
影响越冬期意大利蜜蜂健康的病毒种类研究
2018-05-11 08:18:27陈功文温政胜秦瑶许瑛瑛胡福良郑火青
浙江大学学报(农业与生命科学版) 2018年2期
陈功文,温政胜,秦瑶,许瑛瑛,胡福良,郑火青
(浙江大学动物科学学院,杭州310058)
人类所需食物的三分之一依赖于昆虫授粉[1],每年仅通过昆虫授粉就可使美国的种植业增值超过150亿美元[2]。以蜜蜂为主的授粉昆虫数量不足不仅会影响农作物授粉,对粮食安全产生威胁,还会对植物多样性及生态平衡带来不利影响[3]。西方蜜蜂(Apis mellifera)是世界养蜂业使用的主要蜂种,近年来全球范围的蜂群损失现象严重。由于在越冬期间蜂群面临更大的环境压力,而且蜂群内个体停止更新换代,因此越冬期是蜂群损失的高发期。据报道,西方蜜蜂蜂群越冬期损失率在美国、欧洲和非洲分别达到22.5%~35.8%[4]、7%~30%[5]和 29.6%[6]。虽然 LIU 等[7]对2010—2013年我国12个主要西方蜜蜂饲养省份蜂群越冬损失情况的调查中,我国西方蜜蜂蜂群越冬期间平均损失率仅为8.9%,但近年来在江浙地区出现的大面积越冬蜂死亡现象引起了业界的普遍担忧[8-9]。
病原菌、寄生虫、饲养条件和环境条件是影响蜜蜂健康的主要因素。近年来,蜜蜂病毒流行率很高,大量的流行病学研究都表明病毒感染在越冬蜂群损失中扮演着重要角色[10-12]。目前,已在蜂群中发现24种病毒[13-14],其中最为常见的7种病毒包括:蜜蜂急性麻痹病毒(Acute bee paralysis virus,ABPV)、黑蜂王台病毒(Black queen cell virus,BQCV)、慢性蜜蜂麻痹病毒(Chronic bee paralysis virus,CBPV)、残翅病毒(Deformed wing virus,DWV)、以色列蜜蜂麻痹病毒(Israeli acute paralysis virus,IAPV)、克什米尔病毒(Kashmir bee virus,KBV)和囊状幼虫病毒(Sacbrood virus,SBV)。ABPV、IAPV和KBV是3种同源性较高的急性病毒,感染蜜蜂后会引起病蜂身体颤抖,逐渐麻痹,导致急性死亡[15]。在美国和德国,至少分别有2项长期的研究有力地证明了IAPV和ABPV与冬季蜂群损失间有紧密联系[10,16]。DWV的毒力相对较低,但由于流行率非常高,且与狄斯瓦螨(Varroa destructor)存在很强的协同作用,被认为是导致全球范围内蜂群损失的主要因子之一[17]。BQCV主要对蜂王的幼虫和蛹产生危害,常以隐性感染的方式长期存在于蜂群中,对蜂群的危害较小[18-19];SBV主要感染东方蜜蜂,对西方蜜蜂危害较低;KBV则尚未在我国蜂群中检出。
本研究通过跟踪浙江省3个蜂场48群意大利蜜蜂(西方蜜蜂的主要亚种,Apismellifera ligustica)在越冬前后蜂群病毒感染情况及蜂群越冬表现,分析越冬期蜜蜂病毒感染变化规律,探析影响越冬期蜜蜂健康的病毒种类,为防控蜜蜂病毒病、减少越冬期蜜蜂死亡提供理论指导。
1 材料与方法
1.1 样品收集
在2016年12月—2017年1月蜂群越冬期间,对浙江省金华市2个蜂场(JH1、JH2)各20个蜂群,以及浙江大学实验蜂场(ZJU)8个蜂群进行越冬前和越冬后取样,并记录所有蜂群越冬期间的健康情况。ZJU的8个蜂群在越冬期间每隔1周连续取样3次(总共取样5次)。每群蜜蜂每次取样超过50只,对所取样本检测 ABPV、BQCV、CBPV、DWV、IAPV和SBV等6种常见蜜蜂病毒的感染情况,并对感染率较高的病毒进行基因组拷贝数定量。
1.2 主要试剂和仪器
RNApure超纯总RNA快速提取试剂盒(北京艾德莱生物科技有限公司)、FSQ-201反转录试剂盒(ToYoBo公司,日本)、聚合酶链式反应(polymerase chain reaction,PCR)试剂盒(GenStar2×Taq PCR StarMix)、QPS-201荧光定量PCR试剂盒(ToYoBo公司,日本)、琼脂糖[生工生物工程(上海)股份有限公司]、DNA标志物(TaKaRa公司,日本)、PCR仪(ABI公司,美国)、NanoDrop 2000分光光度计(赛默飞世尔科技公司)、StepOnePlus荧光定量PCR仪(ABI公司,美国)等。
1.3 RNA提取
将25只蜜蜂放入研钵,加入液氮研磨至粉末,取粉末约170 mg于1.5 mL离心管中,同时加入1 mL RNA裂解液,充分振荡15 s后放置15 min,使核蛋白体完全裂解。参照RNApure超纯总RNA快速提取试剂盒的操作说明提取蜜蜂RNA。用NanoDrop 2000测定所提RNA的浓度。
1.4 反转录反应
取800 ng RNA于200μL离心管中,65℃孵育5 min,置于冰上快速冷却。在上述离心管中加入5×RTMaster Mix 4μL,用无核酸酶水加至反应总体积为20μL。
将上述反应液振荡离心后,按37℃、15 min,50 ℃、5 min,98℃、5 min进行反应,然后置于-20℃备用。
1.5 6种常见蜜蜂病毒定性检测
配置20μL PCR反应体系,其中2×Taq PCR StarMix 10μL,cDNA 2μL,上下游引物各1μL,无核酸酶水6μL。扩增程序为:94℃,2 min;94℃、30 s,55 ℃、30 s,72 ℃、30 s,35个循环;72 ℃,5 min。PCR产物在2%琼脂糖凝胶电泳(110 V,30 min)中检测。引物序列见表1。

表1 定性检测6种常见蜜蜂病毒的引物序列Table1 Sequences of primersused for qualitative detection of six common bee viruses
1.6 病毒绝对定量
通过普通PCR确定感染率最高的3种病毒分别为DWV、BQCV和IAPV。由于BQCV主要对蜂王蛹产生危害,对工蜂和蜂群危害较小[20],因此,本试验不对其进行拷贝数分析。试验制备DWV和IAPV的PCR产物标准质粒溶液,用NanoDrop 2000测得其质量浓度分别为95.2和249.8 ng/μL。拷贝数的计算公式为:拷贝数/μL-1=6.02×1023×[质粒质量浓度/(ng/μL)×10-9]/(DNA长度×660)。其中,DNA长度为所使用质粒的碱基数和质粒所连接片段的碱基数之和。计算得知,二者拷贝数分别为3.06×1010和7.94×1010μL-1。
以质粒标准品为模板,按10倍梯度依次稀释6个不同的浓度,并以不同浓度的质粒溶液为模板进行荧光定量PCR。反应体系(10μL)为:模板1μL,上游和下游引物共1μL,Mix酶5μL,纯水3μL。荧光定量PCR扩增程序为:95 ℃,1 min;95 ℃、15 s,60 ℃、1 min,95 ℃、15 s,共40个循环。溶解曲线分析温度程序为:95 ℃,15 s;60 ℃,1 min;95 ℃,15 s。其中60℃至95℃反应总时间为20 min。DWV定量PCR引物序列为:上游引物5'-CGTGGTGTAG TAAGCGTCGT-3';下 游 引 物 5'-TCATCCGTAG AAAGCCGAGT-3'。IAPV定量PCR引物序列为:上游引物 5'-TCGCTGAAGGCATGTATTTC-3';下游引物5'-ATTACCACTGCTCCGACACA-3'。
对病毒质粒进行梯度荧光定量PCR试验后,根据不同浓度对应的CT值建立2种病毒的标准曲线:yDWV=-3.321 3x+40.914,R2=0.999 7;yIAPV=-3.264 6x+38.918,R2=0.995 5(y代表CT值,x代表病毒基因组拷贝数的常用对数,病毒基因组拷贝数为10x)。
2 结果与分析
2.1 蜂群越冬表现
越冬期间JH1和JH2蜂场均未发现蜂群死亡,所有蜂群越冬情况良好。而在ZJU蜂场的8个蜂群中,有1个蜂群在越冬期间死亡,另有4个蜂群在越冬后(1月初)濒临死亡,箱内只剩少许工蜂。
2.2 蜂群病毒感染情况
由图1可知,3个蜂场48群蜜蜂均未感染ABPV、CBPV和SBV,而BQCV、DWV和IAPV感染率较高。JH1蜂场蜂群越冬前和越冬后的病毒感染率均依次为BQCV=DWV>IAPV,其中,BQCV和DWV感染率在越冬前达到95%,越冬后达到100%。JH2蜂场蜂群越冬前仅感染BQCV和DWV,感染率分别为100%和40%;越冬后有2群(10%)出现IAPV感染。ZJU蜂场蜂群越冬前病毒感染率依次为DWV=IAPV>BQCV,其中DWV和IAPV感染率均达到100%;越冬后病毒感染率依次为IAPV>DWV>BQCV。
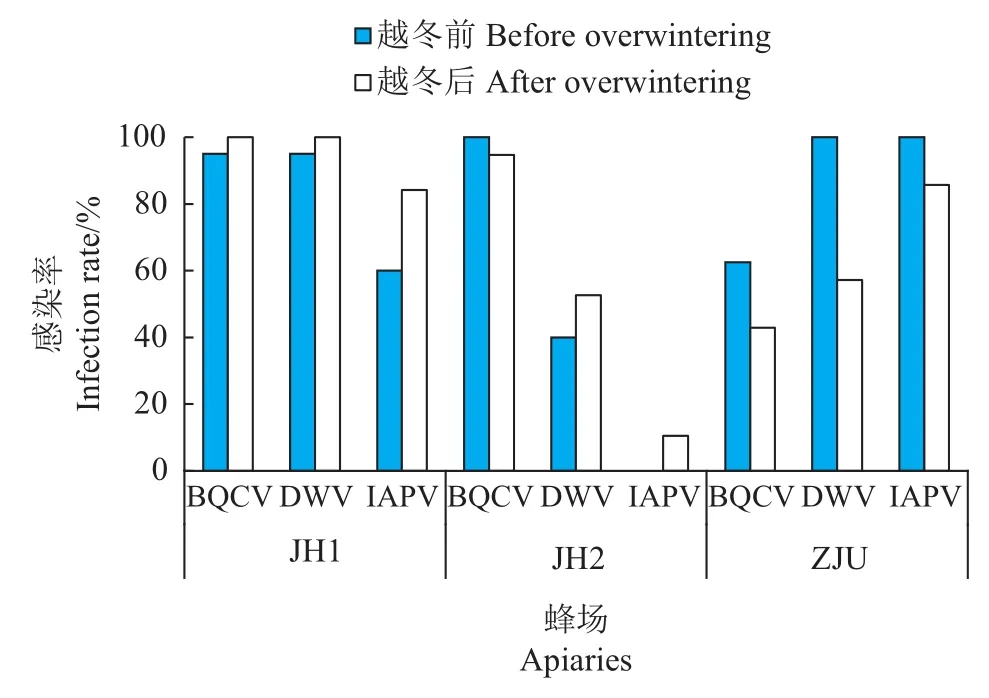
图1 JH1、JH2和ZJU蜂场蜂群越冬前和越冬后病毒感染率Fig.1 Infection ratesof virusesin honey beecoloniesin apiaries JH1,JH2 and ZJU before and after overwintering
2.3 蜂群越冬前和越冬后DWV和IAPV病毒量比较
从图2A中可以看出,JH1蜂场蜂群的DWV和IAPV的病毒基因组拷贝数都相对较高,越冬前DWV基因组拷贝数达到1×107左右,IAPV基因组拷贝数达到1×106左右,蜂群越冬后DWV和IAPV基因组拷贝数都有显著升高(P<0.05)。从图2B中可以看出,JH2蜂场蜂群的病毒基因组拷贝数在越冬前和越冬后都相对较低,越冬前DWV基因组拷贝数只有1×104左右,IAPV基因组拷贝数只有1×102左右,越冬后DWV基因组拷贝数显著上升(P<0.05),但IAPV基因组拷贝数无显著变化(P<0.05)。从图2C中可以看出,ZJU蜂场蜂群的这2种病毒量也都相对较高,其中DWV基因组拷贝数均值接近1×108,越冬后IAPV基因组拷贝数有上升趋势,但差异不显著(P>0.05),可能是因为样本量偏少。
比较ZJU蜂场越冬后濒临死亡蜂群和正常蜂群的病毒基因组拷贝数,结果(图3A)发现,濒临死亡蜂群的IAPV极显著增高(P<0.01),其基因组拷贝数均值达到了1×109以上。比较越冬过程中或越冬后死亡的5个蜂群和越冬后仍健康存活的3个蜂群在越冬前的病毒滴度,发现前者IAPV基因组拷贝数的平均值更高,但两者差异不显著(图3B)。
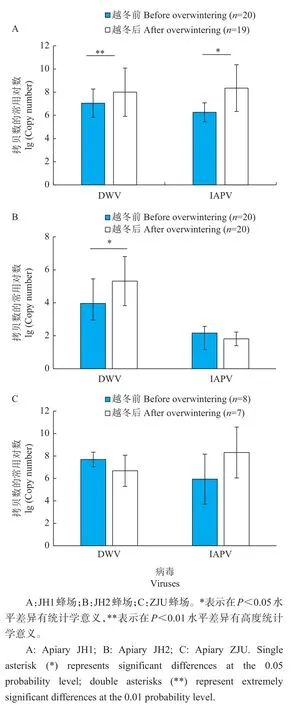
图2 3个蜂场蜂群在越冬前和越冬后DWV和IAPV病毒基因组拷贝数比较Fig.2 Genomecopy numbers of DWV and IAPV in honey bee colonies before and after overwintering in three apiaries
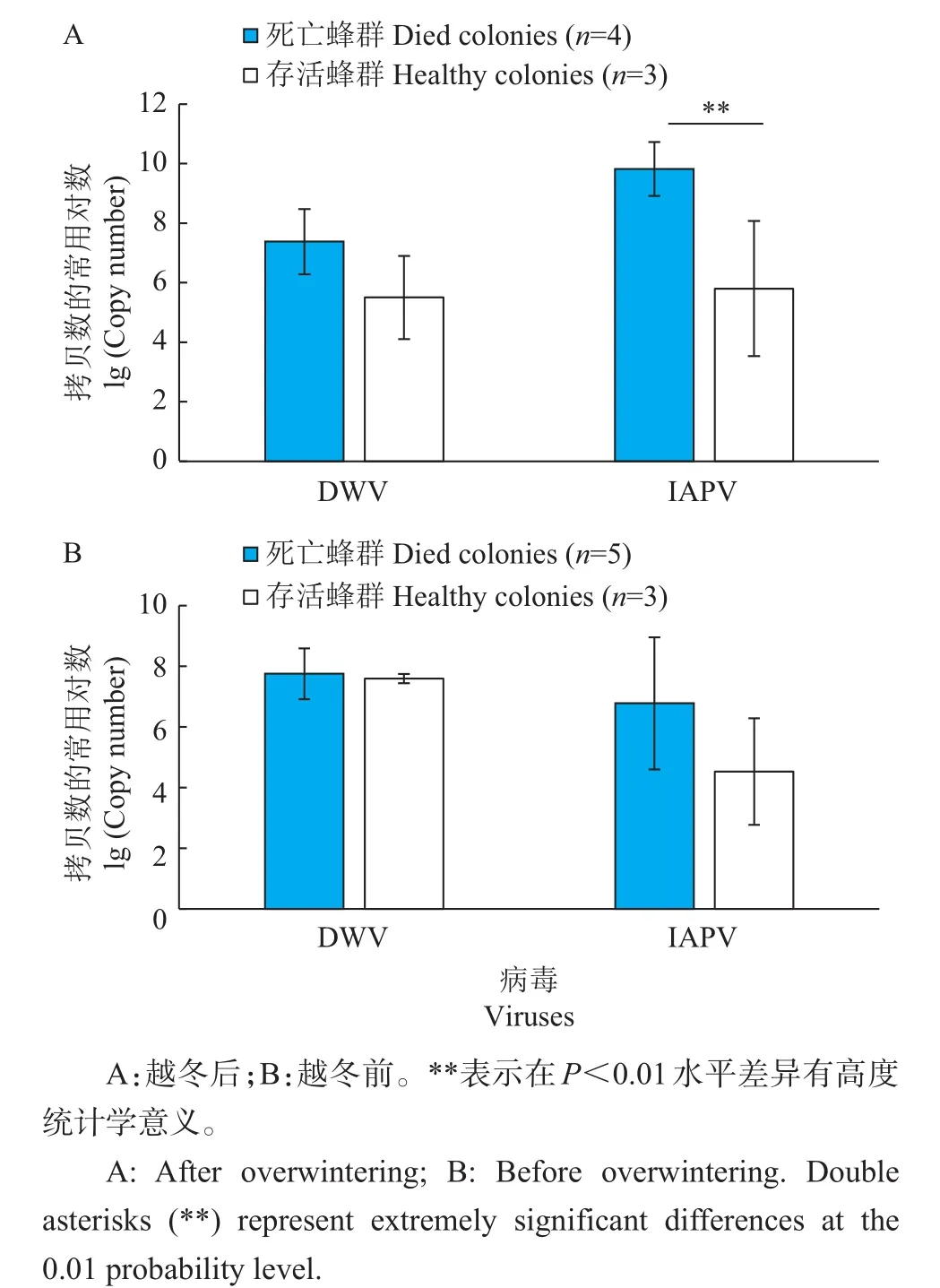
图3 ZJU蜂场死亡蜂群与健康蜂群病毒基因组拷贝数比较Fig.3 Genome copy numbers of viruses in died colonies and healthy colonies in apiary ZJU
2.4 ZJU蜂场蜂群在越冬过程中DWV和IAPV病毒量的变化趋势
对ZJU蜂场8个蜂群越冬期5个时间点的分析结果(图4)表明,在越冬过程中,DWV基因组拷贝数逐渐降低,而IAPV基因组拷贝数呈上升趋势。
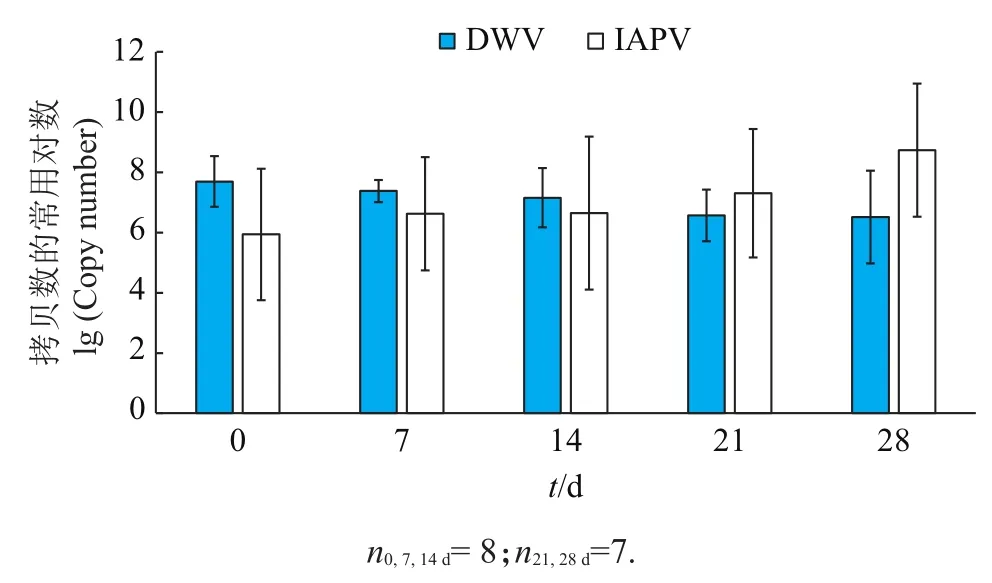
图4 ZJU蜂场蜂群越冬期5个时间点的DWV和IAPV病毒基因组拷贝数变化趋势Fig.4 Genome copy number changes of DWV and IAPV in honey bee colonies from apiary ZJU under five time pointsduring overwintering period
3 讨论
AI等[26]检测了我国18个省份170个西方蜜蜂样本,除KBV未检测到外,DWV、BQCV、SBV、IAPV、ABPV和CBPV都有检测到,以蜂场为单位,感染率分别为94%、44%、21%、18%、6%和9%,在浙江的4个蜂场也未检测到ABPV、CBPV和KBV。在YAÑEZ等[27]的研究中,以蜂场为单位,DWV、IAPV、BQCV和SBV感染率分别为80%、40%、20%和10%。在本研究的3个蜂场中都未检测到的病毒为ABPV、CBPV和SBV,这和上述2项研究中关于这3种病毒在西方蜜蜂上的感染率很低的结论一致。DWV、BQCV和IAPV在3个蜂场中都有检测到,而且这3种病毒都有在某个蜂场某一时间点蜂群感染率达到100%的情况。说明对于越冬意蜂群,这3种病毒是感染率最高的病毒。
本研究所涉及的3个蜂场可以作为3个类型蜂场的典型代表。JH2蜂场蜂群病毒感染率较低,病毒基因组拷贝数也低,健康程度较高。JH1蜂场病毒感染率较高,但在越冬前以毒力较低的BQCV和DWV为主,越冬后DWV和IAPV病毒基因组拷贝数都显著上升。尽管该蜂场蜂群越冬期未出现死亡,但这些蜂群的健康状况堪忧。ZJU蜂场蜂群越冬前BQCV、DWV和IAPV的感染率都很高,从病毒感染情况看,属于健康状态较差的蜂群。这一蜂场出现较高比例(5/8)的越冬蜂群损失,说明整体上病毒感染情况与蜂群越冬期健康有关。
在ZJU蜂场,与越冬后仍健康存活的3个蜂群相比,越冬后取样时濒临死亡蜂群的IAPV基因组拷贝数极显著提高,均值达到1×109以上,且这些蜂群在越冬前IAPV基因组拷贝数也有更高的趋势,达到1×107左右,说明IAPV的高滴度感染与越冬蜂群的死亡有密切关系。在越冬过程中,DWV病毒量呈下降趋势,而IAPV病毒量呈明显上升趋势,进一步说明IAPV是影响越冬蜂健康的重要因素。本研究结果也提示,IAPV感染水平可以作为预测蜂群越冬表现的指标。但由于在本研究中ZJU蜂场样本量较少,因此仍有待进一步研究。
JH1蜂场蜂群越冬后DWV和IAPV病毒量都显著上升,JH2蜂场蜂群越冬后只有DWV病毒量显著上升,而ZJU蜂场蜂群DWV病毒量呈下降趋势,IAPV病毒量呈上升趋势。2种病毒的病毒量在不同蜂场蜂群越冬期间的变化规律不同,说明这2种病毒量的变化和蜂群自身条件存在密切关系,且可能和蜂群越冬表现有关。
蜜蜂病毒的混合感染非常普遍[28],不同病毒间可能存在一定的互作关系。ANDERSON等[29-30]认为,激活KBV可以抑制SBV和BQCV的复制,且KBV与ABPV可能存在竞争关系。IAPV与DWV在某些特定条件下(如ZJU蜂场蜂群健康状态较差时)是否存在竞争抑制关系也值得进一步研究。
综上表明,IAPV是影响越冬期意蜂健康的重要因素。因此,如何降低IAPV感染水平及提高蜜蜂抗IAPV能力是减少蜜蜂越冬期损失所需面对的重要问题。
参考文献(References):
[1]GALLAI N,SALLES J M,SETTELE J,et al.Economic valuation of the vulnerability of world agriculture confronted with pollinated decline.Ecological Economics,2009,68(3):810-821.
[2] CALDERONE N W.Insect pollinated crops,insect pollinators and US agriculture:Trend analysis of aggregate data for the period 1992—2009.PLoS One,2012,7(5):e37235.
[3]BURKLE L A,MARLIN J C,KNIGHT T M.Plantpollinator interactions over 120 years:Loss of species,cooccurrence,and function.Science,2013,339(6127):1611-1615.
[4]LEEK V,STEINHAUERN,RENNICH K,et al.A national survey of managed honey bee 2013—2014 annual colony lossesin the USA.Apidologie,2015,46(3):292-305.
[5] ASTON D.Honey beewinter losssurvey for England,2007-8.Journal of Apicultural Research,2010,49(1):111-112.
[6]PIRK CW W,HUMAN H,CREWE R M,et al.A survey of managed honey bee colony losses in the Republic of South Africa:2009 to 2011.Journal of Apicultural Research,2014,53(1):35-42.
[7] LIU Z G,CHEN C,NIU Q S,et al.Survey results of honey bee(Apis mellifera)colony losses in China(2010—2013).Journal of Apicultural Research,2016,55(1):29-37.
[8] 郑火青,苏晓玲,吉挺,等.从越冬期蜜蜂死亡谈蜜蜂健康养殖.蜜蜂杂志,2016(4):12-15.
ZHENG H Q,SU X L,JI T,et al.How to keep colony health:From the respect of overwintering honey bee death.Journal of Bee,2016(4):12-15.(in Chinese)
[9] 华启云,苏晓玲,赵东绪,等.浙江金华地区越冬意蜂群大量死亡原因分析与应对措施.中国蜂业,2016,67:23-24.
HUA Q Y,SU X L,ZHAO D X,et al.Analysis and countermeasures of the large-scale death of overwintering honey bee colonies(Apis mellifera ligustica)in Jinhua,Zhejiang.Apiculture of China,2016,67:23-24.(in Chinese)
[10]GENERSCH E,VON DER OHE W,KAATZ H,et al.The German bee monitoring project:A long term study to understand periodically high winter losses of honey bee colonies.Apidologie,2010,41(3):332-352.
[11]VANENGELSDORPD,EVANS J D,SAEGERMAN C,et al.Colony collapse disorder:A descriptive study.PLoSOne,2009,4(8):e6481.
[12]RUNCKEL C,FLENNIKEN M L,ENGEL J C,et al.Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses,Nosema,and Crithidia.PLoSOne,2011,6(6):e20656.
[13]MCMENAMIN A J,GENERSCH E.Honey bee colony losses and associated viruses.Current Opinion in Insect Science,2015,8:121-129.
[14]REMNANT E J,SHI M,BUCHMANN G,et al.A diverse range of novel RNA viruses in geographically distinct honey beepopulations.Journal of Virology,2017,91(16):e00158-17.
[15]BONCRISTIANI H F,EVANS J D,CHEN Y P,et al.In vitro infection of pupae with Israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees(Apismellifera).PLoSOne,2013,8(9):e73429.
[16]COX-FOSTER D L,CONLAN S,HOLMES E C,et al.A metagenomic survey of microbes in honey bee colony collapse disorder.Science,2007,318(5848):283-287.
[17]SCHROEDER D C,MARTIN S J.Deformed wing virus:The main suspect in unexplained honeybee deaths worldwide.Virulence,2012,3(7):589-591.
[18]王向辉,郑言,隋佳辰,等.黑蜂王台病毒研究进展.中国畜牧兽医,2016,43(1):248-255.
WANG X H,ZHENG Y,SUI J C,et al.Research advance on black queen cell virus.China Animal Husbandry&Veterinary Medicine,2016,43(1):248-255.(in Chinese with English abstract)
[19]TENTCHEVA D,GAUTHIER L,ZAPPULLA N,et al.Prevalence and seasonal variations of six bee viruses in Apis mellifera L.and Varroa destructor mite populations in France.Applied and Environmental Microbiology,2004,70(12):7185-7191.
[20]TAPASZTI Z,FORGÁCH P,KŐVÁGÓ C,et al.First detection and dominance of Nosema ceranae in Hungarian honeybee colonies.Acta Veterinaria Hungarica,2009,57(3):383-388.
[21]BENJEDDOU M,LEAT N,ALLSOPP M,et al.Detection of acute bee paralysis virus and black queen cell virus from honeybees by reverse transcriptase PCR.Applied and Environmental Microbiology,2001,67(5):2384-2387.
[22]RIBIÈRE M,TRIBOULOT C,MATHIEU L,et al.Molecular diagnosis of chronic bee paralysis virus infection.Apidologie,2002,33(3):339-351.
[23]CHEN Y P,HIGGINS J A,FELDLAUFER M F.Quantitative real-time reverse transcription-PCR analysis of deformed wing virus infection in the honeybee(Apis mellifera L.).Applied and Environmental Microbiology,2005,71(1):436-441.
[24]DI PRISCO G,PENNACCHIO F,CAPRIO E,et al.Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee,Apis mellifera.Journal of General Virology,2011,92(1):151-155.
[25]CHEN Y,ZHAO Y,HAMMOND J,et al.Multiple virus infections in the honey beeand genomedivergence of honey bee viruses.Journal of Invertebrate Pathology,2004,87(2/3):84-93.
[26]AI H X,YAN X,HAN R C.Occurrence and prevalence of seven bee viruses in Apis mellifera and Apis cerana apiaries in China.Journal of Invertebrate Pathology,2012,109(1):160-164.
[27]YAÑEZ O,ZHENG H Q,SU X L,et al.Potential for virus transfer between the honey bees Apis mellifera and A.cerana.Journal of Apicultural Research,2015,54(3):179-191.
[28]GRANBERG F,VICENTE-RUBIANO M,RUBIO-GUERRI C,et al.Metagenomic detection of viral pathogensin Spanish honeybees:Co-infection by aphid lethal paralysis,Israel acute paralysis and lakesinaiviruses.PLoSOne,2013,8(2):e57459.
[29]ANDERSON D L,GIBBSA J.Inapparent virus infections and their interactions in pupae of the honey bee(Apis mellifera Linnaeus)in Australia.Journal of General Virology,1988,69(7):1617-1625.
[30]ANDERSON D L.Kashmir bee virus:A relatively harmless virus of honey bee colonies.American Bee Journal,1991,131(12):767-768.
猜你喜欢
中老年保健(2022年1期)2022-08-17 06:14:30
河北医学(2021年10期)2021-10-27 00:37:14
蜜蜂杂志(2020年2期)2020-12-02 23:34:34
中国蜂业(2020年1期)2020-01-14 01:09:02
中国临床医学影像杂志(2019年6期)2019-08-27 02:59:50
兽医导刊(2019年1期)2019-02-21 01:14:20
中国蜂业(2019年6期)2019-01-09 06:10:25
猪业科学(2018年8期)2018-09-28 01:27:52
中国蜂业(2018年8期)2018-01-20 23:33:56
发明与创新(2015年25期)2015-02-27 10:39:16
