
鸭绿江上游江鳕的性状变异和亚种地位
2018-05-15 02:11杨树勋杨雨壮秦大公殷丽洁
生物学通报 2018年7期
杨树勋 杨雨壮 郑 伟 秦大公 殷丽洁
(1 延边州水产站 吉林延吉 133002 2 北京大学生命科学学院 北京 100871)
0 引言
江鳕Lota lota(Linnaeus)发生学上属北极淡水复合体的种类,是淡水鱼类中最典型的冷水性鱼类,酷寒期于冰下行生殖洄游并于水温近0℃(0.1~0.2℃)时产卵。 该鱼是高纬度北方水域珍贵的经济鱼类,尤以其肥大的肝脏和雄鱼精巢的味美而著称。广泛分布于45°N 以北的欧、亚和北美的内陆水域及海湾处,最南可达美国东侧40°N 的俄亥俄河上游。 在我国仅分布于新疆属北极海水系的额尔齐斯河及东北的黑龙江水系,与黑龙江水系有着地理隔离的鸭绿江上游41~42°N 的水域亦有分布,这亦是欧亚大陆江鳕分布区的最南端角[1]。
江鳕属鳕形目(Gadiformes)鳕亚目(Gadoidae)鳕科(Gadidae)江鳕属(Lota),是鳕科鱼类中唯一的淡水种类。依照谢维多维道夫(1948)[2]的分类,分布在欧洲、亚洲广大北方地区水系流域,包括黑海、里海、贝加尔湖、黑龙江、乌苏里江、鸭绿江上游的江鳕是“江鳕指名亚种”Lota lota lota(Linne);分布在东西伯利亚、 阿拉斯加、 加拿大的江鳕是“细尾江鳕亚种”Lota lota leptura(Hubbs et Schrltz);分布在北美洲的江鳕是“北美斑江鳕亚种”Lota lota maculosa(Lesueuy)[3]。
由于黑龙江水系的江鳕恰位于“江鳕指名亚种”和“细尾江鳕亚种”分布的交错地带,而且位置更加向南扩展,其体态构造的性状变异问题,自然地为鱼类学家所注目[4]。 1958年《生物学通报》所刊“在松花湖内采到的江鳕”一文,是我国对江鳕的首篇文字资料报道,施白南和高岫先生,对江鳕的经济价值和生物学特性做了一般性介绍之后,曾提出对松花湖和鸭绿江江鳕的进一步研究有可能发现一个江鳕新亚种[4]。 此后时跨半个世纪未见这方面的报道。直至2002年笔者发表了对与松花湖同属松花江水系而地理位置更靠近鸭绿江上游的牡丹江上游(含镜泊湖)水域江鳕的性状变异方面的研究结果。 文中用了72 尾江鳕标本37 项体态性状的测算统计数据,与属北极海水系的贝加尔湖支流色楞格河所产江鳕的同样性状进行了对比。 差异显著性检验(t 检验)显示,在全部37 项测算性状中,有17 项差异极显著(P<0.001),7 项差异显著(P<0.05);但用差异系数公式计算,则基本均未达到亚种划分的标准。 2 项刚刚达标的性状经作图分析均难予撑立,结论为尽管两地(水域)江鳕体态性状间存在着不同地理种群间显著差异,其亚种地位还是同属江鳕指名亚种Lota lota lota(Linné)[3]。
本文则是对位处欧、 亚大陆江鳕分布区最南端角,又以长白山为分水岭与松花江上游,有着地理隔离的鸭绿江上游水域的江鳕,进行了同样步骤的系列测量和计算对比,借以察看鸭绿江上游江鳕可否划分出新亚种。
1 材料和方法
本文共测量1+~5+龄,体长15.6~48.8 cm 江鳕标本31 尾(♀♀19,♂♂12),均于2010年1月采自42°N 以南云峰水库以北鸭绿江上游河段。 测量和计算的可量比例性状和可数性状共37 项,测量数据经统计处理后,与“江鳕指名亚种”传统分布区贝加尔湖支流色楞格河江鳕的同样性状项目的统计数据(Сорокин B.H.1976)[5],用Mayr 差异系数公式C.D=|M1-M2|/(s1+s2)[6]进行计算,察看其性状差异有否达到C.D≥1.28 的常用亚种划分标准。对已达标的性状项目,又与属黑龙江水系的松花江支流牡丹江上游江鳕同样性状项目间计算的C.D 值进行察看对比,并将3 个水域同样项目分别做统计数据总体柱形图,再具体分析评价后,方得出结论性意见。
2 结果
表1~表3 汇列了鸭绿江上游31 尾江鳕标本体态性状测量计算的统计数值及Сорокин Β.Η.(1976)报道的从贝加尔湖溯入色楞格河群体江鳕的同样项目测量统计数据[5],以及用Mayr 差异系数公式[6]计算的2 个水域江鳕各对应性状的差异程度的量化数值。
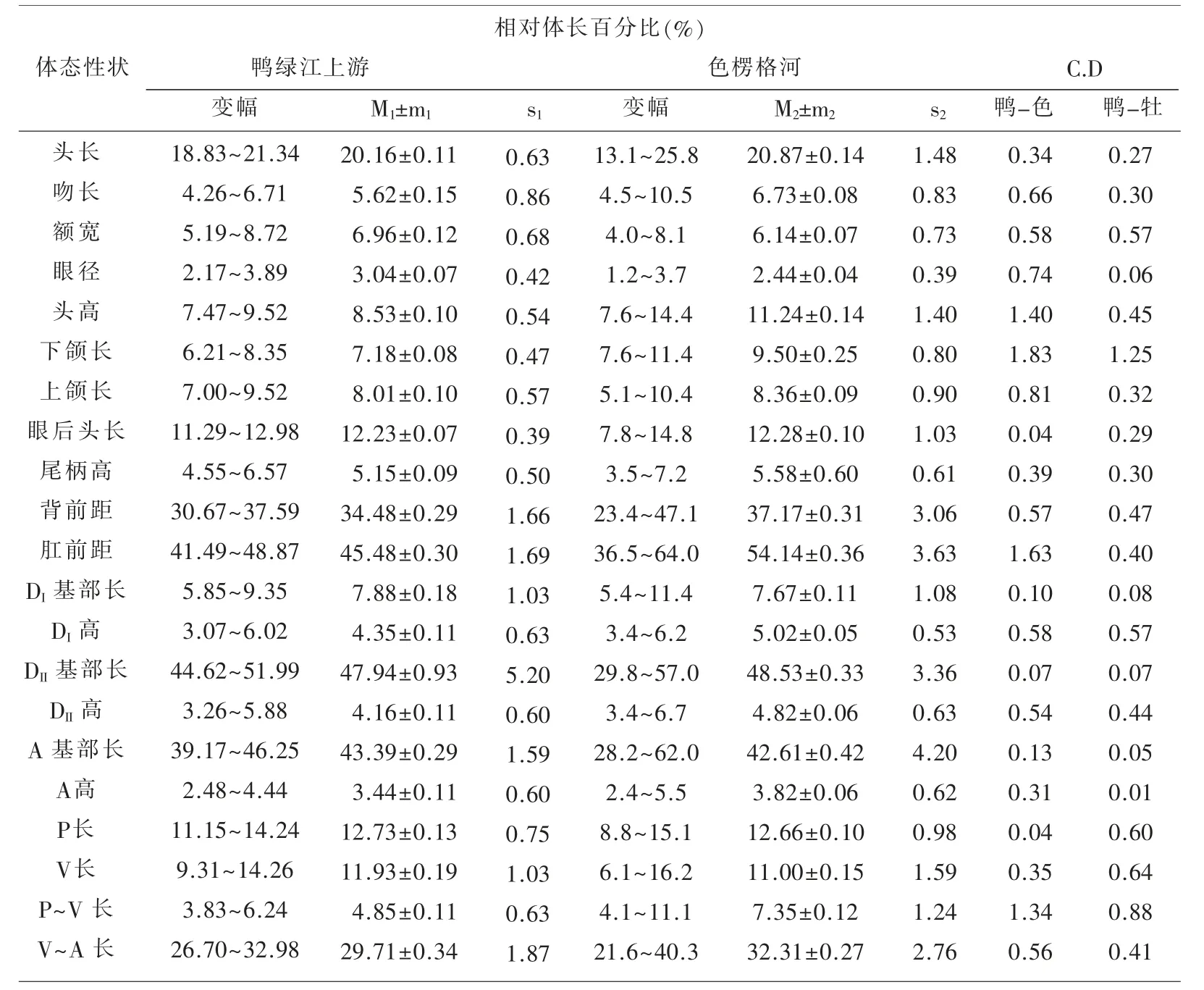
表1 鸭绿江上游与色楞格河江鳕体态性状相对体长百分比的比较
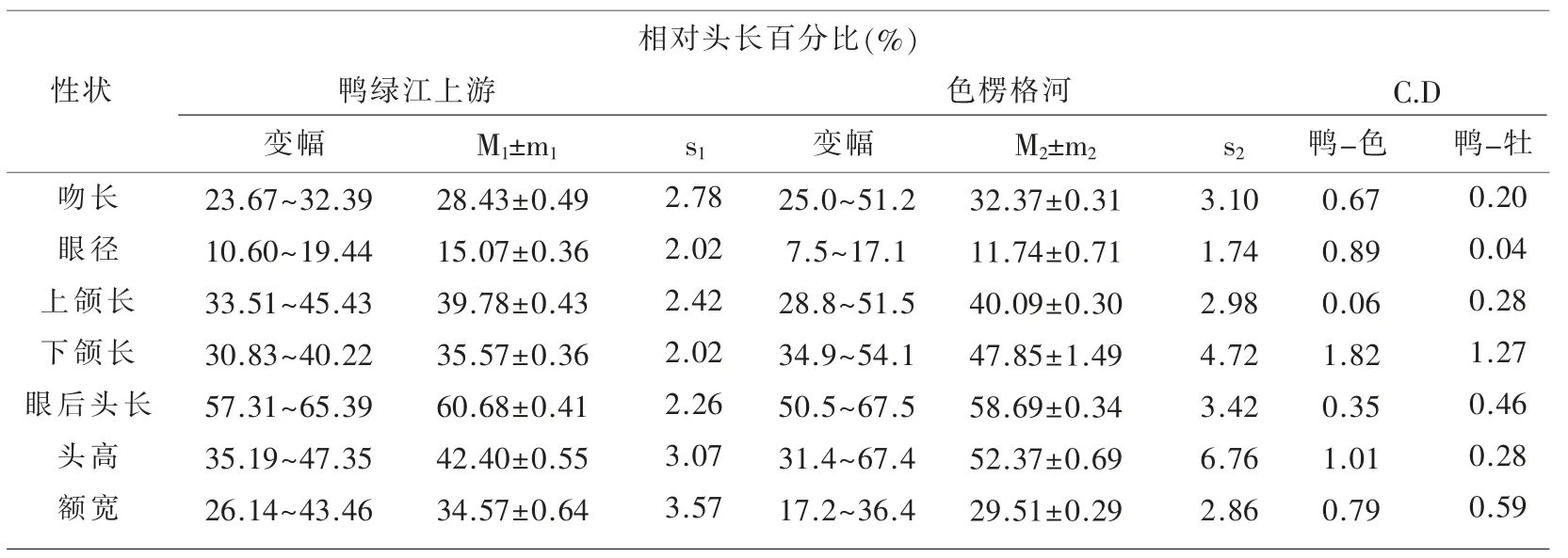
表2 鸭绿江上游与色楞格河江鳕体态性状相对头长百分比的比较
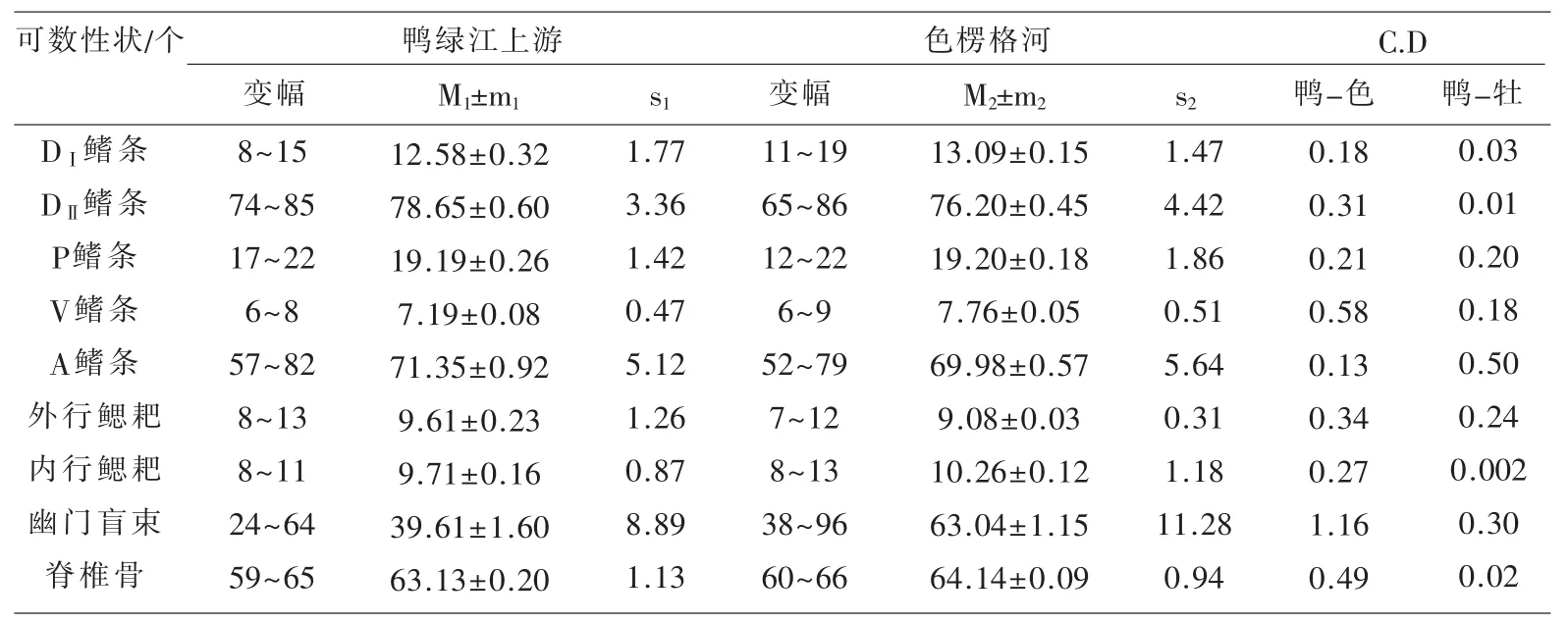
表3 鸭绿江上游与色楞格河江鳕体态可数性状比较
用Mayr C.D≥1.28 亚种划分标准衡量,2 个水域的江鳕间确有5 个性状达标,即头高相对体长的百分比(1.40)、下颌长相对体长的百分比(1.83);肛门前缘至吻端距离相对体长的百分比(1.63)、胸鳍至腹鳍距离相对体长的百分比(1.34);下颌长相对头长的百分比(1.82)。 虽然出现了上述5 个亚种差异达标性状,但不能就此轻易断定出现了新亚种。 Mayr 在提出这一方法时就强调指出:“……方法并没有可能考虑到许多在亚种确认上的生物学和生物地理学因素……还必须借助其他资料予以估量”[6]。作为江鳕指名亚种Lota lota lota(Linné)正宗产地,位处106°E50°N 属北极海水系的贝加尔湖支流色楞格河下游水域,与位处130°E42°N 属太平洋黄海水系的鸭绿江上游水域,其间几乎为整个黑龙江水系的分布区,作为黑龙江最大支流的松花江的发源地(长白山天池),与鸭绿江的发源地(长白山主峰南麓)相距仅20 km,两者属毗邻水域。 整个黑龙江水系包括松花江上游各支流水域均有江鳕分布,所以要确定鸭绿江上游江鳕的亚种地位,必须顾及松花江上游江鳕的性状变异状况和亚种地位。
经用与本文同样的方法,笔者于2002年的研究结果已表明,对可代表松花江上游水系的牡丹江上游(含镜泊湖)江鳕的分类地位,与流入贝加尔湖的色楞格河江鳕同为江鳕指名亚种Lota lota lota(Linné)[3]。所以上述5 个达标性状还必须与牡丹江上游江鳕的同样性状进行对比检验。 经计算两者间差异系数C.D 值分别为:头高/体长(%)0.45;下 颌 长/体 长(%)1.25;肛 前 距/体 长(%)0.40;P ~V 长/体 长(%)0.88;下 颌 长/头 长(%)1.27。5 个性状均未达到和超过C.D≥1.28 的亚种区别标准。
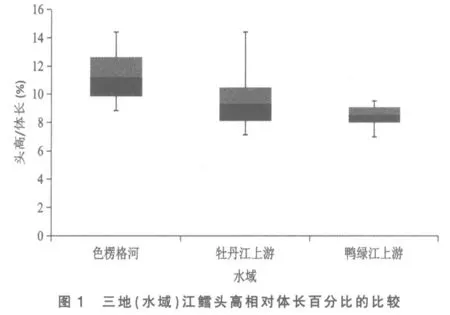
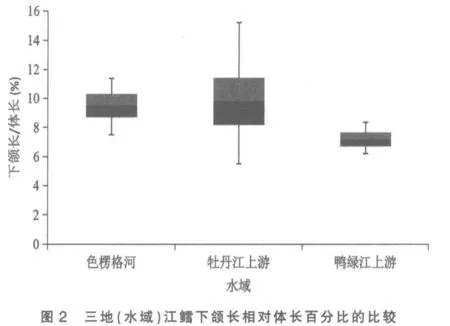
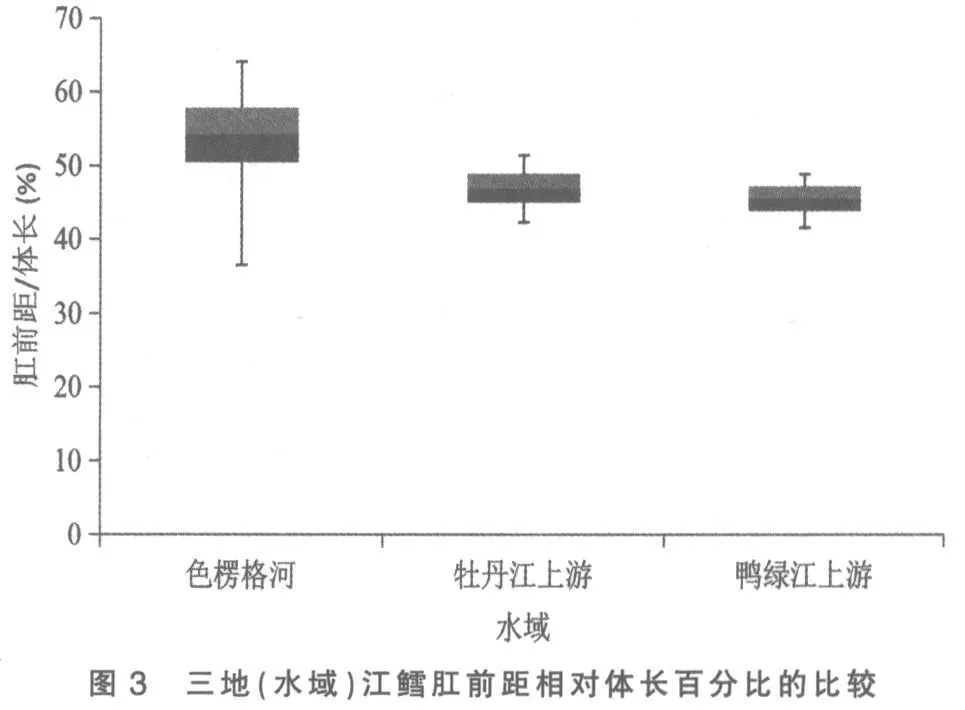
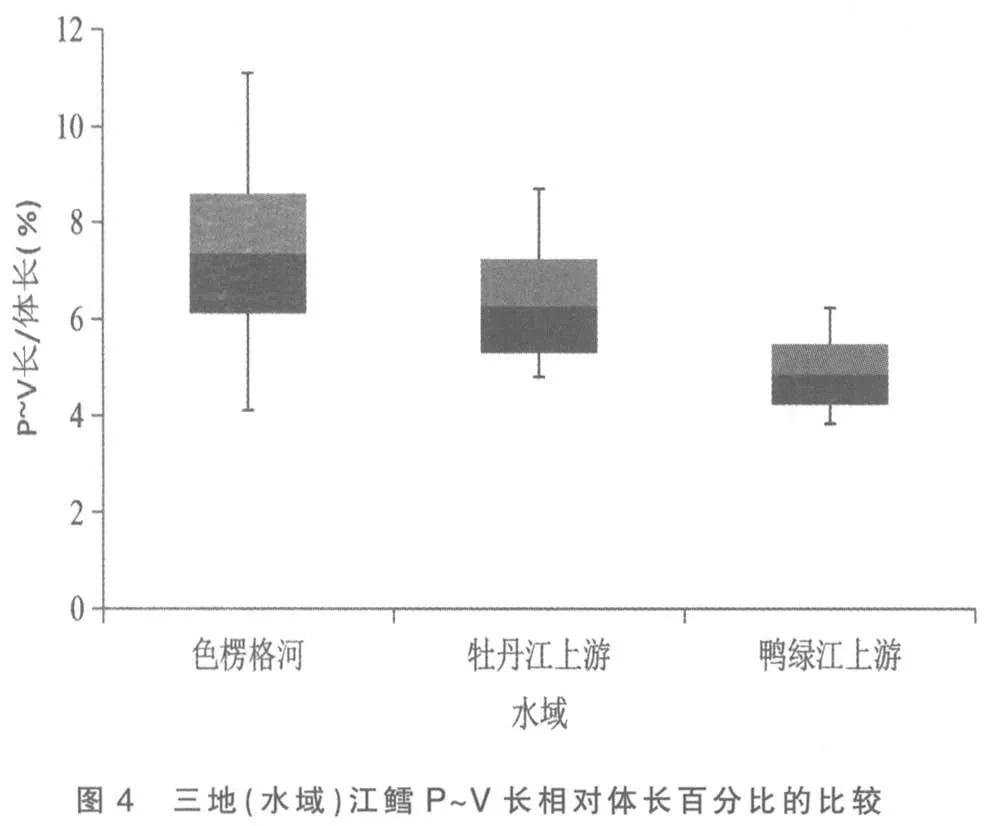
将色楞格河、 牡丹江上游和鸭绿江上游三地(水域)江鳕的这5 个性状分别做其统计数据总体范围及标准差对比柱形图,以更清晰地揭示和显现其性状变异各类情况:图3 为三地(水域)江鳕肛门至吻端距离与体长百分比的比较,可见鸭绿江上游江鳕所测样本变量的中值(平均数)与色楞格河样本中值相距较远,但却与牡丹江上游江鳕相近,依图计算鸭绿江与色楞格河样本间3 倍标准差亦不重叠(此亦Mayr 亚种差异的标志)[6],但与牡丹江上游样本间2 倍标准差即重叠,显然不存在亚种差异; 另外变幅较小的鸭绿江上游与牡丹江上游江鳕样本变量总体均被变幅较大的色楞格河江鳕样本变量总体所涵盖,所以即使C.D 值达标,亦不能立新亚种。从图1、图4 看,三地(水域)江鳕的这2 个性状样本变量的中值及2 倍标准差均呈梯度分布,尽管鸭绿江上游与色楞格河样本间C.D 值达标,但依图计算,3 倍标准差彼此间均会重叠;没达到不予重叠的亚种差异。
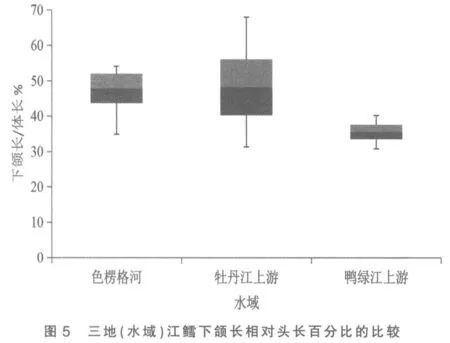
图2 与图5 分别是3 处江鳕下颌长与体长、下颌长与头长百分比的总体范围图。 这2 个表示下颌长度的性状是鸭绿江上游与色楞格河江鳕差异系数达标(C.D≥1.28)的5 个性状中,C.D 值最高的性状(1.82,1.83),与牡丹江上游江鳕也近及达标(1.25,1.27)。2 个性状在图上的显示是:牡丹江上游与色楞格河样本变量的中值非常接近,两者2 倍标准差柱形高度重叠,表明两者同一亚种无疑。鸭绿江上游样本变量的中值,则与上述两地样本中值拉开了较大距离,2 倍标准差柱形亦不重叠,做为亚种差异特征的3 倍标准差,与色楞格河样本确亦不重叠,但与牡丹江上游样本却明显重叠(依图观测),表明没有达到亚种间的差异。
综上所述,属太平洋黄海水系的鸭绿江上游江鳕与属北极海水系的色楞格河江鳕间体态性状变异的差异系数确有5 项达到亚种差异标准,但与发源地相毗邻的松花江支流牡丹江上游江鳕间这5 个性状均没达到亚种分化C.D≥1.28 的差异指标。 既然属黑龙江水系的牡丹江上游江鳕与色楞格河江鳕同为一个亚种[3],显然鸭绿江上游江鳕亦应同属这一亚种,即江鳕指名亚种Lota lota lota(Linné)。 三地(水域)江鳕体态性状间的差异,显然是不同地理种群间的差异。 做为鸭绿江上游江鳕种群体态性状变异的突出特点是下颌较长,差异系数表明其下颌长度明显超过牡丹江上游江鳕,更超过色楞格河江鳕,可称之为鸭绿江长下颌种群。
猜你喜欢
中国造纸(2022年8期)2022-11-24
热带亚热带植物学报(2022年4期)2022-08-18
牡丹江医学院学报(2022年6期)2022-02-14
昆虫学报(2021年11期)2021-12-08
鸭绿江(2021年31期)2021-11-18
——和田盘羊
野生动物学报(2021年4期)2021-11-14
鸭绿江(2021年17期)2021-10-13
儿童时代·快乐苗苗(2021年6期)2021-07-01
活力(2019年17期)2019-11-26
食品与发酵工业(2019年18期)2019-10-11
