
不同番茄品种对镉胁迫的耐抗性差异研究
2018-05-09 05:59:58高夕彤杨志新
华北农学报 2018年2期
高夕彤,李 硕,贾 娟,李 岩,杨志新
(河北农业大学 资源与环境科学学院,河北省农田生态环境重点实验室,河北 保定 071001)
中国是一个农业大国,伴随着农药、化肥施用量在农业上的不断增加以及不合理的耕作施肥方式导致了农田土壤重金属污染日益严重。因Cd的生物有效性高,易在农作物中积累[1],Cd污染超标在农产品重金属超标中已位于首位[2-3]。近20年来,有关Cd对植物的毒害作用及耐性机理已有较多研究,如Cd通过阻碍植物根系生长、抑制水分和养分的吸收等引起一系列生理代谢紊乱[4],当镉进入植物体内并积累到一定程度时,就会通过影响植物的生长发育、抑制植物的呼吸作用和光合作用、减弱植物体中的酶活性、降低植物可溶性蛋白和可溶性糖的含量等途径来影响植物的产量、品质和安全[5-7]。Cd还能抑制植物对其他营养元素的吸收,间接影响植物的生长发育[8]。Cd生物毒性强,超过一定限度不仅影响作物品质和产量,而且会通过食物链危害人体健康[9],摄入过量的镉会对人体的免疫系统、泌尿系统、骨骼、神经系统、生殖系统等造成损伤,同时镉还具有较强的致癌、致畸、致突变的作用[10-11]。目前,Cd污染已成为限制我国蔬菜质量和发展无公害蔬菜产品的主要因素之一。番茄是人类日常生活中大量食用的蔬菜,其品质的好坏直接影响人们的日常食品安全,目前关于筛选番茄品种Cd吸收差异的研究已有报道,但对不同品种的耐抗性资源积累尚不充分,应予以加强,抗Cd番茄品种资源的筛选对于农业土壤重金属修复的种植结构调整具有理论指导意义,同时对番茄品种合理使用、发展绿色无公害蔬菜、保护人类健康具有重要的现实意义。
1 材料和方法
1.1 供试材料
1.1.1 供试蔬菜 供试材料为番茄幼苗,前期通过温室盆栽及实验室水培优选确定的4种不同抗性番茄(LycopersiconesculentumMill) 品种,为普罗旺斯、瑞提娜、凯特2号和908,前3种购于河北省永清县振兴苗厂,后者购于河北农业大学附近的集贸市场。
1.1.2 供试营养液 供试营养液为1/2霍格兰德营养液(pH值6)[12],其组成列表于表1。
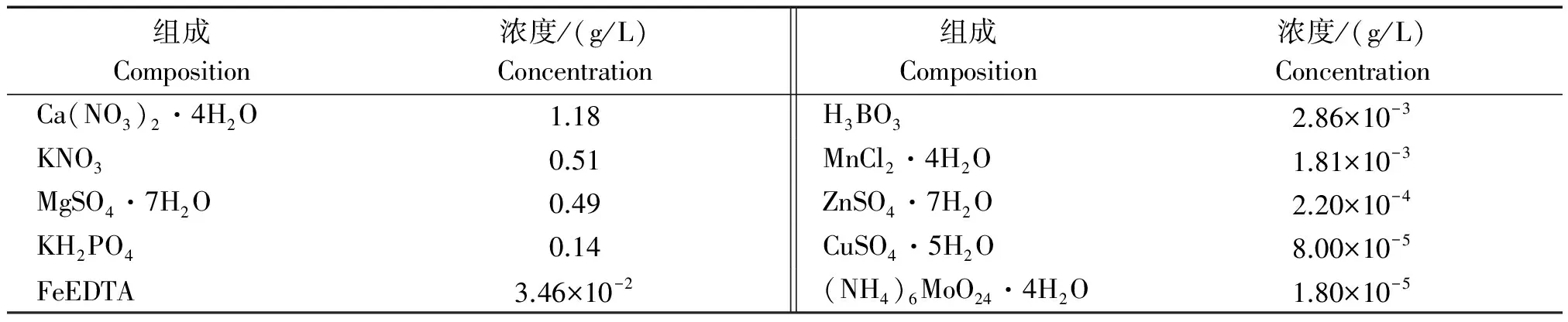
表1 霍格兰德营养液组成 Tab.1 Hoagland nutrient composition
1.2 试验设计
试验于2017年4-5月在河北农业大学资环学院温室内进行。选择长势良好且均匀的幼苗作为试验材料,用去离子水浸洗除去根部营养土。以5株番茄幼苗为一组,移至装有1 L营养液(1/2浓度)的水培罐中,用海绵固定。Cd源为分析纯 CdCl2·2.5H2O,处理浓度分别为0.0,0.3,1.0,1.5 mg/kg。水培期间,25 ℃室温下光照(12 h/d),保持通风,培养10 d测定番茄植株生长指标和抗性指标,以此研究不同品种对Cd的耐性差异。
1.3 样品采集和测定
采集处理10 d后的所有植株样品,用EDTA浸泡除去样品根系表面吸附的Cd后,再用蒸馏水洗净擦干,分离地上部和根系,量取株高和根长。样品105 ℃杀青0.5 h后于65 ℃下烘干后称取干质量并粉碎。植物样品Cd含量采用高压消解-电感耦合等离子体质谱法测定;丙二醛(MDA)含量用 TBA(硫代巴比妥酸)法测定;叶绿素含量采用乙醇提取-分光光度法测定;过氧化物酶(POD)活性用愈创木酚法测定;过氧化氢酶(CAT)活性采用紫外吸收法测定。
1.4 统计处理
采用Excel 2010和SPSS 19.0进行数据处理,LSD法进行差异性比较。
2 结果与分析
2.1 在无镉处理下不同番茄品种生长及其抗性指标的差异特征
在不添加Cd的营养液培养条件(CK)下,番茄幼苗生长10 d后的株高、根长和生物量指标特征见表2。从表2看出,在CK处理下,品种908的株高、根长和生物量总干质量均表现出了强劲的生长优势,3项指标均远高于其他品种;普罗旺斯的根长显著高于瑞提娜和凯特2号,而株高和总生物量却显著低于瑞提娜和凯特2号;瑞提娜和凯特2号2个品种的根长和总生物量指标水平差异不显著,且凯特2号的株高显著高于瑞提娜。可见,不同番茄品种间生长指标及生物量的差异较为显著,反映出了不同番茄品种生物遗传属性的自然差异特征。
在不添加Cd处理下进一步测定了番茄幼苗中丙二醛(MDA)、叶绿素、过氧化物酶(POD)和过氧化氢酶(CAT)活性指标,反映了各品种的自然特性(表3)。结果表明,在CK处理下,不同番茄品种幼苗的MDA、叶绿素含量以及POD、CAT活性均存在较大差异;丙二醛含量在4个品种间有显著的差异性,表现为908≈普罗旺斯>凯特2号≈瑞提娜,908和普罗旺斯表现最高;叶绿素含量大小表现为908>凯特2号>普罗旺斯≈瑞提娜,908表现最高;POD活性为瑞提娜>普罗旺斯≈凯特2号≈908,瑞提娜表现最高;CAT活性为普罗旺斯>凯特2号≈908>瑞提娜,普罗旺斯表现最高。总体来看,各指标在品种间有不同程度的变化特征,进一步证实了不同番茄品种抗性指标生物遗传属性的差异性。
2.2 镉胁迫下不同番茄品种生长指标的变化特征
表2反映了Cd胁迫处理培养10 d后各番茄品种3项指标的测定结果,结果表明,与CK相比较,低浓度Cd(0.3 mg/kg)胁迫对普罗旺斯根长有显著刺激作用(P<0.05),使其增长幅度达24.72%,株高和总干质量虽然也呈现出一定程度的增长趋势(3.09%和14.21%),但与对照差异并不显著;对908根长有显著的抑制作用(P<0.05),降幅为12.47%;对其余品种的各项指标均有一定程度的抑制影响,但均未达显著性影响水平。可见,低镉浓度对普罗旺斯生长有一定的促进作用,尤其根系反应更加敏感,而对其余品种影响不明显。
在中等浓度Cd(1.0 mg/kg)和高浓度Cd(1.5 mg/kg)胁迫下,4个品种的所有指标均比对照有一定程度的降低,其中瑞提娜株高达到了显著降低水平(P<0.05),降幅分别为18.28%和17.01%。说明在中、高Cd含量胁迫下,瑞提娜在生长上受到了显著抑制,抗性表现最弱,而对其余品种的生长指标影响不大。
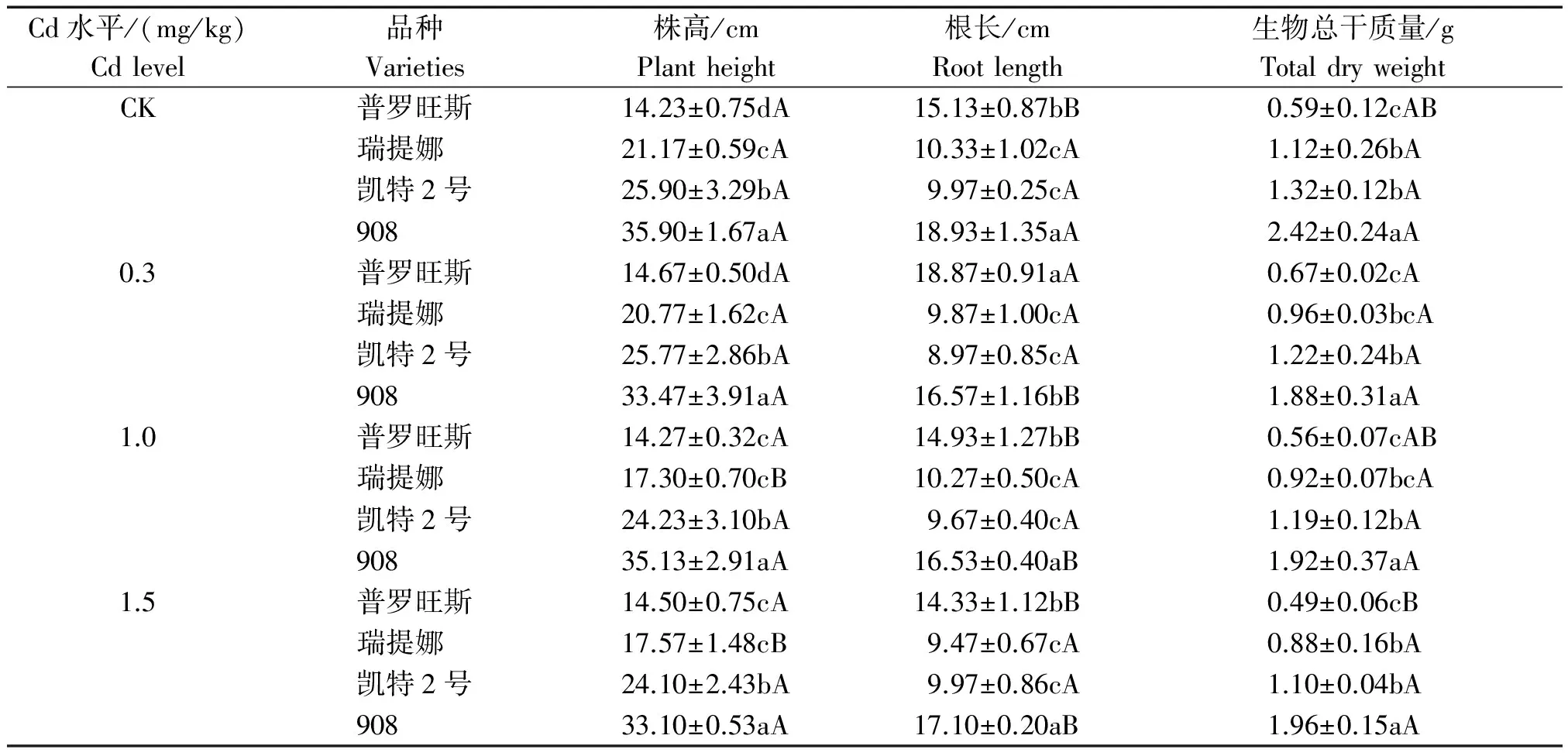
表2 镉胁迫下不同番茄品种的生长指标 Tab.2 Growth indices of different tomato varieties under cadmium stress
注:同列数据后不同小写字母表示同一浓度不同品种间差异显著性(P<0.05);大写字母表示同一品种不同浓度间的差异显著性(P<0.05)。表3-4同。
Note:After the same data,different lowercase letters showed significant difference between the same concentration of different varieties (P<0.05);Uppercase letters showed significant difference between different concentrations of the same variety (P<0.05). The same as Tab. 3-4.
2.3 镉胁迫下不同番茄品种抗性指标的差异特征
丙二醛(MDA)是膜系统脂质过氧化的终产物,其含量可以反映膜脂过氧化程度和植物对逆境抗性的强弱,当细胞遭受逆境胁迫时,细胞膜发生氧化分解,MDA大量积累,其含量越高表明植物抗逆境能力越差[13]。光合色素含量的高低是判定植物光合作用强弱的一个重要生理指标[14],重金属通过干扰呼吸和光合过程中的电子传递、生物酶活性和破坏叶绿体的完整性来影响植物的光合作用。过氧化物酶(POD)广泛分布于植物各器官组织,是活性氧清除系统中的重要酶类,在植物应对逆境环境方面发挥着重要作用,一般而言,POD活性的提高能缓解重金属胁迫。植物在逆境下由于体内活性氧代谢加强而使H2O2发生累积。H2O2可以直接或间接地氧化细胞内核酸蛋白质等生物大分子并使细胞膜遭受损害。过氧化氢酶(CAT)可以清除H2O2是植物体内重要的酶促防御系统之一。因此,植物组织中H2O2含量和CAT活性与植物的抗逆性密切相关。表3反映了在Cd胁迫下不同番茄品种上述4种抗性指标的测定结果。
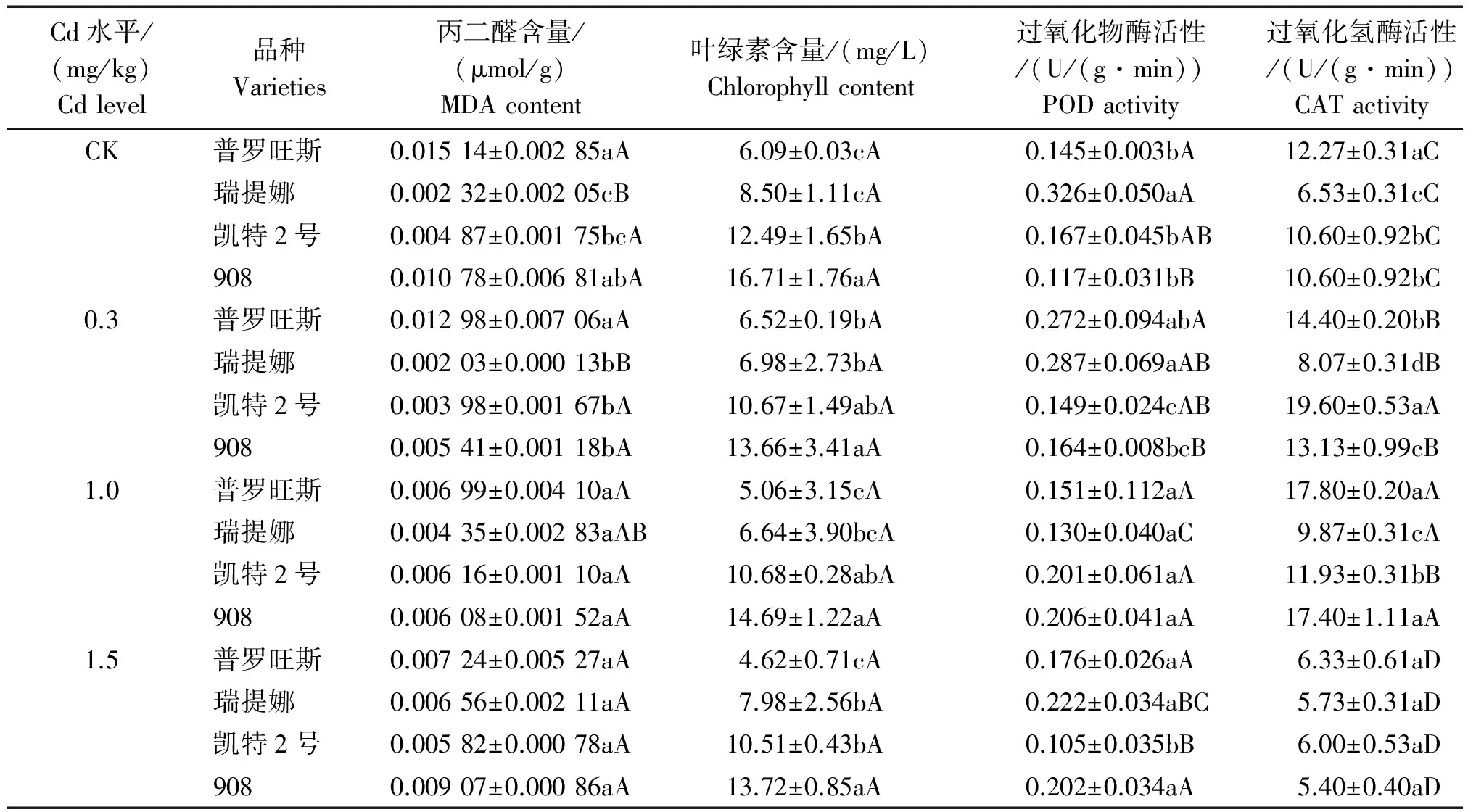
表3 镉胁迫下不同番茄品种的抗性指标 Tab.3 Resistance index of different tomato varieties under cadmium stress
由表3可知,在低浓度Cd(0.3 mg/kg)胁迫下,4项抗性指标含量在不同品种中均有不同表现。4个品种MDA的含量大小表现为普罗旺斯>瑞提娜≈凯特2号≈908;叶绿素含量表现为908≈凯特2号>瑞提娜≈普罗旺斯;POD活性为普罗旺斯≈瑞提娜>凯特2号≈908;CAT活性为凯特2号>普罗旺斯>908>瑞提娜。因不同品种指标属性的自然差异性,仅仅比较Cd胁迫下指标在不同品种间的差异无法反映其抗性大小,因此,为进一步充分说明这一问题,比较了Cd胁迫处理与对照处理的变化,利用显著差异水平以及增减幅度指标判断各个品种的耐Cd抗性。与对照相比,4个品种MDA含量均有不同程度降低,降低12.47%~49.84%;普罗旺斯的叶绿素含量较对照有所增加,增幅为7.06%,其余品种叶绿素含量均有所下降,降低14.56%~18.22%,但各品种均未达显著水平,说明低Cd胁迫对4种番茄幼苗的光合作用尚未产生明显的破坏性影响;凯特2号和瑞提娜中的POD活性比对照有一定的降低,而普罗旺斯和908中的POD活性却呈增加趋势,但均未达显著水平;4个品种中的CAT活性比对照均有显著提高(P<0.05),增幅为17.39%~84.91%。可见,在低Cd含量下4个番茄品种幼苗中的抗氧化酶系统CAT优先启动了抗性功能,以保护膜质免遭破坏的风险。
在中等浓度Cd(1.0 mg/kg)胁迫下,4个品种MDA的含量大小表现差异不明显;叶绿素含量表现为908≈凯特2号>瑞提娜≈普罗旺斯;POD活性差异不显著;CAT活性为普罗旺斯≈908>凯特2号>瑞提娜。与对照相比,4个品种叶绿素含量均有不同程度降低,降低12.06%~21.87%,均未达显著水平;MDA含量在普罗旺斯和908中均有不同程度减少,均未达显著水平,而在瑞提娜和凯特2号中却有不同程度提高,增幅分别为87.50%和26.59%;POD活性在普罗旺斯、凯特2号和908中均有一定提高,分别提高了4.14%,20.60%,76.07%,其中,908的POD活性与对照相比达到了显著水平(P<0.05),而在瑞提娜中的POD活性却显著降低(P<0.05),降幅为60.22%,说明瑞提娜的抗氧化酶系统已经受到严重影响,4个品种CAT活性均有提高,增幅为12.58%~64.15%,均达显著水平,但增幅与低Cd胁迫处理相比有所降低,因此,中等浓度Cd胁迫使CAT活性的抗性表现受到了显著影响。可见,4个品种CAT在中等Cd浓度胁迫下仍继续发挥重要的抵抗作用,而908中的POD在中等Cd胁迫下也启动了抗性功能,其在逆境下表现出了较强的抗性功效。
在高浓度Cd(1.5 mg/kg)胁迫下,4个品种MDA的含量大小差异不明显;叶绿素含量表现为908>凯特2号≈瑞提娜>普罗旺斯;POD活性为908≈普罗旺斯≈瑞提娜>凯特2号;CAT活性在4个品种间表现差异不明显。与对照相比,4个品种叶绿素含量均有所降低,降低6.22%~24.15%,但均未达显著水平;MDA含量在普罗旺斯和908中均有所降低,降低幅度分别为52.18%和15.88%,但均未达显著水平,而在瑞提娜和凯特2号中有一定增加,增幅分别为183.02%和19.55 %,且瑞提娜MDA含量与对照相比达到了显著水平(P<0.05),比中等Cd胁迫处理有大幅提高,并与瑞提娜生长指标受到显著影响相符合。说明了高Cd含量胁迫使瑞提娜番茄幼苗体内活性氧的积累继续增强,膜脂过氧化加剧恶化,细胞膜完整性遭到严重破坏;POD活性在凯特2号和瑞提娜中明显降低,降低幅度分别为37.20%和31.90%,瑞提娜达到了显著水平(P<0.05),而在908和普罗旺斯中却有提高,增幅分别为72.65%和21.61%,其中,908达到了显著水平(P<0.05),与中等Cd胁迫的增幅相持平,说明了908在高Cd胁迫下仍表现出了较为强劲的抗性优势;4个品种CAT活性在高Cd含量下均显著下降(P<0.05),受到了显著影响,降低12.24%~49.06%。可见,在高Cd浓度胁迫下,908的POD继续发挥重要的抵抗作用,仍然表现较强的抗性,而瑞提娜酶系统则受到了严重影响,抗性表现最弱。
比较不同Cd浓度胁迫影响的变化发现,不同番茄品种的指标对Cd浓度胁迫的响应出现了明显的品种差异特征。普罗旺斯、凯特2号、908 3个品种的MDA含量随Cd浓度增加的响应变化差异不大;瑞提娜在低Cd胁迫时的响应没有显著变化,但随Cd浓度提高MDA含量显著增加,从中等Cd胁迫时的增幅87.50%提升到了高Cd胁迫时的增幅183.02%。4个番茄品种幼苗中的CAT随Cd浓度增加呈现先升高后下降的趋势。在低Cd胁迫时CAT优先启动了抗性响应功能,增幅为17.39%~84.91%;在中等Cd胁迫时CAT继续发挥抗性功能,增幅为12.58%~64.15%,但功效有所下降;在高Cd胁迫时4个品种CAT均出现下降趋势,抗性严重下降,降幅为12.24%~49.06%。4个番茄品种幼苗中的POD在低Cd胁迫时没有明显响应变化;在中等Cd胁迫时908中的POD大幅增加,启动了抗性功能;瑞提娜活性下降,抗性严重下降,降幅60.22%;其余品种没有明显响应变化;在高Cd胁迫时908的POD活性继续增强,增幅72.65%,瑞提娜POD活性继续下降,降幅31.90%,其余品种仍没有变化。说明不同品种启动抗氧化酶系统的Cd胁迫浓度存在明显差异。另外发现,在同等胁迫条件下,908植株吸收到体内的高Cd浓度诱导了抗氧化酶活性的高增加幅度,反映了908植株对活性氧清除能力强于其余3个品种。
2.4 镉胁迫下不同番茄品种地上部、根部镉含量的差异特征
由图1,2可知,在不添加Cd对照处理下,4个品种地上部和根部Cd含量并无显著差异性。然而,在所有Cd浓度胁迫下不同品种间呈现出了较大的Cd吸收差异特性。从地上部Cd含量看,908地上部Cd含量均显著高于其他品种(P<0.05),分别为普罗旺斯、瑞提娜和凯特2号的6~12倍、6~7倍、5~8倍,普罗旺斯Cd含量最低,但后3种差异并不显著。可见,在中、高等Cd胁迫下普罗旺斯地上部吸收Cd的能力表现最弱,908表现最强。
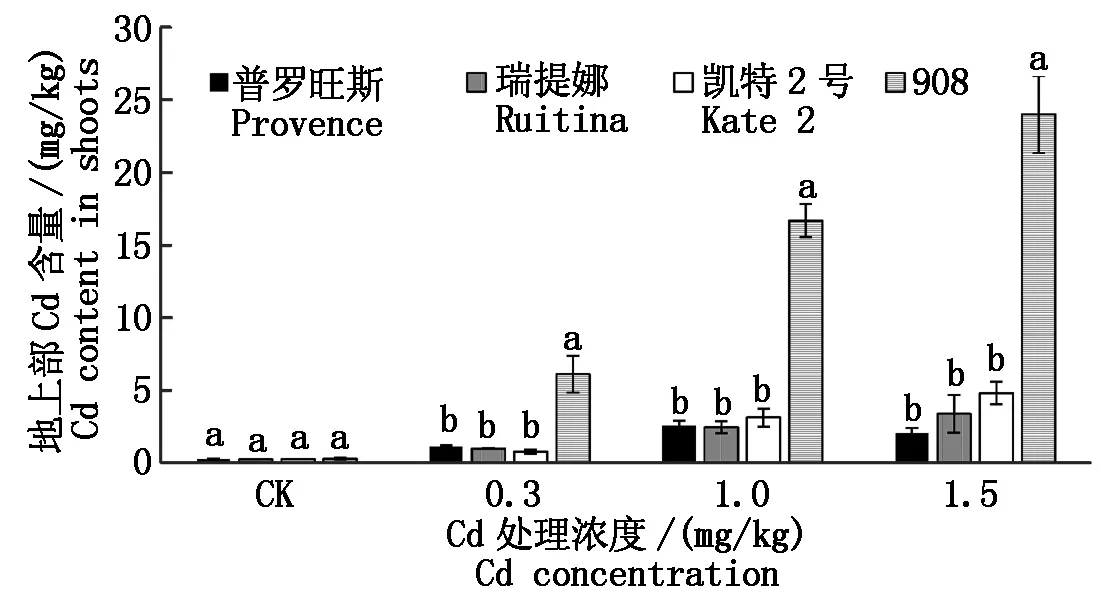
图柱上不同字母表示处理间在0.05水平上差异显著。图2同。Values followed by different letters are significantly different at the 0.05 probability level. The same as Fig.2.
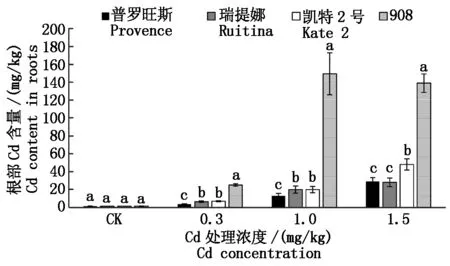
图2 不同处理浓度下4种番茄根部 Cd含量的差异性Fig.2 Difference of Cd content in four tomato roots under different treatment concentrations
从根部Cd含量看,在所有Cd浓度胁迫下,908根部Cd含量均显著高于其他品种,分别为普罗旺斯、瑞提娜和凯特2号的5~12倍、4~8倍、3~8倍,与地上部Cd含量表现出一致的规律性。在Cd浓度为0.3,1.0 mg/kg时,根部Cd含量在凯特2号和瑞提娜之间差异不明显,但两者均显著高于普罗旺斯;在Cd浓度为1.5 mg/kg时,凯特2号根部Cd含量显著高于普罗旺斯和瑞提娜,且后两者差异不显著。可见,在中、高等Cd胁迫下普罗旺斯根部吸收Cd的能力最弱,908最强。
从番茄品种对Cd积累的角度看,前人研究成果认为,理想的Cd低积累植物应同时具备以下4个特征[15]:该植物的地上部和根部的重金属含量都很低或者可食部位低于有关标准;该植物对重金属的累积量小于土壤中的该重金属的浓度,即富集系数(EF)<1;该植物从根部重金属向地上部转运能力较差,即转运系数<1;该植物对重金属毒害具有较高的耐受性,在较高的重金属污染下能够正常生长且生物量无显著下降。基于上述4点特征,综合本研究的Cd积累和生长指标特征,在中等Cd胁迫下除了瑞提娜受到影响比较大外,其余3个品种的生长指标基本没有受到影响,且转运系数均<1(表4)。因此,中、高等Cd胁迫下凯特2号、普罗旺斯和908基本满足低Cd积累植物特征,但908植株的地上部和根部Cd含量最高,普罗旺斯表现最低,可以选择普罗旺斯为Cd低积累番茄品种。由于本试验是在添加Cd培养液中的番茄生长幼苗进行取样分析的,关于不同基因型番茄在土壤环境下生育后期植株对Cd的胁迫响应以及果实Cd的累积量差异有待于进一步研究和证实。
从番茄品种对Cd的抗性角度看:一般高Cd抗性植物应同时具备以下5个特征:植株的生长发育受抑制较小;叶绿体受破坏较小,光合作用受到影响较小;植株生物量变化较小,干质量减轻较少;体内丙二醛增幅较小、游离脯氨酸积累增多;体内SOD、POD、CAT酶活性增加。在4个番茄品种中,908在中、高等Cd胁迫下符合5个特征的基本表现,属于高Cd抗性番茄品种。
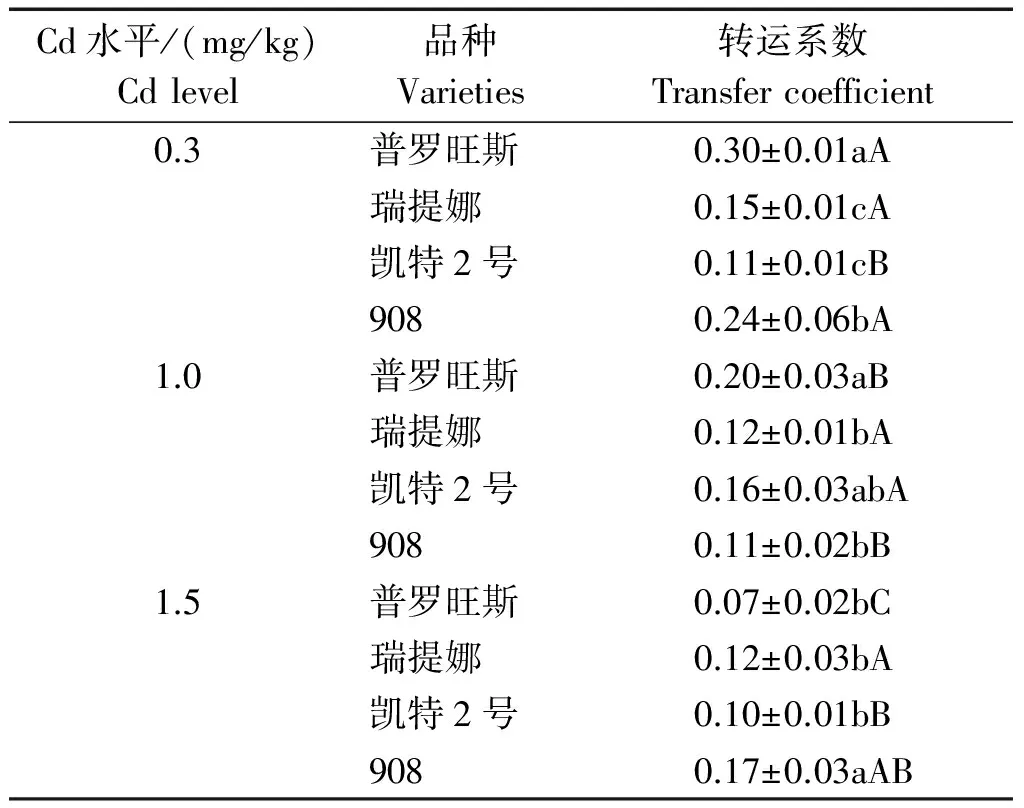
表4 镉胁迫下不同番茄品种的转运系数 Tab.4 Transfer coefficients of different tomato varieties under cadmium stress
3 讨论
Cd为植物非必需营养元素,在植物体内过量积累会对植物造成毒害,通常表现为植物生长迟缓、根系生长受阻、养分代谢失调及其他的生理紊乱,严重时甚至导致植物死亡[16]。有研究表明[17-18],一般植物体内Cd含量达到0.2~0.8 mg/kg就对植物产生毒性效应,导致植物枯萎和生长受到抑制。本研究结果表明,4个番茄品种添加Cd培养10 d后,低Cd浓度对普罗旺斯生长有一定的促进作用,尤其根系反应更加敏感,而对其余品种的影响不明显;中、高Cd浓度显著抑制了瑞提娜的株高生长,对其余品种的影响不明显。这一结果表现与佘玮等[19]得出的同一植物不同品种对Cd的积累性和耐性有很大差异的结论相一致。
本研究证实,在Cd胁迫处理下,908地上部和根部Cd含量均显著高于其他品种,地上部为其他品种的5~12倍,根部为其他品种的3~12倍。不同作物种类及同种作物不同品种间Cd积累量存在显著差异已经得到研究者的认可[20-22],然而,导致Cd积累量差异的机制目前还不十分明确。Becher、Weber等[23-24]研究证实,重金属超积累拟南芥halleri根茎中AhIRT3、AhZIP3、AhZIP6和AhZIP12,及根部AhZIP9的高表达可能是Cd大量吸收的主要原因。赵首萍等[25]也推测,高Cd积累番茄品种根系对离子态Cd较高的吸收速率及Nramp2、Nramp3和ZIP的高表达可能是获得高Cd积累量的原因之一。另外,本研究认为,CK处理下908的株高和生物总干质量均表现出了强劲的生长优势,2项指标均远高于其他品种,这可能说明了番茄植株生物量对Cd积累的影响也是一个重要因素,其植株蒸腾拉力对高Cd积累不容忽视,但目前关于普罗旺斯、瑞提娜、凯特2号和908 4个品种表现的差异机理尚不能给出充分、深入的解释,有待于进一步深入研究。
抗氧化酶系统是番茄植株中重要的保护系统之一。POD和CAT是其中重要的抗氧化酶类,其活性的提高能够清除Cd2+诱导产生的活性氧,降低其对番茄幼苗的毒害效应,对番茄幼苗具有一定的保护作用。但是,这种保护作用是有一定限度的。本研究已证实,4个番茄品种幼苗中的CAT在低、中等Cd胁迫时启动了抗性保护功能,而在高Cd胁迫时4个品种CAT抗性系统均受到了严重影响,活性显著下降;908中的POD抗性系统在中、高等Cd胁迫时启动了抗性保护功能,而瑞提娜却受到了严重影响,活性显著下降,其余品种没有出现显著响应信号。同时,与其余品种相比,品种908根、茎部Cd含量高,再加上抗氧化酶POD和CAT在胁迫下活性增幅最强,可以推测,908根、茎部在高Cd胁迫下具有较强的活性氧清除能力,从而导致MDA的产生有所下降,在一定程度上避免了对细胞质膜的损伤。
在不添加Cd营养液的培养条件下,番茄幼苗生长10 d后的生长指标及体内抗性指标特征表现出了较大的自然差异性。品种908在株高、根长、总生物量生长指标以及抗性指标MDA和叶绿素含量上均表现出最高的自然特征;普罗旺斯在CAT活性上表现最为突出;瑞提娜在POD活性上表现最高。
在不同Cd含量胁迫条件下,4个品种对生长指标的响应有不同之处。普罗旺斯根系对低Cd含量胁迫的响应最为敏感,受到了明显的刺激作用,比对照增长24.72%;瑞提娜株高对中、高Cd含量胁迫的响应最为敏感,受到了明显的抑制作用。
不同番茄品种的抗性指标对Cd浓度胁迫的响应出现了明显的品种差异特征。与对照相比,瑞提娜幼苗中的MDA含量在高浓度Cd胁迫时显著升高,质膜受到了显著影响;4个番茄品种幼苗中的CAT在低、中等Cd胁迫时启动了抗性响应功能,酶活性明显增强,在高Cd胁迫时酶活性大幅下降,抗性功能严重削弱;908的POD在中、高等Cd胁迫时启动了抗性功能,酶活性大幅增强,瑞提娜的POD活性大幅下降,抗性严重遭受影响。
在所有Cd浓度胁迫下不同品种间呈现出了较大的Cd吸收差异特性。4个番茄品种中,908地上部和根部的Cd含量均显著高于其他品种,为其他品种的3~12倍,普罗旺斯含量最低。综合推断普罗旺斯属于Cd低积累品种;908属于高Cd抗性品种。
参考文献:
[1] 刘明明. 邻苯二甲酸二丁酯和镉复合作用对油菜生理生化指标和土壤酶活性的影响研究[D]. 泰安:山东农业大学,2014.
[2] 姚会敏,杜婷婷,苏德纯. 不同品种芸薹属蔬菜吸收累积镉的差异[J]. 中国农学通报,2006,22(1):291-294.
[3] 陈 瑛,李廷强,杨肖娥,等. 不同品种小白菜对镉的吸收积累差异[J]. 应用生态学报,2009,20(3):736-740.
[4] 李 婧,周艳文,陈 森,等. 我国土壤镉污染现状、危害及其治理方法综述[J]. 安徽农学通报,2015,21(24):104-107.
[5] 庞荣丽,王瑞萍,谢汉忠,等. 农业土壤中镉污染现状及污染途径分析[J]. 天津农业科学,2016,22(12):87-91.
[6] 赵本行,陈康姜,何楚斌,等. 大豆作物对污染土壤中重金属镉的富集研究[J]. 天津农业科学,2013,19(11):15-17.
[7] 徐照丽,段玉琪,杨宇虹,等. 不同土类中外源镉对烤烟生长及土壤生物指标的影响[J]. 华北农学报,2014,29(S):176-182.
[8] 黄运湘,廖柏寒,王志坤,等. 不同大豆品种镉毒害效应及耐镉差异性[J]. 湖南农业大学学报:自然科学版,2008,34(5):519-524.
[9] 王蕴赟,李 波,施丽莉,等. 浙江省土壤镉污染现状及修复技术[J]. 浙江化工,2016,47(7):42-44.
[10] 朱德强. 土壤镉污染及其修复方法[J]. 现代农业科技,2017(11):175-177.
[11] 苗亚琼,林 清. 广西土壤重金属镉污染及对人体健康的危害[J]. 环境与可持续发展,2016,41(5):171-173.
[12] 范洪黎,周 卫. 镉超富集苋菜品种(AmaranthusmangostanusL.)的筛选[J]. 中国农业科学,2009,42(4):1316-1324.
[13] 王 梅,徐正茹,张建旗,等. 遮阴对10种野生观赏植物生长及生理特性的影响[J]. 草业科学,2017,34(5):1008-1016.
[14] 胡国涛,于 阳,杨 兴,等. 速生树种竹柳对镉的吸收,积累与分布特性[J]. 环境科学学报,2016,36(4):1508-1514.
[15] 廖芳芳,付文婷,王永平,等. 镉低积累农作物筛选研究进展[J]. 南方农业,2015,9(25):58-60.
[16] 何梨香,黄运湘,黄楚瑜,等. 圆叶决明对镉胁迫的生理响应[J]. 草业学报,2016,25(2):198-204.
[17] Singh B R,Narwal R P,Jeng A S,et al. Crop uptake and extractability of cadmium in soils naturally high in metals at different pH levels[J]. Communications in Soil Science and Plant Analysis,1995,26(13/14):2123-2142.
[18] Ye Z H,Baker A,Wong M H,et al. Zinc,lead and cadmium tolerance uptake and accumulation by the common Ree.phragmitesaustralis(Cav.)Trin.ex steudel[J]. Annals of Botany,1997,80(3):363-370.
[19] 佘 玮,揭雨成,邢虎成,等. 苎麻耐镉品种差异及其筛选指标分析[J]. 作物学报,2011,37(2):348-354.
[20] 林忠全,李佳繁,康友宏,等. 土壤外源镉污染对番茄幼苗生长影响研究[J]. 北方园艺,2009(5):81-83.
[21] 宋 建,金凤媚,薛 俊,等. 不同番茄品系吸收和积累镉能力的差异[J]. 安徽农业科学,2016,44(5):33-35.
[22] 刘维涛,周启星,孙约兵,等. 大白菜(BrassicapekinensisL.)对镉富集基因型差异的研究[J]. 应用基础与工程科学学报,2010,18(2):226-235.
[23] Becher M,Talke I N,Krall L,et al. Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyper accumulatorArabidopsishalleri[J].Plant Journal,2004,37(2):251-268.
[24] Weber M,Harada E,Vess C,et al. Comparative microarray analysis ofArabidopsisthalianaandArabidopsishalleriroots identifies nicotianamine synthase,a ZIP transporter and other genes as potential metal hyper accumulation factors[J].Plant Journal,2004,37(2):269-281.
[25] 赵首萍,张永志,张 棋,等. 两种基因型番茄对镉胁迫响应差异[J]. 植物营养与肥料学报,2015,21(5):1261-1268.
猜你喜欢
纺织科技进展(2021年5期)2021-07-22 08:41:32
纺织科技进展(2021年4期)2021-07-22 06:47:00
纺织科技进展(2021年3期)2021-06-09 08:07:12
空间科学学报(2021年1期)2021-05-22 01:36:34
读友·少年文学(清雅版)(2020年5期)2020-09-09 09:40:50
经济技术协作信息(2018年7期)2019-01-14 03:05:44
环境保护与循环经济(2017年5期)2018-01-22 02:56:44
天津音乐学院学报(2017年3期)2017-10-09 04:58:47
海外星云(2016年17期)2016-12-01 04:18:43
中国果菜(2016年9期)2016-03-01 01:28:44
