性关系最优回归模型
D: Simpson优势度指数, Simpson index;H: Shannon-Wiener多样性指数, Shannon-Wiener index;E: Pielou均匀度指数, Pielou index;βW: Whittaker指数, Whittaker index
3.4 拟合效果检验
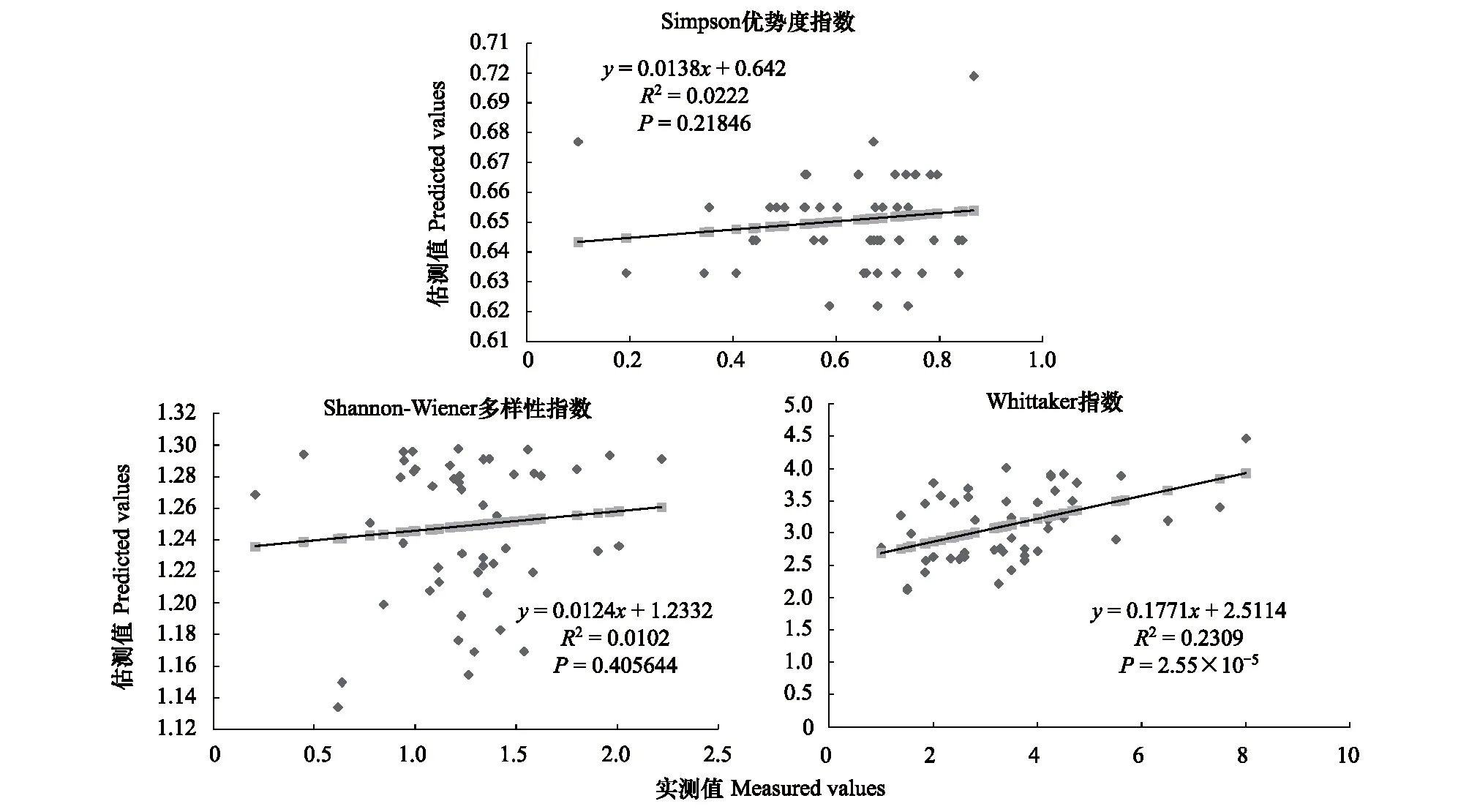
图1 100m尺度下植物多样性实测值与估测值回归模型Fig.1 Regression model of the measured values and predicted values in 100m scale
精度检验结果显示(图1—图10),从100—1000m,拟合效果随着尺度的变化差异显著。均匀度指数在所有尺度下拟合效果均不显著;优势度指数只在500m尺度下拟合果较好;在100、500、600、700、900m尺度下,Shannon多样性指数实测值与回归模型预测值拟合效果显著,β多样性实测值与预测值在100、400、500、800、900、1000m尺度下拟合效果较好。综上所述,在500m尺度下,景观格局对植物多样性影响作用较大,主要景观格局指数是MPI、TCA,运用逐步回归建立的景观模型能够较好的预测植物多样性情况。研究还发现1000m尺度是景观格局对β多样性影响显著的尺度,这可能这预示着需要将研究尺度扩大到1000m以上,才可获得全面准确的沙漠地区植被β多样性信息。
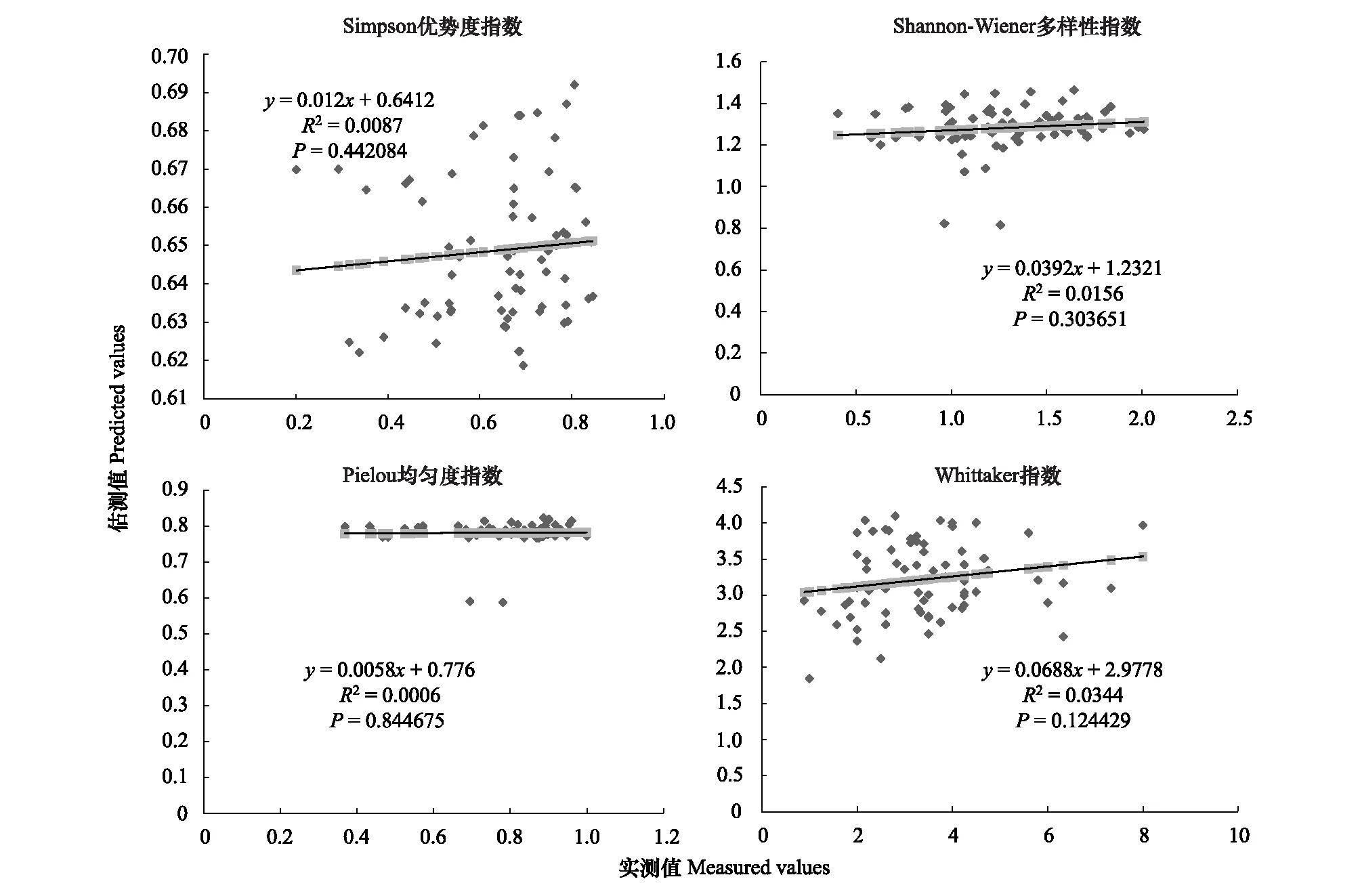
图2 200m尺度下植物多样性实测值与估测值回归模型Fig.2 Regression model of the measured values and predicted values in 200m scale
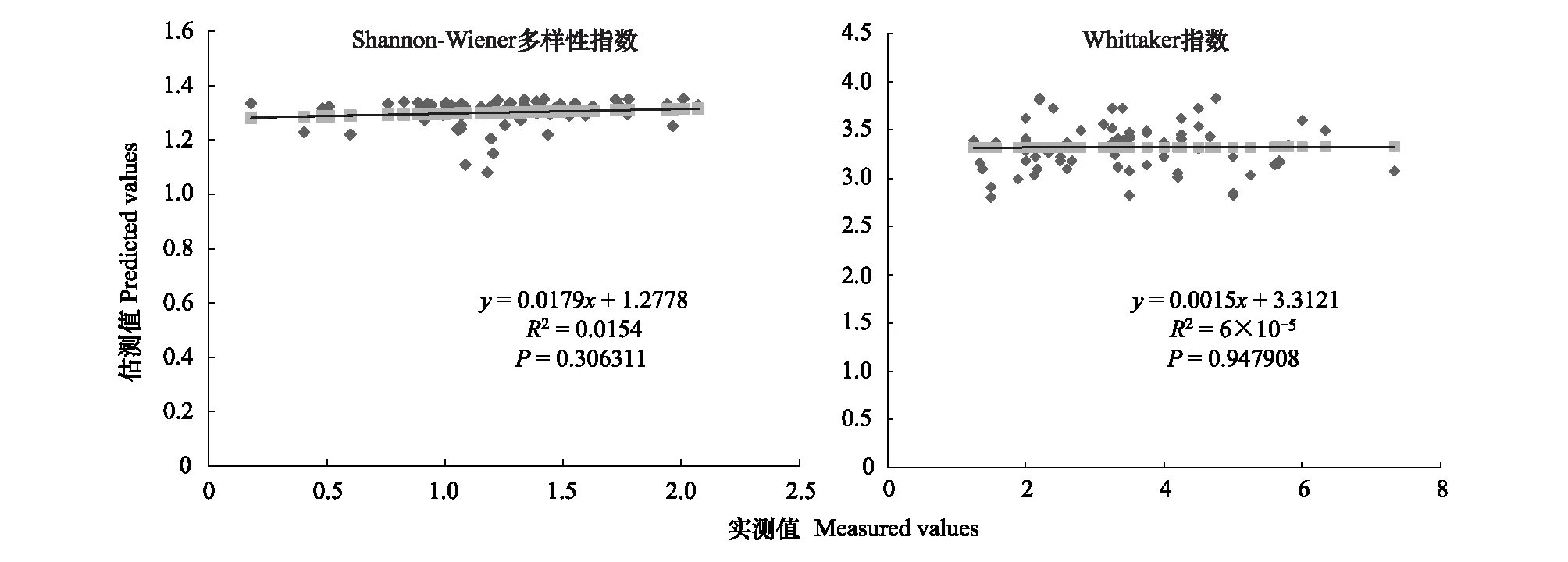
图3 300m尺度下植物多样性实测值与估测值回归模型Fig.3 Regression model of the measured values and predicted values in 300m scale
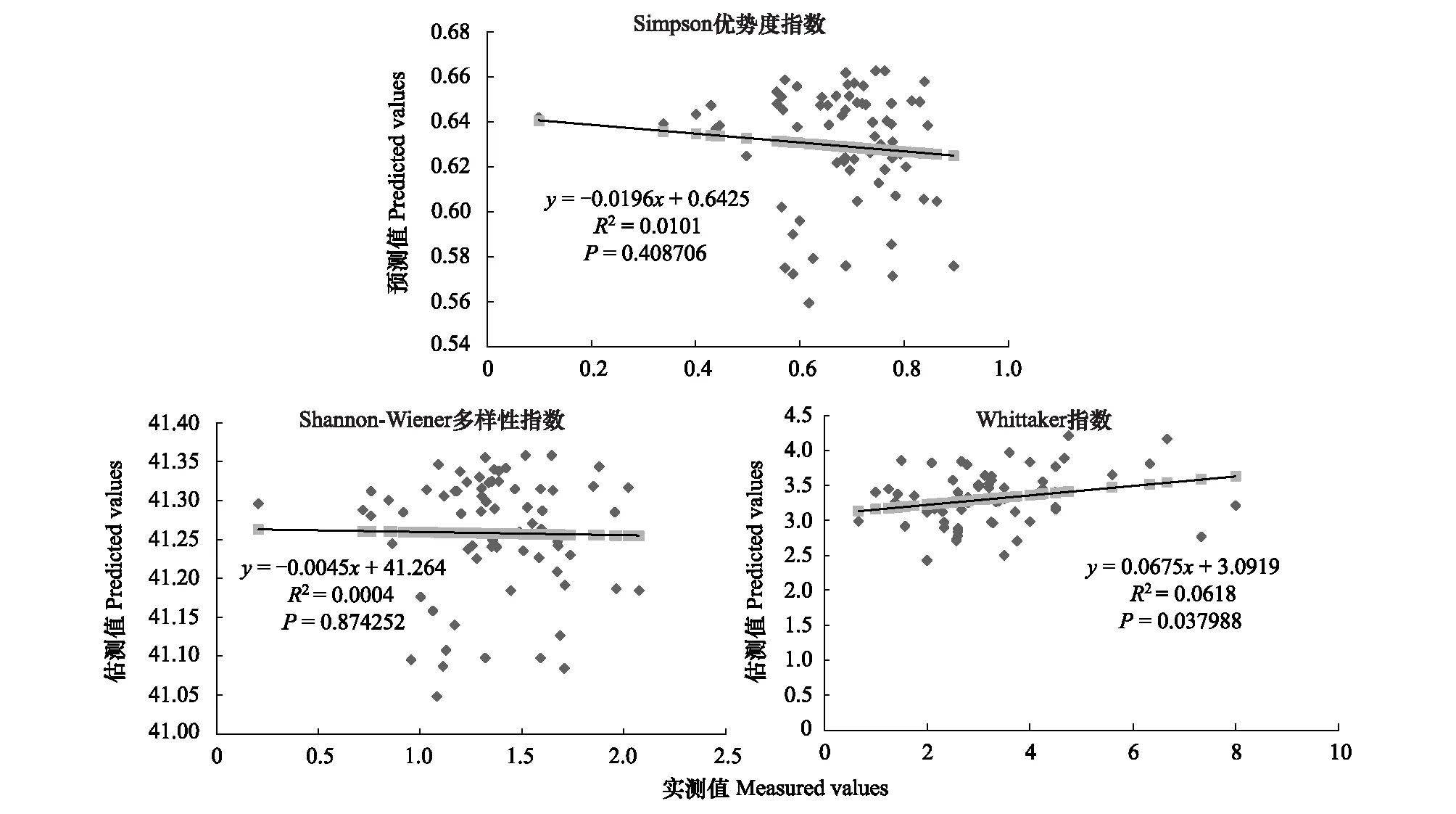
图4 400m尺度下植物多样性实测值与估测值回归模型Fig.4 Regression model of the measured values and predicted values in 400m scale
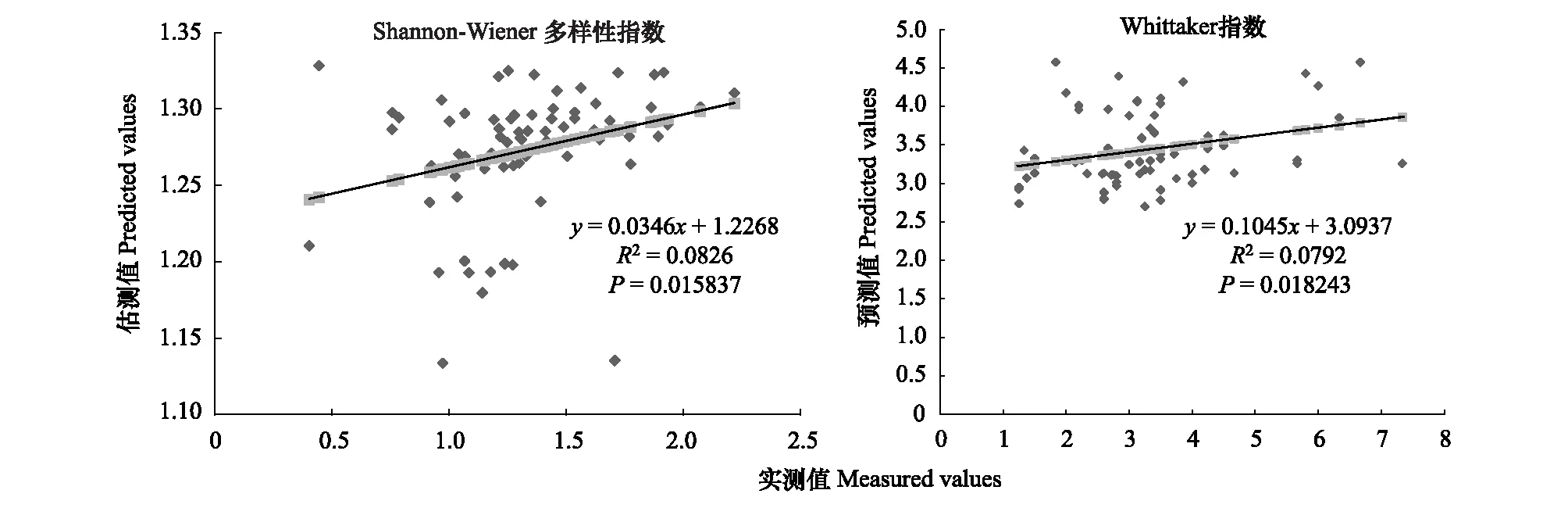
图5 500m尺度下植物多样性实测值与估测值回归模型Fig.5 Regression model of the measured values and predicted values in 500m scale
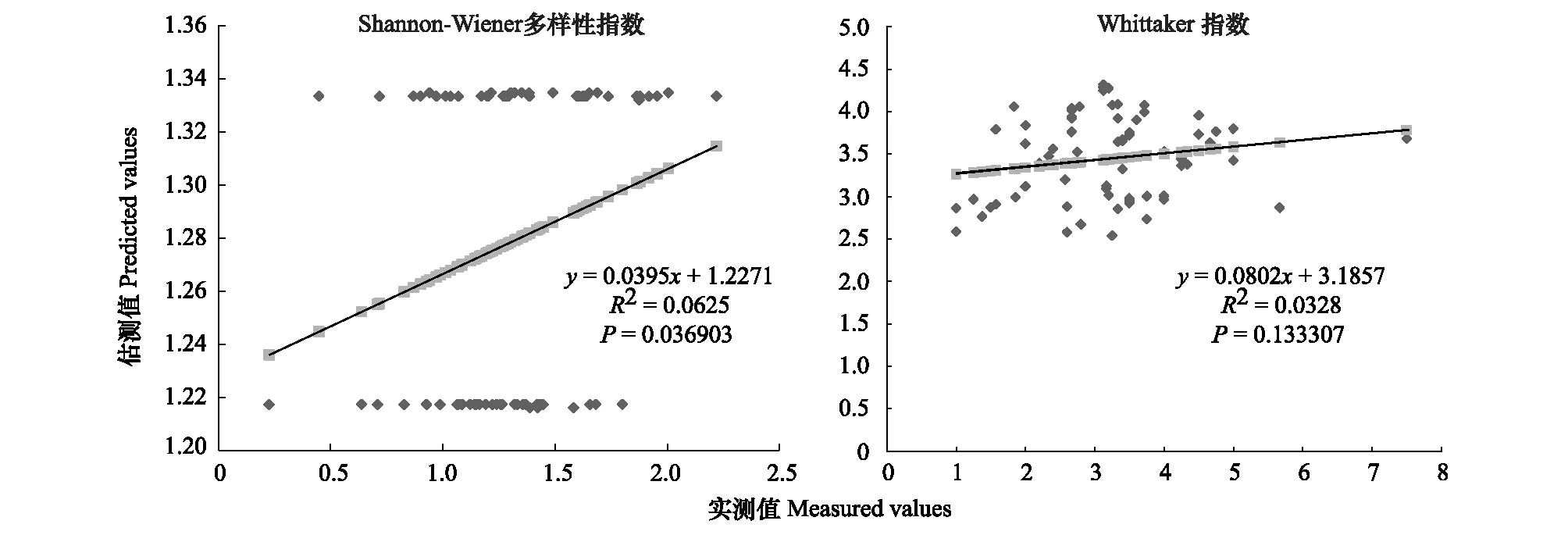
图6 600m尺度下植物多样性实测值与估测值回归模型Fig.6 Regression model of the measured values and predicted values in 600m scale
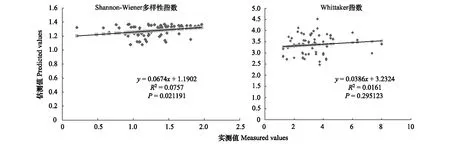
图7 700m尺度下植物多样性实测值与估测值回归模型Fig.7 Regression model of the measured values and predicted values in 700m scale
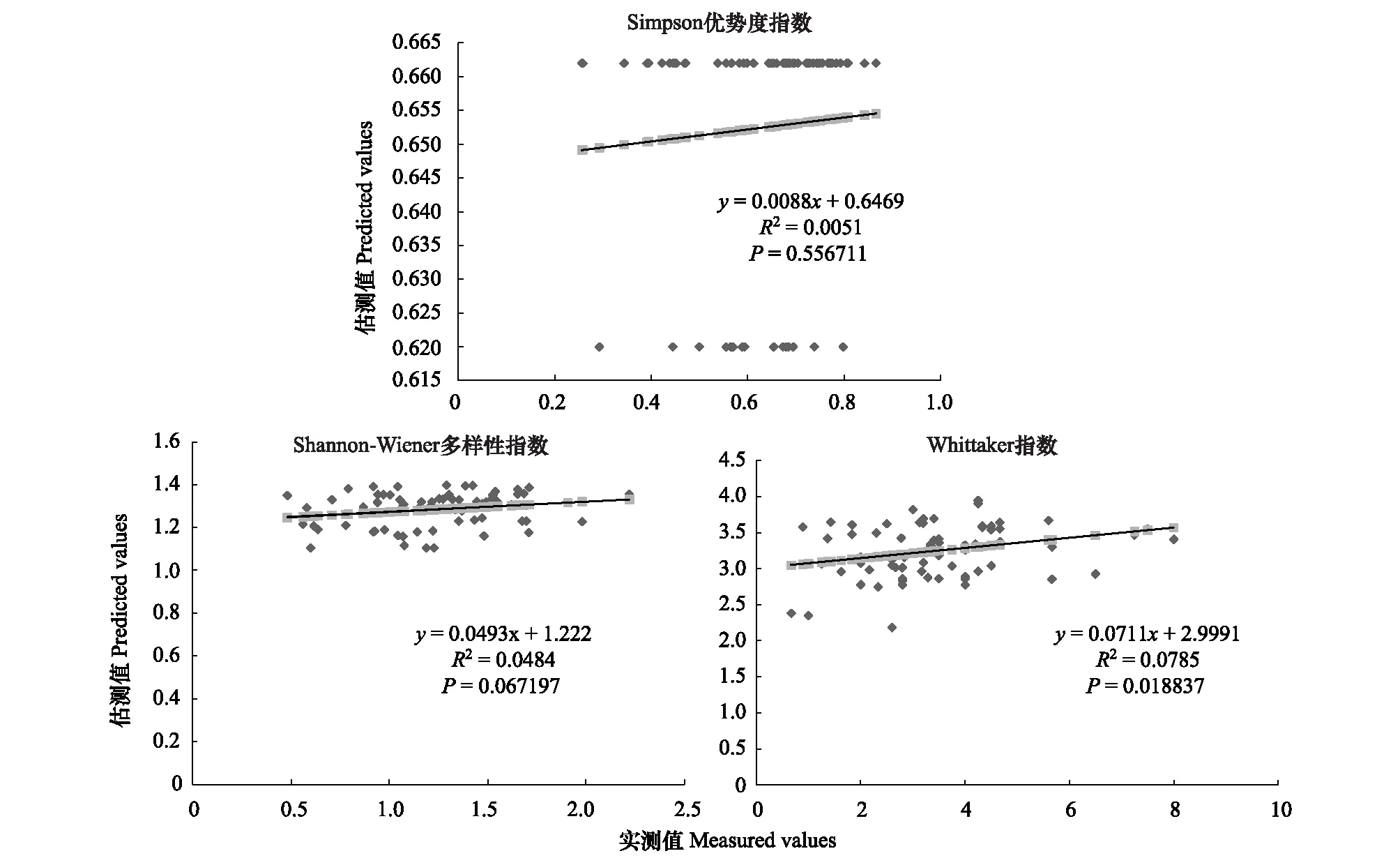
图8 800m尺度下植物多样性实测值与估测值回归模型Fig.8 Regression model of the measured values and predicted values in 800m scale
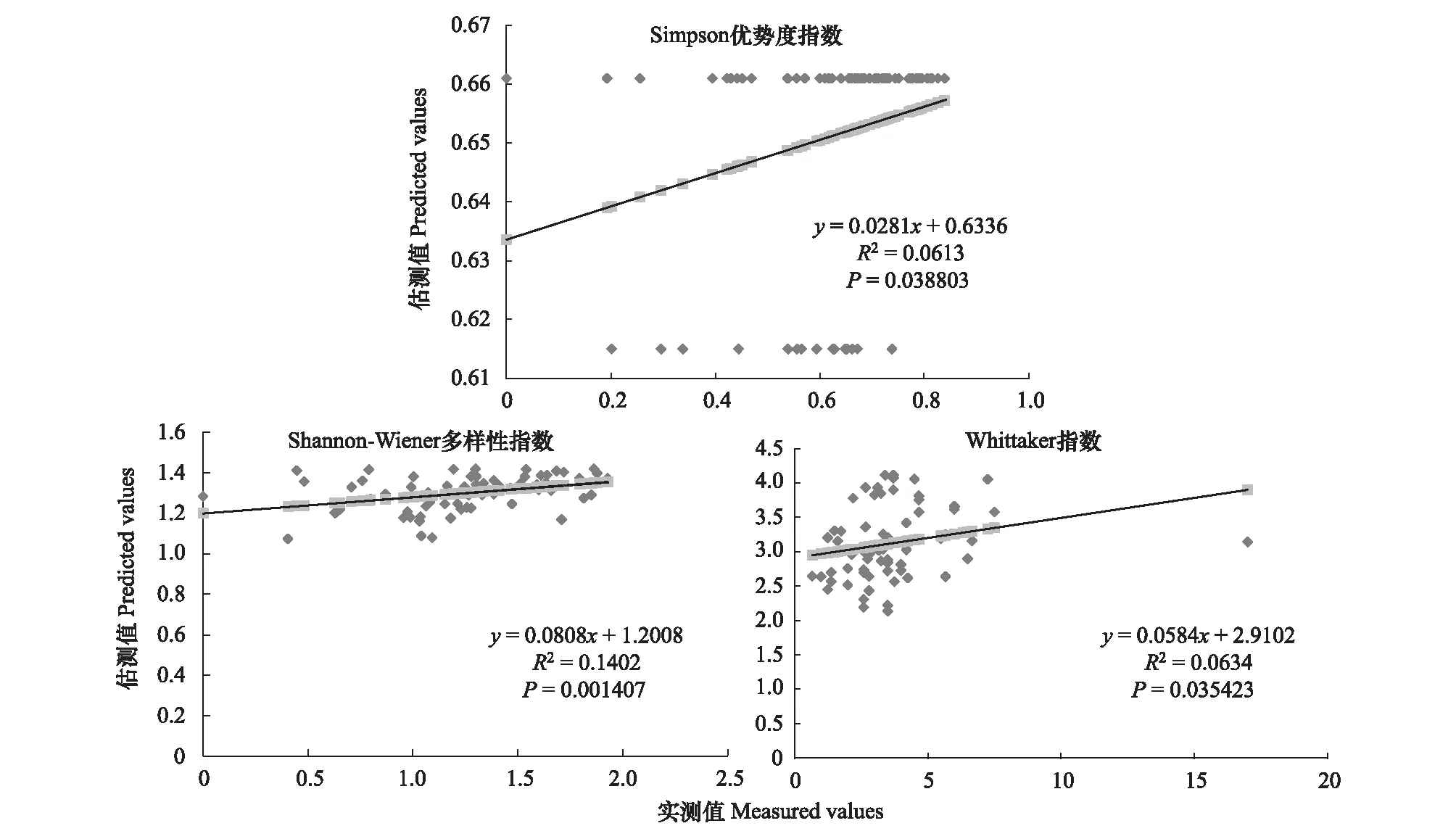
图9 900m尺度下植物多样性实测值与估测值回归模型Fig.9 Regression model of the measured values and predicted values in 900m scale
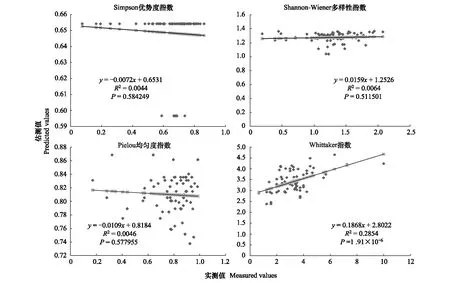
图10 1000m尺度下植物多样性实测值与估测值回归模型Fig.10 Regression model of the measured values and predicted values in 1000m scale
4 讨论
4.1 沙地景观斑块数量和形状影响植物多样性
沙地中不同的景观要素,为植物生存提供了重要的物质基础和环境基础。斑块数量是衡量景观格局的重要指标,已有的研究显示,NumP在小尺度下对植物多样性的影响不显著,随着尺度增加逐渐成为影响植物多样性的主要因子[6,9,18],这与本研究类似。在相关性分析中发现,在大尺度下,斑块数量与Shannon植物多样性关系显著,斑块数量增多导致植物生存面积减小,种群被分割为小种群[19],导致等位基因丢失严重[20],且较小的面积会增加遗传漂变的几率[21],有害基因表达,威胁植物种群的正常生长[22]。但经过逐步回归后,NumP被剔除,这可能是由于景观格局指数之间相关性较强,且其他指数与植物多样性指数更为密切导致的结果。例如斑块面积与数量呈显著相关性。根据生境异质性假说,认为面积大的斑块中具有较多的生境斑块,且容纳的物种数较高[23],故许多尺度下斑块核心面积指数(NCA)与植物多样性指数呈现出显著正相关性。
斑块的形状是植物多样性的重要影响因素,形状复杂的斑块边缘密度较高,边缘生境多样性较高,生物多样性也较高。而且边缘区较多,与外界接触方便,易于进行物质循环和能量流动,边缘效应通过改变生境环境和影响生态过程来影响植物种群的大小和存活率[24]。所以一般认为狭窄的或是不规则的斑块有着较大的边缘长度和边缘密度,景观异质性较高,植物多样性相对较高[2,15]。采用冗余分析法表明,复杂的景观形状(PSSD)的影响效应也是如此[6]。本文采用逐步回归法分析显示,在100、400、800m尺度下,形状指数均与Shannon多样性指数呈现显著的正相关关系,表明复杂的斑块形状有助于提高物种多样性。
4.2 沙地景观多样性及配置方式影响植物多样性
景观多样性(SDI,SEI,PRD)能够提高整个景观异质性,异质性较高的景观有助于提高植物多样性,与大量研究结果一致[6- 10,22- 23]。景观的聚集及离散程度影响植物多样性。Paulo在研究景观格局对生物多样性的影响时发现,景观的紧凑程度对物种和地域之间的关系具有重要的影响[25]。本研究发现,尽管在大尺度下(900、1000m)与植物多样性相关性显著的景观格局指数较多,但经过逐步回归发现,对植物多样性影响作用较大的是景观配置及多样性指数,这可能是由于大尺度下,景观间距离指数生态效应更加显著。β多样性指数与MNN指数呈显著正相关性,异质种群理论认为,适宜的生境总是以离散形式存在,但生境间的距离不能太大,避免基因间交流障碍[20, 24];同时景观连接度对植物种子传粉影响很大,较低的连接度导致传粉率降低[7],形成生殖隔离,同时增加种间近亲交配的概率[26]。直接影响到种子传播效率和植物种子库[17,27]。许多研究表明景观之间的距离越远,生境隔离程度越高,对植物影响越大,尤其是对低传播力的物种影响较大,因为生境间的隔离可能导致种群连通性降低,促进种群灭绝,而聚集的生境会增加生境多样性和生境间的连通性,从而为植物提供更广阔的生存空间和生活资源,有助于增加物种的多样性[7]。故景观斑块间较近的距离有助于提高β多样性。
5 结论
总体来看,本研究验证了沙地疏林草原地区景观格局与植物多样性之间存在相关性,景观格局是物种α多样性和β多样性重要的影响因素,但是具有很强的尺度依赖性。运用逐步回归建立的模型表明,景观格局主要影响Shannon多样性指数和β多样性指数,斑块面积指数在中尺度上(400—500m)对植物多样性贡献较大,在较大尺度上(900—1000m)景观多样性指数和形状指数对植物多样性影响最大;在500m尺度下,各景观格局指数与植物多样性呈现出较密切的关系。未来在不同尺度上管理和应用沙地草地的植物多样性时,可以根据需要的空间尺度,构建有利于植物多样性的优化的景观格局。
参考文献(References):
[1] Walz U. Landscape structure, landscape metrics and biodiversity. Living Reviews, 2011, 5: 3.
[2] Torras O, Gil-Tena A, Saura S. How does forest landscape structure explain tree species richness in a Mediterranean context? Biodiversity and Conservation, 2008, 17(5): 1227- 1240.
[3] López-Martínez J O, Hernández-Stefanoni J L, Dupuy J M, Meave J A. Partitioning the variation of woody plant β-diversity in a landscape of secondary tropical dry forests across spatial scales. Journal of Vegetation Science, 2013, 24(1): 33- 45.
[4] Amici V, Rocchini D, Filibeck G, Bacaro G, Santi E, Geri F, Landi S, Scoppola A, Chiarucci A. Landscape structure effects on forest plant diversity at local scale: exploring the role of spatial extent. Ecological Complexity, 2015, 21: 44- 52.
[5] Bar-Massada B, Wood E M, Pidgeon A M, Radeloff V C. Complex effects of scale on the relationships of landscape pattern versus avian species richness and community structure in a woodland savanna mosaic. Ecography, 2012, 35(5): 393- 411.
[6] Fan M, Wang Q F, Mi K, Peng Y. Scale-dependent effects of landscape pattern on plant diversity in Hunshandak Sandland. Biodiversity and Conservation, 2017, 26(9): 2169- 2185.
[7] Sork V L, Smouse P E. Genetic analysis of landscape connectivity in tree populations. Landscape Ecology, 2006, 21(8): 821- 836.
[8] 何志斌, 赵文智, 常学向, 常学礼. 荒漠植被植物种多样性对空间尺度的依赖. 生态学报, 2004, 24(6): 1146- 1149.
[9] 彭羽, 范敏, 卿凤婷, 薛达元. 景观格局对植物多样性影响研究进展. 生态环境学报, 2016, 25(6): 1061- 1068.
[10] Monteiro A T, Fava F, Gonçalves J, Huete A, Gusmeroli F, Parolo G, Spano D, Bocchi S. Landscape context determinants to plant diversity in the permanent meadows of southern European Alps. Biodiversity and Conservation, 2013, 22(4): 937- 958.
[11] Dan Turtureanu P, Palpurina S, Becker T, Dolnik C, Ruprecht E, Sutcliffe L M E, Szabó A, Dengler J. Scale- and taxon-dependent biodiversity patterns of dry grassland vegetation in Transylvania. Agriculture, Ecosystems & Environment, 2014, 182: 15- 24.
[12] 高俊峰, 马克明, 冯宗炜. 景观组成、结构和梯度格局对植物多样性的影响. 生态学杂志, 2006, 25(9): 1087- 1094.
[13] Matthies S A, Rüter S, Prasse R, Schaarschmidt F. Factors driving the vascular plant species richness in urban green spaces: using a multivariable approach. Landscape and Urban Planning, 2015, 134: 177- 187.
[14] Moser D, Zechmeister H G, Plutzar C, Sauberer N, Wrbka T, Grabherr G. Landscape patch shape complexity as an effective measure for plant species richness in rural landscapes. Landscape Ecology, 2002, 17(7): 657- 669.
[15] Yamaura Y, Kawahara T, Iida S, Ozaki K. Relative importance of the area and shape of patches to the diversity of multiple taxa. Conservation Biology, 2008, 22(6): 1513- 1522.
[16] 米凯. 城市化对植物多样性影响及空间格局优化研究——以北京市顺义区为例[D]. 北京: 中央民族大学, 2015.
[17] 彭羽, 蒋高明, 李永庚, 刘美珍, 牛书丽, 于顺利, 杨道斌. 浑善达克沙地榆树疏林自然保护区核心区设计的初步研究. 植物生态学报, 2005, 29(5): 775- 780.
[18] 覃凤飞, 安树青, 卓元午, 项华均, 郑建伟, 陈兴龙. 景观破碎化对植物种群的影响. 生态学杂志, 2003, 22(3): 43- 48.
[19] Wang Y Y, Zhu Y Q, Wang Y F. Differences in spatial genetic structure and diversity in two mosses with different dispersal strategies in a fragmented landscape. Journal of Bryology, 2012, 34(1): 9- 16.
[20] Cristofani-Yaly M, Novelli V M, Bastianel M, Machado M A. Transferability and level of heterozygosity of microsatellite markers inCitrusspecies. Plant Molecular Biology Reporter, 2011, 29(2): 418- 423.
[21] Finger A, Kettle C J, Kaiser-Bunbury C N, Alentin T, Mougal J, Ghazoul J. Forest fragmentation genetics in a formerly widespread island endemic tree:Vateriopsisseychellarum(Dipterocarpaceae). Molecular Ecology, 2012, 21(10): 2369- 2382.
[22] Hernandez-Stefanoni J L. Relationships between landscape patterns and species richness of trees, shrubs and vines in a tropical forest. Plant Ecology, 2005, 179(1): 53- 65.
[23] 董翠芳, 梁国付, 丁圣彦, 卢训令, 汤茜, 李栋科. 不同干扰背景下景观指数与物种多样性的多尺度效应——以巩义市为例. 生态学报, 2014, 34(12): 3444- 3451.
[24] Moreira R A, Fernandes G W, Collevatti R G. Fragmentation and spatial genetic structure inTabebuiaochracea(Bignoniaceae) a seasonally dry Neotropical tree. Forest Ecology and Management, 2009, 258(12): 2690- 2695.
[25] Campos P R A, Rosas A, de Oliveira V M, Gomes M A F. Effect of landscape structure on species diversity. PLoS One, 2013, 8(6): e66495.
[26] 刘庆福. 景观异质性对多尺度物种多样性的影响——区分环境与人类活动的时间[D]. 呼和浩特: 内蒙古大学, 2016.
[27] Reitalu T, Purschke O, Johansson L J, Hall K, Sykes M T, Prentice H C. Responses of grassland species richness to local and landscape factors depend on spatial scale and habitat specialization. Journal of Vegetation Science, 2012, 23(1): 41- 51.
猜你喜欢
军事文摘(2021年18期)2021-12-02
内蒙古林业(2021年6期)2021-06-26
幼儿智力世界(2021年4期)2021-05-17
内蒙古民族大学学报(社会科学版)(2020年2期)2020-11-06
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
科普童话·百科探秘(2018年7期)2018-09-13
太空探索(2016年5期)2016-07-12
小说月刊(2015年12期)2015-04-23
时代英语·高三(2014年5期)2014-08-26
