
万峰山森林群落演替中土壤肥力和微生物数量特征分析
2018-04-27 11:35:53张敏倩袁剑飞钟在云华朝晖冯显帮邹哲宇朱陈浩郭联华
浙江林业科技 2018年1期
张敏倩,袁剑飞,钟在云,谭 斌,华朝晖,冯显帮,邹哲宇,朱陈浩,郭联华
(1. 浙江农林大学 林业与生物技术学院,浙江 杭州 311300;2. 江西省遂川县林业局,江西 遂川 343900;3. 四川省邻水县万峰山林场,四川 邻水 638500;4. 浙江省遂昌县林业局,浙江 遂昌 323300)
土壤是植物群落的重要环境因子之一,它在对植物群落发生作用的同时,自身发育也受到植物群落的影响[1]。研究表明,土壤的发育与植物群落演替是两个密不可分、相辅相成的过程[2],但以往研究多强调不同植被现状对土壤的影响,而较少注意植被演替与土壤发育之间的关系[3]。
土壤微生物作为土壤生态系统重要组成部分,在土壤生态系统的能量流动和养分转化中起着重要的作用[4],不仅影响植被的生长,也受到植被影响,两者相互作用,相互影响,因此,微生物成为土壤质量变化的最敏感指标之一,在土壤质量评价中越来越受到重视[5]。
地上枯落物是指由植物地上部分产生并归还到地表的所有有机物质的总称,其质量不仅决定输入土壤的养分组成和数量,也能够通过自身的性质影响土壤微生物的数量和活性,进而影响土壤中有机物质的分解[6-7]。
天然更新的常绿阔叶林有利于生物多样性的增加与土壤养分的保蓄[8]。润楠Machilus pingii为樟科Lauraceae润楠属Machilus常绿大乔木,国家Ⅱ级重点保护野生植物,我国西南特有种,主要分布于四川盆地西缘山地[9],其作为珍贵的用材树种,具有很高的经济价值和收藏价值。四川邻水县万峰山风景区属亚热带常绿阔叶林,是全国最大的润楠集中分布地之一,然而,目前随着当地的旅游资源开发,润楠资源破坏相当严重,润楠野生资源处于濒危状态,种群数量在15 000株以下[9],同时,市场对润楠木材的需求不断增加,导致润楠木材供求矛盾加剧,因此,润楠林生长状况受到越来越广泛的关注。目前关于润楠生理特性和幼苗生长等方面的研究较多[10-11],而尚未见到在群落演替过程中其对土壤影响的研究报道。
关于森林演替模式,国内早有大量研究,认为常绿阔叶林的演替规律一般是按照草本植物群落-灌草丛-灌丛-针叶林-针阔叶混交林-常绿阔叶林6个演替阶段进行的[12],或者是其中的部分演替阶段[13],三峡库区、缙云山等北亚热带和中亚热带森林类型也基本遵循该规律[14-15]。本实验采用空间代替时间的方法,以四川邻水县万峰山林场中针叶林-针阔混交林-润楠林为演替序列过程为对象,分析在植被恢复过程中不同植被类型土壤微生物数量的差异,以及土壤理化性质与土壤微生物数量特征间的耦合关系,以期进一步探讨植被类型-土壤的相互作用过程,为森林群落科学经营和润楠资源保护提供建议和依据。
1 研究方法
1.1 研究区自然概况
研究地点位于四川铜锣山脉万峰山,107°05′46″ ~ 107°06′22″ E,30°24′07″ ~ 30°24′32″ N,属国家天然林保护和退耕还林(草)工程建设区。万峰山地处四川盆地东部,华蓥山东麓,位于邻水县龙安镇境内,海拔800 ~1 054 m,地势高峻,植物种类繁多,至今仍有保护完好的亚热带原始林33万余hm2。该区属于亚热带季风气候区,气候温暖湿润,年平均气温17℃,年平均日照1 230.7 h,68%降水量集中在5,10月。森林植被类型为亚热带常绿阔叶林,地带性土壤类型为黄壤,地带性植被以樟科、松科、杉科等为主,植被多为次生阔叶林和针叶林,多呈镶嵌状分布,次生阔叶林主要有润楠,喜树Camptotheca acuminata,酸枣Ziziphus jujubavar.spinosa,青冈Cyclobalanopsis glauca等组成。人工林主要由马尾松Pinus massoniana,杉木Cunninghamia lanceolata,油茶Camellia oleifera等组成。
1.2 实验设计和野外样品采集
2015年7月,在野外详细踏查的基础上,在针阔混交林、润楠林及针叶林3个不同植被覆盖类型区中各选择4个海拔、坡向和坡度相近,面积为20 m×20 m的样地,在各样地中按照“S”型选取5个代表性的样点,用土钻于表层(0 ~ 20 cm)采集土壤样品,5个样点的土壤混匀为1个土样。采集的新鲜土壤在实验室内一分为二,一份过2 mm土壤筛后,直接用于微生物种类及数量的测定;另一份自然风干后用于土壤基本理化性质的测定。对枯落物的采集同样按照“S”型选取5 个代表性的样点,拾取每个样点1 m×1 m面积的枯落物若干,及时装入无菌保鲜袋带回实验室,置于75℃烘箱中烘8 h,用于相关指标的测定。不同植被类型区基本情况见表1。

表1 样地概况Table 1 Information of the sample plots
1.3 样品分析
1.3.1 土壤及枯落物理化性质测定 枯落物样品经H2SO4-H2O2消煮后,蒸馏法测定全氮,钼锑抗比色法测全磷,火焰光度法测全钾;土壤速效钾采用火焰光度法;pH值用酸度计法;有机碳用硫酸重铬酸钾外加热法;碱解氮采用碱解扩散法;土壤孔隙度用环刀法测定[16]。
1.3.2 土壤微生物特征分析方法 采用稀释平板计数法进行土壤细菌、放线菌、真菌数量的测定[17],培养基分别为牛肉膏蛋白胨培养基、改良高氏一号(苯酚 500 mg·L-1)培养基、马丁(Martin)孟加拉红-链霉素(链霉素30 mg·L-1)培养基。按照配方配制培养基后,先在121℃下灭菌15 min,然后冷却至45 ~ 50℃备用。凝固后的培养基可加热溶解后使用。细菌在37℃,放线菌在30℃下培养10 d,真菌在25℃下培养5 d。
结果计算方法:每克样品中的菌落数=同一稀释度6次重复的菌落平均数×稀释倍数。
1.4 数据处理
用SPSS 13.0软件对土壤微生物数量及土壤理化性质进行单因素方差分析和相关性分析。显著性水平设定为P=0.05或P=0.01。
2 结果与分析
2.1 不同森林植被类型枯落物营养元素和土壤理化性质
3种植被类型枯落物营养元素含量大小顺序均为全氮>全钾>全磷(表2)。不同植被类型枯落物中3种营养元素的含量不同,其中全氮、全磷含量以润楠林最高,分别为12.241,0.216 g·kg-1;针阔混交林枯落物全钾含量(1.387 g·kg-1)显著高于其他2种植被(P<0.05)。
从表3可知,3种植被类型土壤呈强酸性,其中润楠林土壤pH值为4.4,显著高于在针阔混交林和针叶林(P<0.05)。土壤总孔隙度介于37.618% ~ 45.425%,以针阔混交林土壤最高,但不同植被类型间没有显著差异(P>0.05)。针阔混交林土壤有机碳和碱解氮的质量分数均显著高于润楠林和针叶林(P<0.05),分别达27.000 mg·kg-1,248.500 mg·kg-1。不同植被类型土壤速效钾质量分数介于114.0 ~ 128.0 mg·kg-1,无显著性差异(P>0.05)。根据土壤养分分级标准[18],3种植被类型土壤质量均较高。
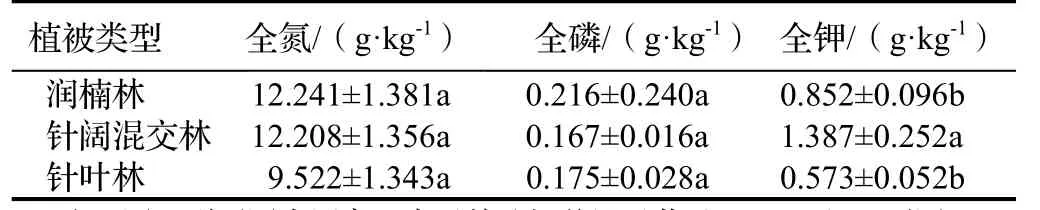
表2 不同植被类型枯落物营养元素含量Table 2 Nutrient content in forest litter under different vegetation types

表3 不同林型土壤理化性质Table 3 Soil physical and chemical properties under different forest types
2.2 不同植被类型土壤微生物数量
不同植被类型表层(0 ~ 20 cm)土壤微生物总量大小依次为针阔混交林>针叶林>润楠林(表4),这3种植被类型下的土壤微生物总量存在显著差异(P<0.05),从不同类群微生物数量来看,放线菌和真菌数量均在润楠林中最大,其中放线菌比其他2种类型林地中的数量高,细菌数量在针阔混交林中最大;统计不同微生物类群的百分比发现,3大类微生物在微生物区系组成中细菌占主导地位,其数量在不同植被类型土壤下占微生物总量的96.37% ~ 99.70%,其次是放线菌,含量最少的是真菌,仅占微生物总数的0.12% ~ 0.32%。
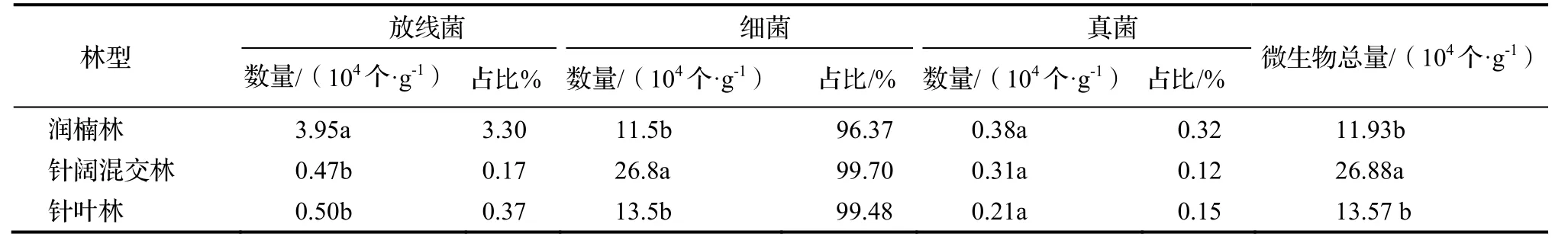
表4 3种不同林型土壤的三种微生物数量Table 4 Microbial populations at 0-20 cm soil under different forest types
2.3 土壤微生物数量特征与土壤理化性质部分因子间的相关性
不同微生物种类和数量与土壤理化性质部分因子间存在一定的相关性(表 5),润楠林土壤细菌、真菌数量与土壤 pH值有显著或极显著负相关性,放线菌、细菌数量与总孔隙度的相关性达极显著水平,真菌数量与速效钾呈极显著正相关;针阔混交林中真菌数量与土壤 pH值呈显著负相关,与有机碳、碱解氮、速效钾均呈极显著正相关,细菌数量与速效钾、总孔隙度的相关性达极显著水平,放线菌数量与总孔隙度也呈极显著正相关;针叶林中放线菌数量与土壤 pH值、有机碳、碱解氮、速效钾、总孔隙呈显著或极显著正相关,细菌数量与土壤pH值有显著负相关性,真菌数量与总孔隙度的相关性也达显著水平。

表5 不同林型下土壤微生物数量与土壤理化性质部分因子间的相关性分析Table 5 Correlation analysis on soil microbial populations with soil physical and chemical properties under different forest types
3 结论与讨论
林地土壤理化性质是土壤质量的重要反映指标,反映了森林对土壤的影响程度,是衡量土壤肥力高低的主要指标,既受植被枯落物类型的影响,也影响土壤微生物数量[19-20]。从研究结果看,在同一地域条件下,由于其地上植物群落不同,其土壤理化性质也不同。针叶林、针阔混交林和润楠林3种植被类型的土壤理化性质总体表现为:土壤总孔隙度、土壤微生物数量、养分含量以针阔混交林最高,润楠林比针叶林稍高。
地上枯落物是指由植物地上部分产生并归还到地表的所有有机物质的总称,其质量不仅决定输入土壤的养分组成和数量,也能够通过自身的性质影响土壤微生物的数量和活性,进而影响土壤中有机物质的分解[6-7]。植被类型不同从而地上枯落物不同,对土壤理化性质和微生物数量有显著影响。宋会兴等研究表明,随着演替进展,土壤中有机质进一步积累,土壤厚度增加,持水、保肥能力增强,针叶树种逐渐被常绿阔叶树种取代[2],枯落物也从质地较硬的针叶变为易于被分解的阔叶,加上随时间推移,土壤积累的枯枝落叶营养元素含量增加,土壤有机质质量分数提高,结构疏松,能够为微生物活动提供良好的营养和通气条件,有利于微生物的生长繁殖,促进对枯落物的分解,进而增加土壤养分质量分数。本研究表明,枯落物营养元素含量、土壤微生物数量和养分质量状况依次为针阔混交林>润楠林>针叶林,其中针阔混交林的微生物总量和养分质量分数普遍显著高于其他2种植被类型,这不符合从针叶林进展演替至阔叶林土壤状况的变化规律,这可能与该地区的润楠阔叶林并不是由针阔混交林演替而来,而是因人为择伐其它树种而由针阔混交林直接转变成润楠阔叶林有关,针阔混交林中树种多样性丰富[21],其土壤养分的循环、转化和积累状况良好,更有利于微生物生长繁殖,土壤肥力水平更高[22]。
微生物种类与数量影响土壤的生物化学活性及土壤养分的组成与转化,是维持和恢复林地生产力的主要因子之一[23],既反映了土壤各因素对微生物的影响,也反映了微生物对植物生长发育、土壤理化性质的影响和作用[24]。根据微生物数量与土壤理化性质的相关性分析发现,土壤总孔隙度与三大微生物数量关系密切,细菌和放线菌数量在润楠林和针阔混交林中与总孔隙度均达极显著相关,说明这2种微生物在改善土壤结构方面起着一定作用。但在针叶林中,放线菌、真菌两种数量占比小的微生物与总孔隙度相关性显著,而数量最多的细菌与总孔隙度相关性不显著,陈婷等认为微生物对枯落物的分解作用受枯落物本身特性的影响[25],造成本研究这一结果的原因可能是针叶林枯落物木质素含量较多,抑制了数量最多的细菌对枯落物的分解作用,从而间接影响其对土壤总孔隙度的作用[26]。润楠林中放线菌和真菌数量最多,放线菌的数量比其它2种林地高一个数量级,相差近8倍以上,这可能是由于润楠林的pH值较于其他两种林地更高,放线菌更适于在中性或微碱性土壤环境中生存[27]。
本研究所选的针叶林-针阔混交林-润楠林的群落演替序列中土壤动态变化初步反映了四川万峰山林地不同演替阶段土壤状况的变化。研究结果显示,针阔混交林树种多样性丰富,土壤养分状况最好,土壤结构疏松,更有利于微生物生长繁殖;润楠林的土壤状况比针阔混交林稍差,但总体上超过针叶林,适合放线菌的生长。
参考文献:
[1] 焦菊英,马祥华,白文娟,等. 黄土丘陵沟壑区退耕地植物群落与土壤环境因子的对应分析[J]. 土壤学报,2005,42(5):744-752.
[2] 宋会兴,苏智先,彭远英. 渝东山地黄壤肥力变化与植物群落演替的关系[J]. 应用生态学报,2005,16(2):223-226.
[3] 常超,谢宗强,熊高明,等. 三峡库区不同植被类型土壤养分特征[J]. 生态学报,2009,29(11):5978-5985.
[4] 李静鹏,徐明锋,苏志尧,等. 不同植被恢复类型的土壤肥力质量评价[J]. 生态学报,2014,34(9):2297-2307.
[5] 刘占锋,傅伯杰,刘国华,等. 土壤质量与土壤质量指标及其评价[J]. 生态学报,2006,26(3):901-913.
[6] ZAK D R,HOMES W E,WHITE D C,et al.Plant Diversity Soil Microbial Communities, And Ecosystem Function: Are There Any Links?[J].Ecology,2003,84(8):2042-2050.
[7] WARDLE D A,YEATES G W,BARKER G M,et al.The influence of plant litter diversity on decomposer abundance and diversity[J]. Soil Biol Biochem,2006,38(5):1052-1062.
[8] 胡婵娟,郭雷,刘国华. 黄土丘陵沟壑区不同植被恢复格局下土壤微生物群落结构[J]. 生态学报,2014,34(11):2986-2995.
[9] 丁鑫,肖建华,黄建峰,等. 珍贵木材树种楠木的野生资源调查[J]. 植物分类与资源学报,2015,37(5):629-639.
[10] 程凡. 宜昌润楠化学成分和生物活性的研究[D]. 北京:北京林业大学,2013.
[11] 温美君. 不同强度遮荫环境下刨花润楠幼苗生长及生理特性研究[D]. 南宁:广西大学,2014.
[12] 于明坚. 青冈常绿阔叶林群落动态研究[J]. 林业科学,1999,35(6):42-51.
[13] 韩天丰,周国逸,李跃林,等. 中国南亚热带森林不同演替阶段土壤呼吸的分离量化[J]. 植物生态学报,2011,35(9):946-954.
[14] 程瑞梅,肖文发,王晓荣,等. 三峡库区植被不同演替阶段的土壤养分特征[J]. 林业科学,2010,46(9):1-6.
[15] 雷泞菲,苏智先,宋会兴,等. 缙云山常绿阔叶林不同演替阶段植物生活型谱比较研究[J]. 应用生态学报,2002,13(3):267-270.
[16] 袁久坤,周英. 利用取土钻改进环刀法准确测定土壤容重和孔隙度[J]. 中国园艺文摘,2014(3):25-26.
[17] 姚槐应,黄昌勇. 土壤微生物生态学及其实验技术[M]. 北京:科学出版社,2006:130-140.
[18] 鲁如坤. 土壤农业化学分析方法[M]. 北京:中国农业科技出版社,2000:146-195.
[19] 王希华,黄建军,闫恩荣. 天童国家森林公园常见植物凋落叶分解的研究[J]. 植物生态学报,2004,28(4):457-467.
[20] 池振明. 现代微生物生态学[M]. 北京:科学出版社,2005:8-9.
[21] 黄宇. 杉木纯林、杉-阔混交林与阔叶纯林生态系统土壤质量研究[D]. 北京:中国科学院生态环境研究中心,2004.
[22] 于洋,王海燕,丁国栋,等. 华北落叶松人工林土壤微生物数量特征及其与土壤性质的关系[J]. 东北林业大学学报,2011,39(3):76-80.
[23] 赵萌,方晰,田大伦. 第2代杉木人工林地土壤微生物数量与土壤因子的关系[J]. 林业科学,2007,43(6):7-12.
[24] 陈声明. 微生物生态学导论[M]. 北京:高等教育出版社,2007:101-102.
[25] 陈婷,郗敏,孔范龙,等. 枯落物分解及其影响因素[J]. 生态学杂志,2016,35(7):1927-1935.
[26] 马琳. 荒漠草原典型植物群落枯落物分解对土壤性质的影响[D]. 银川:宁夏大学,2012.
[27] 姜怡,李文均,徐平,等. 盐碱环境放线菌多样性研究[J]. 微生物学报,2006,46(2):191-195.
猜你喜欢
国际呼吸杂志(2019年22期)2019-12-09 09:20:36
江苏农业科学(2019年21期)2019-01-03 02:05:58
现代农业研究(2018年4期)2018-08-11 10:25:02
中国绿色画报(2018年5期)2018-08-07 07:12:26
森林工程(2018年4期)2018-08-04 03:23:16
山东化工(2017年5期)2017-09-16 02:43:15
——以四川盆地龙马溪组页岩气储层为例
石油与天然气地质(2017年3期)2017-07-10 10:28:01
生态学报(2016年7期)2016-04-11 03:31:01
应用海洋学学报(2014年4期)2014-11-22 07:43:54
天然产物研究与开发(2014年6期)2014-04-27 14:15:56
