
穿龙薯蓣叶部形态多样性及其与光合产物的关系
2018-05-09 05:59:59温金明石晓东高润梅戎丹仁柳昱旻
华北农学报 2018年2期
温金明,石晓东,高润梅,戎丹仁,柳昱旻
(山西农业大学 林学院,山西 太谷 030801)
1943年,Marker研究发现,薯蓣皂苷元是合成甾体激素药物的原料,薯蓣属(DioscoreaL.)植物的开发利用便在全世界范围内迅速展开,目前,全球甾体激素类药物多于60%以薯蓣皂苷元为原料[1]。我国对薯蓣植物资源的开发利用始于20世纪50年代,国内含薯蓣皂苷元的根状茎植物有17种1亚种3变种,其中,盾叶薯蓣(DioscoreazingiberensisC. H. Wright)和穿龙薯蓣(DioscoreanipponicaMakino)的薯蓣皂苷元含量较高、品质较好[2-7]。盾叶薯蓣是我国的特有种,主要分布于长江流域,在南方多省区已实现产业化,但薯蓣皂苷元产量仍难以满足工业需要,供求矛盾突出[5,8-9]。所以,在我国北方地区发掘与薯蓣皂苷元相同药用成分的其他植物资源,是解决甾体药物原料供应的重要课题。
穿龙薯蓣在我国主要分布于山西、陕西、东北等地区,多生于海拔100~1 700 m的稀疏杂木林内和山腰河谷两侧的山坡灌木丛中及林缘[10]。其根茎入药历史悠久,有调节免疫、改善心血管功能、镇咳、祛痰、平喘等多种药理作用,治疗类风湿性关节炎、冠心病、心绞痛、慢性气管炎、慢性布鲁菌病、脂肪瘤等疗效较好[11]。穿龙薯蓣根茎中薯蓣皂苷元含量为0.93%~2.26%,但受土壤、水分、温度、光照等环境因子的影响,各产地的药材质量存在差异[12-14]。因此,筛选高含量的种质对于穿龙薯蓣的栽培具有重要意义。传统的种质筛选方法是对所有种质皂苷含量进行测定分析,盲目性较大,且费时费力费财,不利于大规模筛选。叶片特征对于盾叶薯蓣种质筛选和保存具有一定的指导意义,根据叶片形态对薯蓣种质进行粗筛,无疑能加快高含量种质筛选的进程。已有研究表明,三角叶薯蓣(DioscoreadeltoideaWall.)的薯蓣皂苷元含量与叶片的长度、宽度呈正相关[15];宽叶型盾叶薯蓣的薯蓣皂苷元含量较长叶型植株含量高[12,16];盾叶薯蓣叶的叶绿素含量与光合速率正相关,光合速率越强,光合产物含量越大,根状茎产量越高,薯蓣皂苷元含量越高[17]。
本试验以山西省蟒河和关帝山2个种源地的穿龙薯蓣植株为研究对象,分析穿龙薯蓣叶片的形态多样性特征,比较不同叶形的叶绿素与光合产物的含量,以期探讨叶形态与光合产物间的相关性,为穿龙薯蓣的良种选育工作提供基础资料。
1 材料和方法
1.1 试验材料
供试种源分别取之于山西省蟒河自然保护区和关帝山庞泉沟自然保护区。
1.2 试验地概况
蟒河自然保护区(112°22′10″~112°31′35″E,35°12′30″~35°17′20″N)位于晋城市阳城县境内,海拔300~1 572 m,年均降水量600~900 mm,年均温14 ℃,无霜期180 d,属暖温带半湿润大陆性季风气候[18]。
关帝山庞泉沟自然保护区(37°20′~38°20′N,110°18′~111°18′E)位于吕梁山中段,处于交城、方山两县之间,海拔1 300~2 800 m,年均降水量约820 mm,年均温4.3 ℃,无霜期180 d,属暖温带半湿润大陆性季风气候[19]。
1.3 试验方法
2014年9-10月分别于试验种源地采集穿龙薯蓣部分地下根茎,沙藏越冬至2015年4月,取出根茎植于山西农业大学林学院苗圃,常规水肥管理。2017年5月,选取2个种源地长势旺盛的穿龙薯蓣植株,按照叶色、叶裂数等叶部形态的差异,选取180枚成熟叶片,进行叶形态测量和生理指标测定。
1.4 测定项目及方法
1.4.1 叶形态指标测量 采用EPSON Scan扫描仪获取叶片的数字图像,利用 Image J软件分别测量叶长(叶基到叶尖的长度)、叶宽(垂直于叶柄方向的叶片最宽处)、叶面积和叶张角(叶基部夹角)[20]。人工数取叶裂数,计算叶片长宽比。为了分析的便利,对叶张角进行分级:Ⅰ. 叶张角<40°;Ⅱ. 40°≤叶张角<50°;Ⅲ. 50°≤叶张角<60°;Ⅳ. 60°≤叶张角<70°;Ⅴ. 叶张角≥70°。
1.4.2 叶生理指标测定 叶绿素含量测定采用直接浸提法[21],可溶性蛋白质含量测定采用考马斯亮蓝染色法[22],可溶性糖和淀粉含量的测定采用蒽酮比色法[22],还原糖含量的测定采用3,5-二硝基水杨酸法[23]。
1.5 数据统计与分析
采用统计软件SPSS 19.0对数据进行方差分析和相关分析,差异水平为0.05;采用Excel 2013进行图表制作。
2 结果与分析
2.1 穿龙薯蓣叶形态多样性分析
测量计算穿龙薯蓣的叶长、叶宽、长宽比、叶面积、叶裂数和叶张角等形态指标,并就叶形态差异和指标之间的相关性进行分析。
2.1.1 叶形态差异 穿龙薯蓣叶形态多样性明显,表现在叶色深浅、叶长、叶宽、叶裂、叶张角等方面,结果如表1所示。
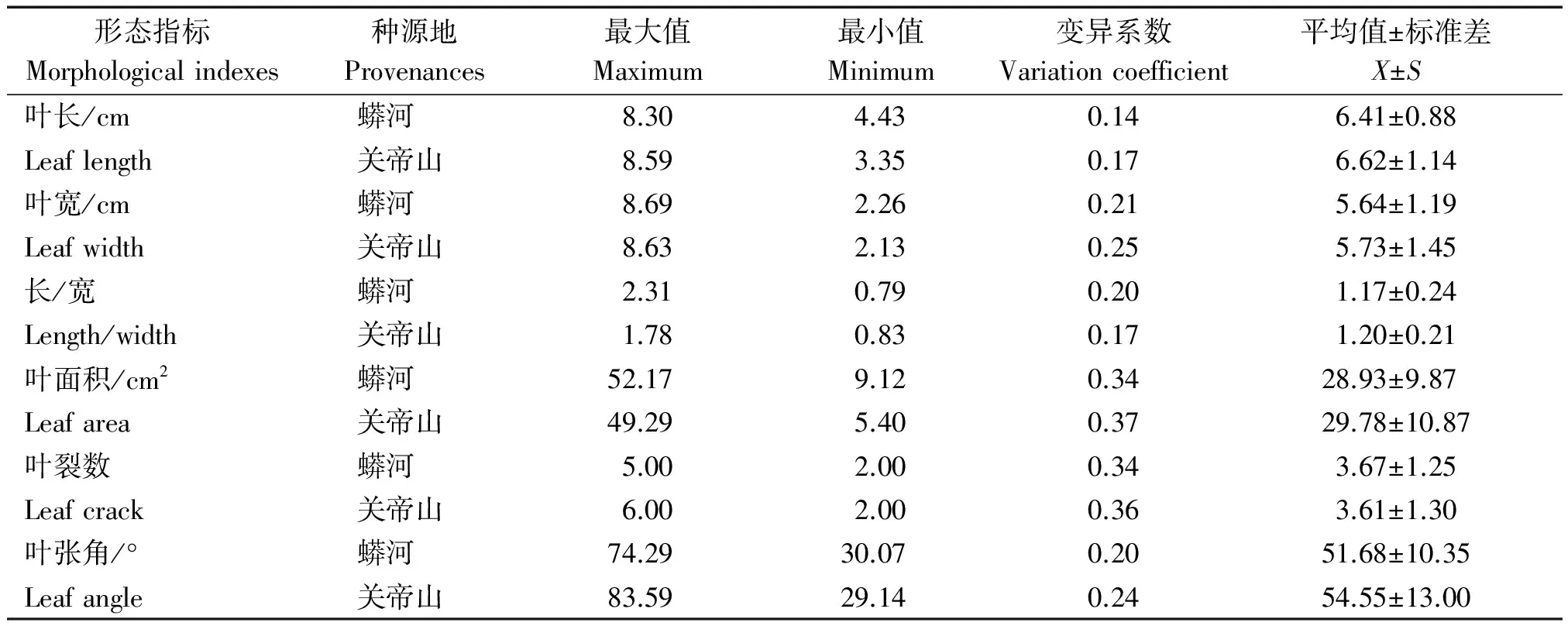
表1 穿龙薯蓣叶形态差异 Tab.1 Leaf morphological differences of Discorea nipponica Makino
由表1可知,就2个种源地比较而言,除了长宽比之外,关帝山种源地穿龙薯蓣叶片的叶长、叶宽、叶面积、叶裂数和叶张角的变异系数均大于蟒河种源地,即关帝山种源地的叶形态变异大于蟒河种源地。2个种源地都存在2种叶型:长叶型(叶片长度大于宽)和宽叶型(叶片宽度大于长度):蟒河种源的叶长为4.43~8.30 cm,叶宽为2.26~8.69 cm,长宽比为0.79~2.31;关帝山种源的叶长为3.35~8.59 cm,叶宽为2.13~8.63 cm,长宽比为0.83~1.78。叶片大小差异直接造成叶面积的明显差异:蟒河种源地叶面积为9.12~52.17 cm2,关帝山种源地5.40~49.29 cm2,变异系数高达0.34和0.37。此外,叶形态多样性在叶裂数和叶张角方面也有体现:2个种源地的叶裂数变异系数分别为0.34和0.36;叶张角差异明显,关帝山种源地多于蟒河种源地近3°。
进一步统计分析穿龙薯蓣2个种源地不同叶型和各级叶张角所占的比例,结果如图1所示。
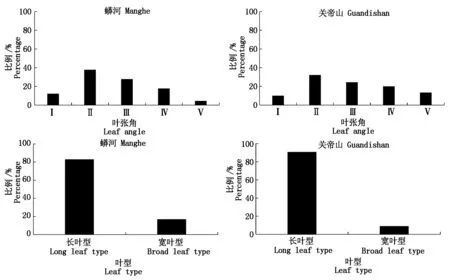
图1 两个种源地的叶型和叶张角比例 Fig.1 Percentage of leaf type and leaf angle of two provenances
由图1可知,就2个种源地的叶型均以长叶型居多,蟒河种源长叶型占83%,关帝山种源长叶型占91%。就叶张角来看,2种源均以第Ⅱ级(40°~50°)比例最大,蟒河种源占38%,关帝山种源占32%;Ⅲ、Ⅳ级次之,即多数叶片的叶张角为40°~70°。蟒河种源Ⅱ、Ⅲ、Ⅳ级叶张角占总叶数的83%,关帝山占77%;关帝山种源有>10%的叶片叶张角>70°,蟒河种源有<10%叶片叶张角>70°,但约有12%的叶片叶张角<40°。
统计分析不同叶色的叶型与各级叶张角的比例,结果如图2所示。
由图2可知,2种叶色的叶片均以长叶型占据优势,深色叶中,长叶型占80%,浅色叶中,长叶型占比大于深色叶片,占96%。就叶张角来看,深色叶片中各级叶张角占比接近,Ⅰ~Ⅳ级占89%;浅色叶片中叶张角比例差异较大:第Ⅱ级比例最高,占40%;Ⅱ~Ⅳ级比例占90%,即多数叶片的叶张角为40°~70°;Ⅰ级占比最少,为3%。
2.1.2 叶形态指标间的相关性 对穿龙薯蓣叶片的叶色、叶长等形态指标进行相关性分析,结果如表2所示。
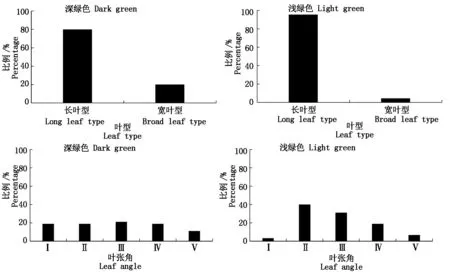
图2 不同叶色的叶型和叶张角比例 Fig.2 Percentage of leaf type and leaf angle of different leaf color
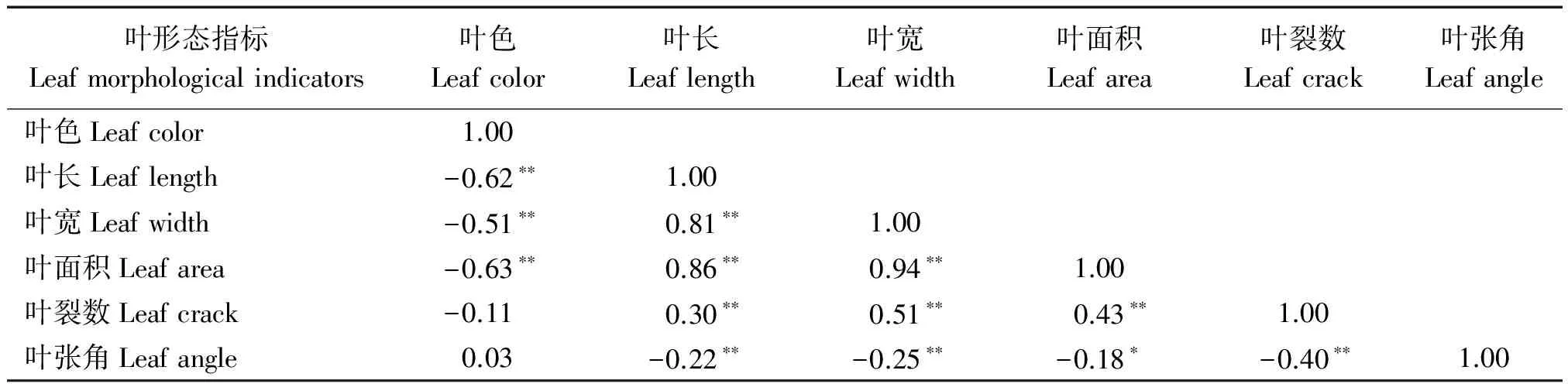
叶形态指标Leafmorphologicalindicators叶色Leafcolor叶长Leaflength叶宽Leafwidth叶面积Leafarea叶裂数Leafcrack叶张角Leafangle叶色Leafcolor1.00叶长Leaflength-0.62∗∗1.00叶宽Leafwidth-0.51∗∗0.81∗∗1.00叶面积Leafarea-0.63∗∗0.86∗∗0.94∗∗1.00叶裂数Leafcrack-0.110.30∗∗0.51∗∗0.43∗∗1.00叶张角Leafangle0.03-0.22∗∗-0.25∗∗-0.18∗-0.40∗∗1.00
注:**.在0.01水平上显著相关;*.在0.05水平上显著相关。表3-4同。
Note:**.The significant correlation at the level of 0.01;*.The significant correlation at the level of 0.05. The same as Tab.3-4.
由表2可知,叶长与叶宽、叶面积、叶裂数呈极显著正相关,与叶张角呈极显著负相关;叶宽与叶面积、叶裂数呈极显著正相关,与叶张角呈极显著负相关;叶面积与叶裂数呈极显著正相关,与叶张角呈显著负相关;叶裂数与叶张角呈极显著负相关;叶色与叶长、叶宽、叶面积呈极显著负相关。通过以上分析可知,穿龙薯蓣叶部形态指标间显著相关,均可用作穿龙薯蓣种质资源筛选。
2.2 叶绿素含量分析
对穿龙薯蓣叶片的叶绿素含量进行测定,并进行各形态指标不同类型的含量比较,结果列于表3。
由表3可知,穿龙薯蓣叶片叶绿素含量与种源地、叶型相关性不显著,与叶色相关性极显著。穿龙薯蓣叶片的叶绿素含量在不同种源地和叶型之间无显著差异,在不同叶裂数间存在显著差异。深绿色叶片的Chla、Chlb和ChlT含量的平均值均高于浅绿色叶片,但Chla/Chlb的平均值小于浅绿色叶片。叶5裂的叶片中,Chla和ChlT含量平均值最高,显著高于其他3,4裂的叶片;叶6裂的叶片中,Chla/Chlb的平均值最高,为8.62,显著高于2,3,5裂的叶片。叶绿素含量在不同类型之间叶张角的差异较小。
2.3 叶片光合产物含量分析
对穿龙薯蓣叶片的光合产物含量进行测定,结果如表4所示。
由表4可知,穿龙薯蓣叶片的可溶性蛋白质、可溶性糖和还原糖含量与种源地和叶型的相关性不显著,与叶色的相关性极显著。穿龙薯蓣叶片的淀粉含量与种源地相关性极显著,与叶色相关性显著,与叶型相关性不显著。穿龙薯蓣深绿色叶片的4种光合产物含量的平均值都大于浅绿色叶片。2裂叶片的可溶性蛋白质和还原糖含量显著高于其他裂数叶片,可溶性糖含量显著高于4裂、5裂和6裂的叶片,淀粉含量显著高于4裂和5裂的叶片。不同类型叶张角叶片除了淀粉,其他3种光合产物含量均差异显著,含量的整体变化趋势是随叶张角的增大,光合产物含量显著增加。
进一步分析相关性强弱,结果如图3所示。
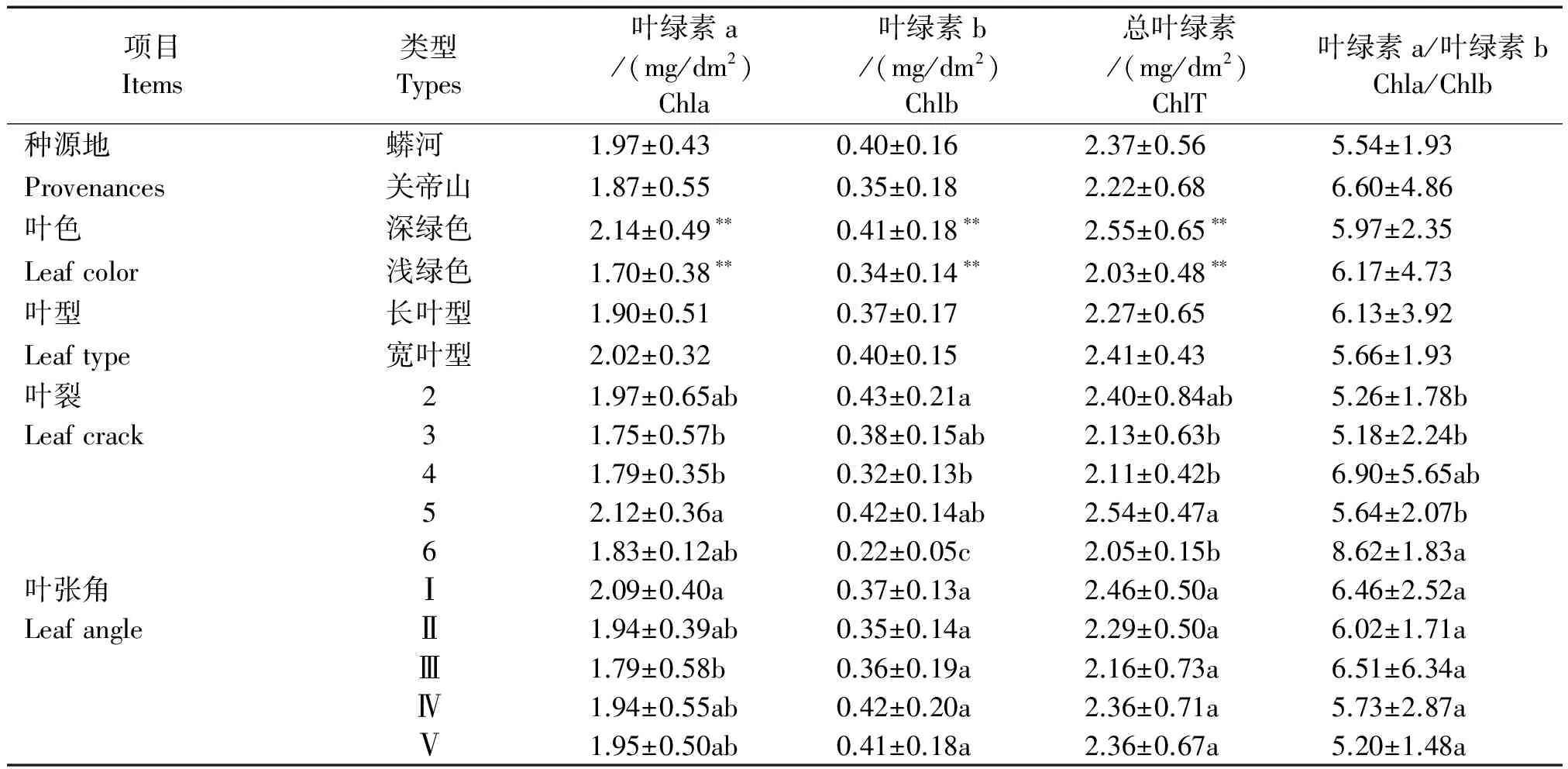
表3 穿龙薯蓣叶绿素含量 Tab.3 Chlorophyll content of Discorea nipponica Makino
注:同一列不同小写字母示差异显著(P<0.05)。表4同。
Note:Different letter in the same column indicate significant difference (P<0.05). The same as Tab.4.
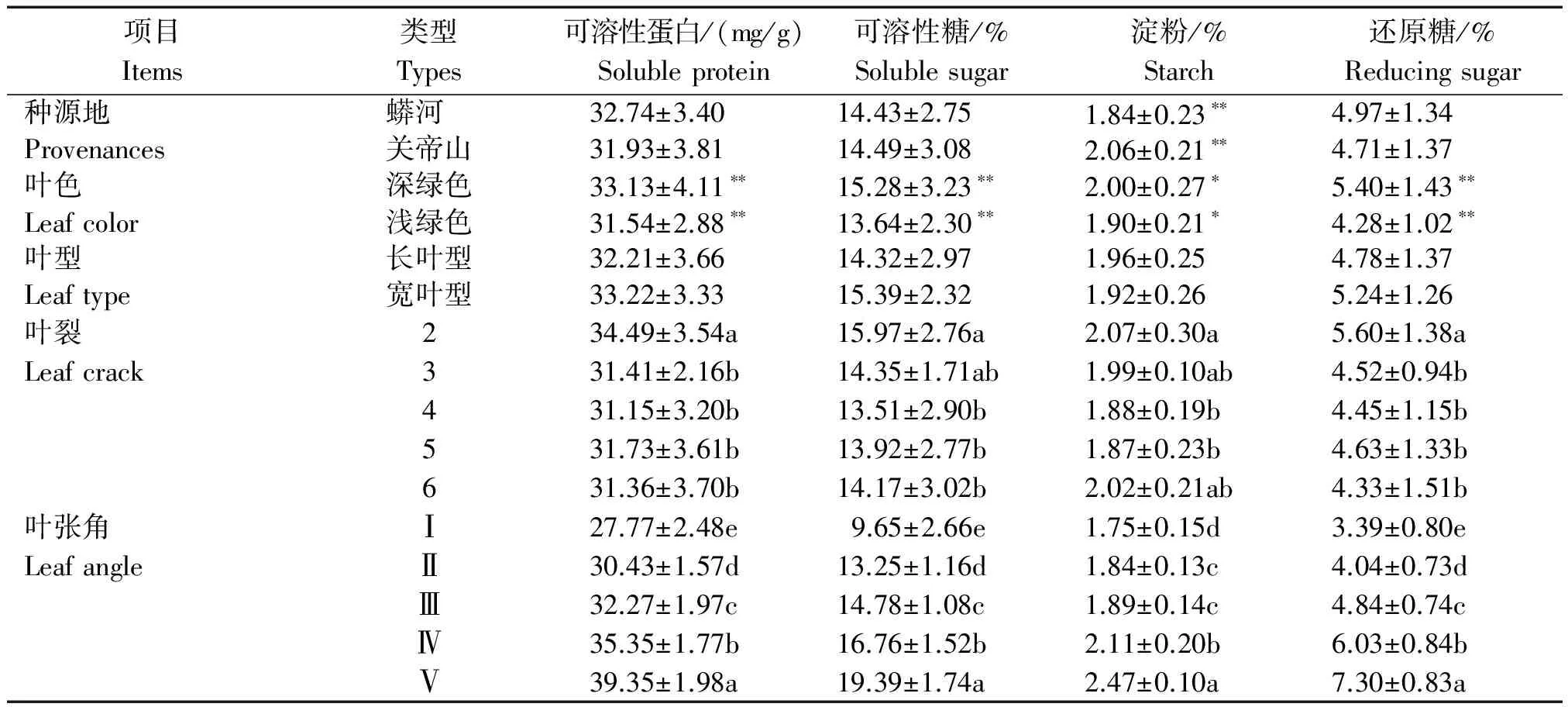
表4 穿龙薯蓣光合产物含量 Tab.4 Photosynthate content of Discorea nipponica Makino
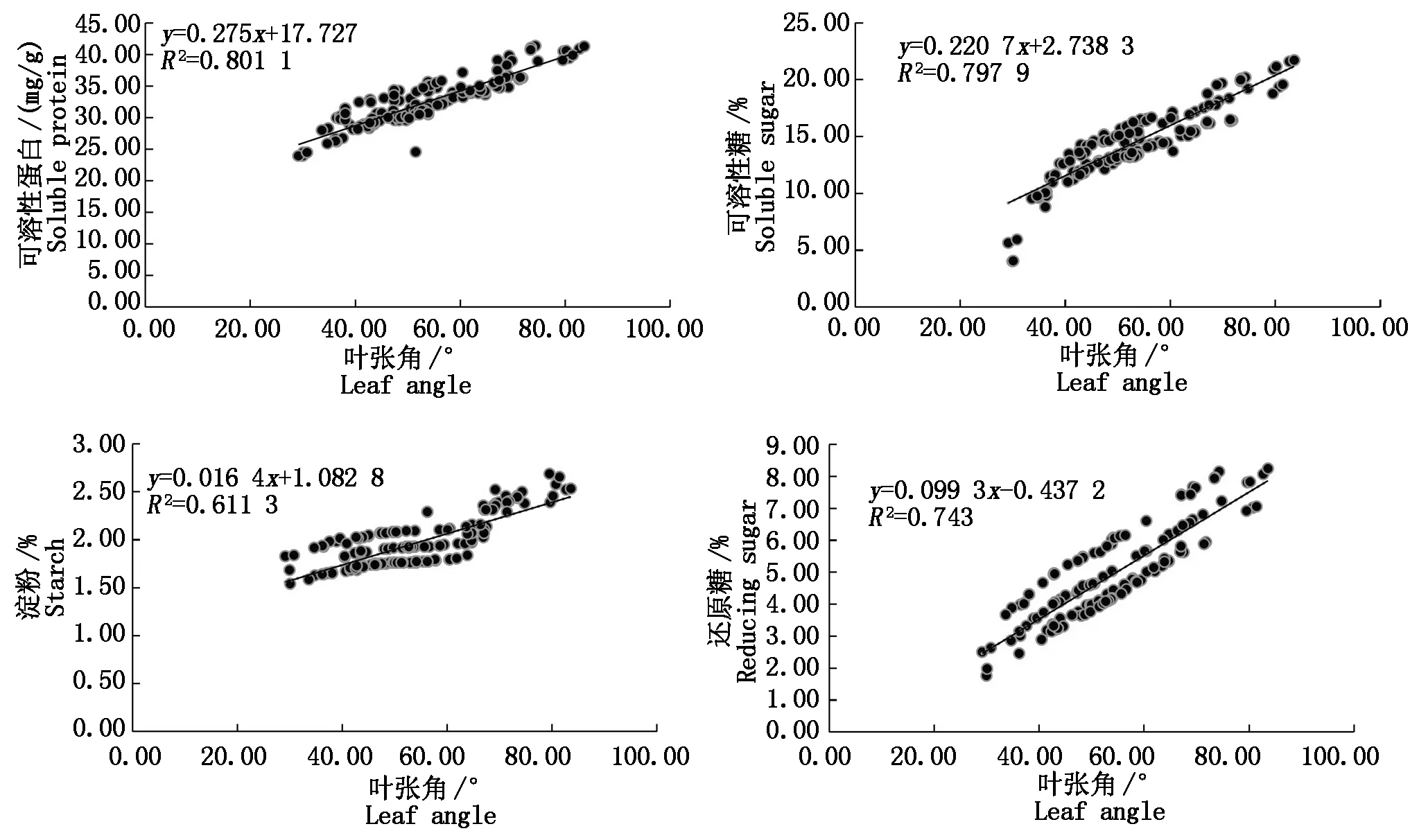
图3 叶张角与光合产物含量的相关性Fig.3 Correlation between leaf angle and photosynthate content
由图3可知,穿龙薯蓣叶张角与叶片内光合产物的含量显著相关,但不同产物的相关性强弱不同,依次为可溶性蛋白>可溶性糖>还原糖>淀粉。
3 讨论与展望
3.1 薯蓣属植物叶形态多样性情况
薯蓣科中薯蓣属种类最多,共6组600种,多数种叶部形态多样,以盾叶薯蓣和穿龙薯蓣为甚。其中,盾叶薯蓣叶形为三角状卵形、心形或箭形,通常3浅裂至3深裂,叶脉掌状分布[24];穿龙薯蓣叶片卵圆形,叶色有深绿、浅绿二色,常呈2~6浅裂或深裂。此外,不同分布区的薯蓣属植物在叶形态也存在差异[25]。本研究根据长宽比可以把薯蓣属植物的叶片分成两大类型:长叶型和宽叶型。穿龙薯蓣存在2种叶型,2个种源地叶型都以长叶型为主,关帝山种源的长叶型比例高于蟒河种源。多数穿龙薯蓣的叶张角为40°~70°。深绿色叶片多为厚纸质,浅绿色叶片多为薄纸质。叶形态指标之间的差异反映了穿龙薯蓣叶部形态特征的多样性,表明穿龙薯蓣是一个叶形态“多型性”的种。此外,穿龙薯蓣叶部形态指标间显著相关,均可用作穿龙薯蓣种质资源筛选。
3.2 叶形态与有效成分间的关系
植物所处的生态环境及丰富的形态多样性可能与有效成分之间存在关系,为进一步分析穿龙薯蓣不同种源地、不同类型叶片与其地下根状茎有效成分薯蓣皂苷元含量之间的关系,本研究先对2个种源地不同形态叶片的光合产物含量进行了测定。植株体内的可溶性蛋白质含量的高低间接反映各种代谢活动的强弱,植物产品的品质性状,常以糖含量作为重要指标。可溶性糖是总糖的一个重要组成部分,是植物碳水化合物代谢中重要的运输形式,可溶性糖含量的高低,反映了植株体内可利用物质和能量的供应基础[26-27]。淀粉作为营养性多糖,在淀粉酶作用下可以水解为可溶性糖[28]。还原糖是指可被氧化充当还原剂的糖。本研究表明,穿龙薯蓣叶张角与叶片内可溶性蛋白质、可溶性糖、淀粉、还原糖含量显著相关,含量的整体变化趋势是随叶张角的增大,叶片的这4种光合产物含量增大。说明叶张角可用作穿龙薯蓣种质资源筛选时首选的叶部形态指标。
穿龙薯蓣叶部形态、叶绿素含量和光合产物含量产生差异的原因有两方面:一是生境的多样性:叶形态的差异是对不同生境条件的长期适应造成的;蟒河和关帝山2个分布区的海拔、林下光照、降水量、日平均气温、土壤类型等立地条件不同,使穿龙薯蓣产生了与之相适应的形态类型。二是遗传的多样性:薯蓣属植物的染色体数目变异较大,且倍性复杂[29-30];有研究结果显示,怀山药和大薯形态特征丰富,染色体数目为40条或60条,叶形态特征与染色体数目存在一定的相关性[31-32]。
3.3 穿龙薯蓣研究的展望
穿龙薯蓣叶部形态存在多样性,为选种栽培提供了丰富的资源。根据叶形态指标进行粗筛,这可以减少筛选的盲目性,提高准确度,缩短野外筛选的时间,大大加快高质量种质筛选的进程,对于野生穿龙薯蓣资源中高薯蓣皂苷元含量的种质筛选、保护和繁殖有着重要意义,对解决甾体药物原料供应有着现实的经济价值。但基于多样性研究,尤其是遗传多样性的研究方面起步较晚,今后应加强以下几方面的研究:一是应用常规研究方法与先进的生物学技术相结合,分析穿龙薯蓣皂苷元含量与多样性特征之间的关系,研究和评价穿龙薯蓣资源的多样性;二是应用DNA分子标记的方法比较和分析高皂苷元含量类型的特征性基因片段加强遗传多样性的研究;三是加强穿龙薯蓣代谢过程的研究,探索穿龙薯蓣药用部分(地下茎)生长规律与薯蓣皂苷元积累的动态关系,设法对穿龙薯蓣代谢过程进行改造,使之具有最佳薯蓣皂苷元产生模式;四是对薯蓣皂苷元含量高的穿龙薯蓣品种,进行形态、生理和DNA水平等方面的研究,从中筛选更优良的品种,运用组织培养等方法进行扩大利用[17]。
参考文献:
[1] Ren Y,Chen Y,Hu B,et al. Microwave-assisted extraction and a new determination method for total steroid saponins fromDioscoreazingiberensisC.H. Wright[J]. Steroids,2015,104:145.
[2] 张新新. 盾叶薯蓣的化学成分及其药理作用研究[D].西安:西北大学, 2015.
[3] 雷 震, 魏晋宝, 杨光义, 等. 盾叶薯蓣中薯蓣皂苷元的含量测定与提取方法研究[J]. 河北医药, 2015(17): 2671-2675.
[4] 聂凌鸿,林淑英,宁正祥. 薯蓣属植物中薯蓣皂苷元的研究进展[J]. 中国生化药物杂志,2004,25(5):318-320.
[5] 李开泉,邹盛勤,陈 武. 薯蓣属植物的研究开发现状[J]. 林产化工通讯,2004,38(2):26-29.
[6] Yuan Y,Zhang L M,Dal Y,et al. Physicochemical properties of starch obtained fromDioscoreanipponicaMakino comparison with other tuber starches[J]. Journal of Food Engineering,2007,82(4):436-442.
[7] 黄含含,李 霞,高文远,等. 薯蓣属药用植物的亲缘关系研究[J]. 中国中药杂志,2015,40(17):3470-3479.
[8] 滕文锐, 陈俊英. 盾叶薯蓣皂素提取及其资源综合利用研究进展[J]. 科技广场, 2014(12): 251-256.
[9] 汪 朋. 盾叶薯蓣中甾体皂苷的提取和薯蓣皂苷元绿色制备工艺研究[D].无锡:江南大学, 2015.
[10] 侯明明. 山西省野生穿龙薯蓣所处群落分类、组成及种间关系研究[D].晋中:山西农业大学, 2016.
[11] 蒋玉宝,张素梅,于元杰. 穿龙薯蓣根的诱导和快繁及薯蓣皂苷元含量的测定[J]. 华北农学报,2007,22(S):189-192.
[12] 李朝阳,杨朝霞,陈功锡,等. 不同居群盾叶薯蓣生长、叶形态及皂苷元含量的研究[J]. 生命科学研究,2003,7(1):58-64.
[13] 贾晓晴, 尹海波, 陶 玲, 等. 不同产地穿山龙活性成分与环境因子相关性研究[J]. 辽宁中医杂志, 2017(4): 820-824.
[14] 姚 佳, 尹海波, 赵 容, 等. 基于连作障碍条件下穿山龙无机元素吸收规律及总皂苷含量分析[J]. 中国实验方剂学杂志, 2015(24): 61-64.
[15] Bindroo B,Bhat B K,1985. Correlation studies inDioscoreadeltoideaWall[J]. Crop Improvement,1998,12(1):49-51.
[16] 杭悦宇,夏 冰,周义峰,等. 武当山地区盾叶薯蓣薯蓣皂苷元含量及生物特性的相关性研究[J]. 林产化学与工业,2004,24(4):68-72.
[17] 薛 焱. 不同居群及株系盾叶薯蓣的形态、生理特性研究[D]. 武汉:华中农业大学,2005:23,35.
[18] 高润梅,石晓东,樊兰英,等. 山西省南方红豆杉自然分布与群落生态学特征[J]. 应用生态学报,2016,27(6):1820-1828.
[19] 陈顺治. 关帝山华北落叶松林下植物多样性和土壤碳氮磷的海拔梯度格局[D].晋中:山西农业大学, 2016.
[20] 戴志聪,杜道林,司春灿,等. 用扫描仪及Image J软件精确测量叶片形态数量特征的方法[J]. 广西植物,2009,29(3):342-347.
[21] 舒 展,张晓素,陈 娟,等. 叶绿素含量测定的简化[J]. 植物生理学报,2010,46(4):399-402.
[22] 李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:94-195.
[23] 白雪媛,赵 雨,刘海龙,等. 不同产地人参中的总糖,还原糖和可溶性多糖含量的比较研究[J]. 中国现代应用药学,2012,29(1):39-42.
[24] 梁艳丽, 赵庆云, 杨 燕, 等. 盾叶薯蓣种质资源比较研究[J]. 中国农学通报, 2014, 30(36):110-114.
[25] 《山西植物志》编辑委员会. 太原:山西植物志[M]. 5卷. 北京: 中国科学技术出版社, 2004: 446-449.
[26] 李 勇. 山西不同种源穿龙薯蓣抗旱性及皂苷元含量分析[D].晋中:山西农业大学, 2016.
[27] 曾光辉,周 琳,黎星辉. 自然越冬期间茶树叶片生理生化指标和解剖结构的变化[J]. 植物资源与环境学报,2017,26(1):63-68.
[28] 吕文河, 霍丹丹, 李 勇, 等. 马铃薯块茎不同部位淀粉含量及淀粉合成关键酶活性差异比较[J]. 东北农业大学学报, 2017, 48(6): 1-8.
[29] 黄和平, 高山林, 黄璐琦, 等. 同源四倍体盾叶薯蓣遗传稳定性研究[J]. 中药材, 2014, 37(3):365-368.
[30] 夏 薇, 吴文嫱, 许 云, 等. 大薯种质资源多样性研究进展[J]. 植物生理学报, 2017,53(5):781-784.
[31] 尹海波,张 囡,康廷国. 穿龙薯蓣药材和土壤无机元素的相关性分析[J]. 中国实验方剂学杂志,2011,17(19):137-141.
[32] 吴文嫱,王晓倩,翟李楠,等. 薯蓣属植物遗传多样的研究进展[J]. 热带农业科学,2015,35(10):29-38.
猜你喜欢
航空发动机(2022年4期)2022-10-13 04:51:40
东方汽轮机(2022年1期)2022-04-13 02:25:00
节能技术(2022年1期)2022-03-18 11:57:28
北方文学(2019年21期)2019-08-18 15:23:25
中成药(2017年9期)2017-12-19 13:34:31
中成药(2017年9期)2017-12-19 13:34:28
小雪花·初中高分作文(2017年4期)2017-06-29 07:49:48
水利规划与设计(2017年12期)2017-02-06 03:39:50
西南农业学报(2016年5期)2016-05-17 05:42:46
医学研究杂志(2015年8期)2015-06-22 14:00:57
