
富营养化水体对水芹抗氧化系统和营养品质的影响
2018-04-26 02:02雷代东刘明鹏贾永霞
西北植物学报 2018年3期
张 帅,雷代东,王 晨,熊 瑞,刘明鹏,贾永霞
(四川农业大学 资源学院,四川温江 611130)
水体富营养化现象在湖泊、河流、水库等地表水中日趋严重,已成为当今世界水环境的一个突出性难题[1-3]。利用植物修复技术来净化富营养化水体是目前行之有效的生态治理措施[4-5]。大量研究表明,水生植物尤其是水生蔬菜以其较好的经济效益和环境效益正受到研究人员的广泛关注[6-8]。目前相关研究多集中于水生蔬菜对水体的净化效果方面[9-10],如莴笋、生菜等对氮的去除率可达66.45%~93.74%,对磷的去除率可达85.62%~97.48%[9];然而,关于水生蔬菜在富营养化水体中的生理响应及营养品质等方面的报道较少。因此,水生蔬菜能否安全用于富营养化水体的生态修复尚不明确。
水芹(OenanthejavanicaDC.)为伞形科水芹菜属水生蔬菜,喜干凉,忌炎热干燥,营养丰富,是中国中部和南部地区冬季的主要蔬菜之一。本试验以水芹为材料,通过浮床栽培技术,研究水芹的营养品质和抗氧化系统在富营养化水体中的变化,明确富营养化水体对水芹生长和营养品质的影响,为水生蔬菜进行富营养化水体的生态修复提供理论依据。
1 材料和方法
1.1 试验材料
供试水芹品种为赛雪白芹,由天津科润蔬菜研究所提供,叶柄嫩白粗壮,实心;叶片绿色,香味浓,肉质细腻;耐寒性、抗病性较好,易栽培,产量高,经济效益好。是四川地区主要推广品种之一。
1.2 试验设计
试验于2016年10~12月在四川农业大学温江校区的塑料大棚内进行。试验用水为人工配置富营养化水体,水体氮、磷由(NH4)2SO4、KNO3和KH2PO4提供,其余营养成分根据1/2Hoagland营养液进行配制[11]。水体中氮、磷初试浓度以四川省成都市温江区主要河流中氮、磷的实际测定值为依据来确定。本试验共设低、中、高3个浓度梯度(均属于劣V类水,中度或重度富营养化),并以1/2Hoagland营养液栽培水芹(正常栽培)为对照。具体浓度见表1。
种子经消毒、浸种、催芽后播于装有蛭石的穴盘中。气候条件为亚热带温润季风气候,年平均气温15.9 ℃。根据基质干湿程度浇灌营养液,当幼苗长出5~6片真叶时用清水清洗根系,先于1 mmol·L-1CaSO4·2H2O溶液中饥饿培养2 d,然后选择生长状况良好、长势基本一致的水芹移栽至上述3种程度富营养化和对照水体中。植株培养采用浮床栽培的方式,选用40 cm×30 cm×20 cm塑料水箱为栽培容器,每个水箱种植12株,每个处理重复3次。试验期间用去离子水补充水分,保持水量不变。观察并记录其生长状况,30 d后集中采样进行各项指标测定。
1.3 测定项目及方法
1.3.1生物量每处理随机抽取10株,将植株从水箱中取出,用自来水冲洗干净,然后用去离子水多次冲洗,擦干后分成地上部和根系两部分,分别称重。
1.3.2营养品质维生素C(Vc)含量测定采用2,6-二氯靛酚滴定法[12],可溶性糖含量测定采用蒽酮比色法[13];蛋白质含量测定采用考马斯亮蓝G-250法[13];亚硝酸盐含量测定采用分光光度法(GB 5009. 3-2010)。
1.3.3抗氧化酶活性超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)法[14],以抑制氮蓝四唑光还原50%为一个酶活性单位(U);过氧化物酶(POD)活性测定采用愈创木酚法[14],以吸光度OD470每分钟增加0.01为一个酶活性单位(U);抗坏血酸过氧化物酶(APX)活性采用紫外吸收法[14]测定,以吸光度OD290每分钟变化0.01为一个酶活性单位(U);过氧化氢酶(CAT)活性采用紫外分光光度法[13]测定,以使吸光度OD240每分钟减少0.1为一个酶活性单位(U)。酶活性均以U·mg-1表示。

表1 富营养化水体中氮磷初始浓度
1.3.4丙二醛含量及电解质渗透率丙二醛(MDA)含量的测定采用硫代巴比妥酸法(TBA)[12],以μmol·g-1表示MDA含量。电解质渗透率采用电导率仪法[12]测定。
1.4 数据处理
采用SPSS17.0和Excel 2010软件对数据进行统计分析,并利用LSD法进行差异显著性检验(P<0.05),结果以平均值±标准差(Mean±SD)表示。
2 结果与分析
2.1 不同程度富营养化水体对水芹生长的影响
从表2可知,在富营养化水体中,水芹地上部、地下部和全株最终鲜重在T1、T2处理间均无显著差异,但两处理均显著低于相应对照(CK);而T3处理水芹地上部、地下部和全株最终鲜重均显著高于T1、T2处理,但是与对照无显著差异。同时,各处理水芹生物量在富营养化水体中生长30 d后均有显著的增加,T1~T3处理的全株最终鲜重是全株初始鲜重的4.18~7.00倍,且以T3处理和对照增幅更大。说明水芹可以在富营养化水体中正常生长,且氮、磷浓度的增加有利于水芹的生长。
2.2 不同程度富营养化水体对水芹抗氧化酶活性的影响
图1显示,在富营养化水体中生长30 d后,水芹叶片的SOD、POD活性在T1~T3处理间均无显著差异,且三者均显著高于对照,分别为对照的2.12~2.33倍和1.69~2.03倍;叶片中CAT、APX活性在T1~T3处理与相应对照间均无显著差异。同时,在富营养化水体中生长30 d后,T1和T2处理的水芹根系SOD活性显著高于对照,而T3处理居中,且与T1、T2处理及对照均无显著差异;T1~T3处理水芹根系的POD活性均显著高于对照,为对照的1.74~2.24倍,其中又以T1处理的POD活性最高,显著高于T2与T3处理;水芹根系中CAT、APX活性均与对照无显著差异。另外,与叶片相比较,水芹根系中SOD、POD、APX活性明显较高,而根系中CAT活性明显较低。说明在富营养化水体中,水芹叶片和根系均能维持较高水平的抗氧化酶活性。

表2 富营养化水体中水芹单株鲜重的变化
注:LSD法进行差异显著性检验,同列中不同字母表示处理间在0.05水平存在显著差异
Note: Significance test with least significant difference (LSD) test, different letters in the same columns mean significant differences among different treatments at 0.05 level
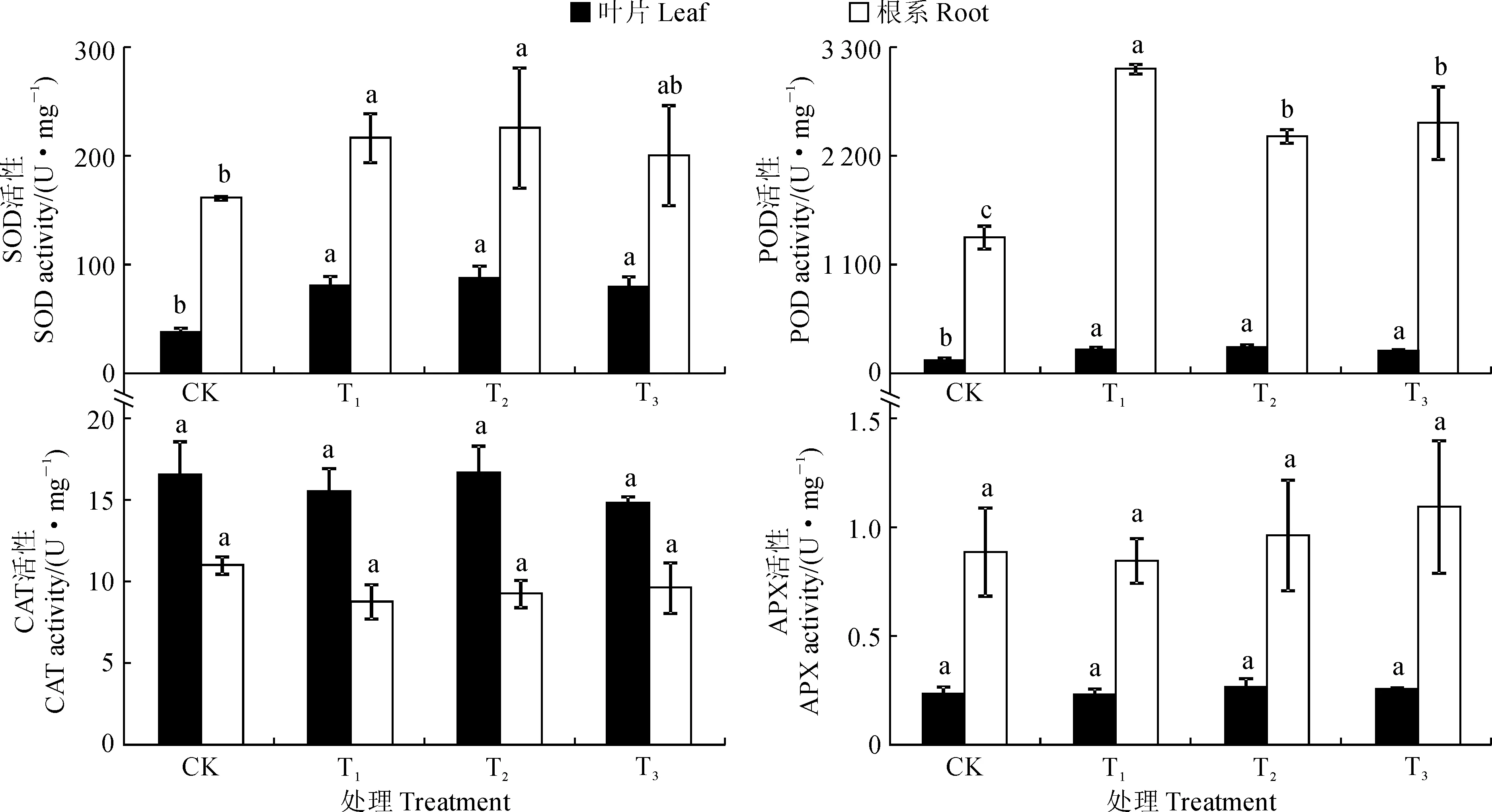
LSD法进行差异显著性检验,不同字母表示处理间在0.05水平存在显著差异,下同图1 不同程度富营养化水体中水芹抗氧化酶活性的变化Significance test with least significant difference (LSD) test, different letters mean significant differences between different treatments at 0.05 levels, the same as belowFig.1 Changes of antioxidant enzyme activities of O. javanica DC. in different eutrophic water
2.3 不同程度富营养化水体对水芹MDA含量及电解质渗透率的影响
由图2可以看到,在3种程度富营养化水体中生长30 d后,水芹叶片的MDA含量和电解质渗透率均与对照无显著差异,且水芹根系的MDA含量和电解质渗透率也与对照无显著差异。说明水芹在3种程度富营养化水体中均未受到明显的氧化伤害。
2.4 不同程度富营养化水体对水芹营养品质的影响
从图3来看,在不同程度富营养化水体中生长30 d后,T1~T3处理中水芹叶片的可溶性蛋白含量有随着氮、磷浓度的增加逐渐升高的趋势,但均显著低于对照,为对照的78%~85%;而水芹叶片中可溶性糖含量虽然逐渐降低,但是与对照无差异;水芹叶片中的Vc含量显著高于对照,为对照的1.10~1.34倍。同时,在3种程度富营养化水体中生长30 d后,水芹叶片中亚硝酸盐(以亚硝酸钠计)含量随着氮、磷浓度的增加而逐渐降低,并以T1处理最高(为对照的1.70倍),且显著高于对照,而T2~T3处理与对照无显著差异。另外,各处理水芹叶片中亚硝酸盐含量均低于GB2762-2017《食品中污染物限量》中不得大于20 mg·kg-1的国家标准。可见,富营养化水体中氮、磷浓度在一定范围内的升高有利于水芹可溶性蛋白和Vc的合成与积累,而其叶片中亚硝酸盐含量又处于国标安全范围内。

图2 不同程度富营养化水体中水芹MDA及电解质渗透率的变化Fig.2 Changes of MDA content and electrolyte leakage of O. javanica DC. in different eutrophic water

图3 不同程度富营养化水体中水芹营养品质及亚硝酸盐含量的变化Fig.3 Changes of nutritional quality and nitrite content of O. javanica DC. in different eutrophic water
3 讨 论
氮、磷是水体富营养化的2个重要因子,也是植物所必需的大量营养元素,它们在环境中的浓度会影响植物的生长和代谢。本试验中,随着水体富营养化程度的增加,水芹生物量呈增加的趋势,这与周晓红等[15]的研究结果一致;水芹幼苗在富营养化水体中生长30 d后,生物量均有大幅度提高,全株鲜重增长4.18~7.00倍,且水芹在高浓度富营养化水体中的生物量与正常栽培条件下无差异。说明水体氮、磷浓度的增加有利于促进水芹的生长发育,水芹可以在富营养化水体中正常生长。
对于湖泊、河流、水库等淡水资源来说,无机态氮浓度大于1 mg·L-1,PO43--P浓度大于0.1 mg·L-1即达到水体富营养化的标准(SL395-2007)。然而,富营养化水体中氮磷浓度远低于水生蔬菜的正常所需。因此,富营养化水体对于水生蔬菜而言可能是一种营养缺乏的环境。在正常环境条件下,植物体内的活性氧代谢处在不断变化的动态平衡之中;而在逆境胁迫下(如营养过剩或缺乏),活性氧产生和消除的平衡遭到破坏,活性氧大量积累,造成膜脂过氧化伤害[16];同时植物会启动SOD、POD、CAT和APX等抗氧化酶清除活性氧,减轻或消除膜脂过氧化伤害[17]。SOD是植物抗氧化系统的主导酶,它的主要功能是催化超氧阴离子生成H2O2,而POD与CAT、APX均可以催化H2O2分解成水和氧气,彻底解除活性氧的毒害。本试验中,各浓度富营养化水体处理的水芹叶片和根系中SOD活性显著高于对照,这与和华龙等[18]对凤眼莲的研究结果相似。徐华林[19]发现,低浓度氮、磷处理的鸭舌草POD活性显著高于高浓度氮、磷处理。与此相似,本试验中水芹叶片和根系的POD活性均显著高于对照,这可能是因为各浓度处理中水芹较高的SOD活性,致使H2O2含量增加,即POD的底物浓度增加,刺激了POD活性升高[20]。据报道,不同浓度硝态氮处理下,金鱼藻CAT和APX活性先升高后降低又升高,且各处理间差异显著[21]。而本试验中水芹叶片和根系中CAT和APX活性均与对照无显著差异,这与苦草中的相关研究结果相似[22],说明在H2O2的清除过程中,主要是POD发挥了重要作用。抗氧化酶活性的变化表明,水芹清除活性氧的能力较强。另外,MDA含量及电解质渗透率是反映植物受逆境胁迫程度重要指标[20]。本试验中,各浓度处理的水芹叶片和根系的MDA含量和电解质渗透率均与对照无显著差异,这与青萍对富营养化水体的响应[23]相似,说明在3种程度富营养化水体中,水芹均未发生膜脂过氧化损伤。水芹体内抗氧化系统的变化表明,本试验中氮、磷浓度对水芹而言可能是一种营养缺乏的逆境胁迫,致使水芹体内出现了轻度的活性氧代谢失调;而水芹通过增强抗氧化酶(SOD和POD在此过程中发挥了重要作用)活性,清除过多的活性氧,以避免膜脂过氧化伤害。因此,水芹对3种程度富营养化水体具有较好的适应性,生理代谢受影响较小。
氮、磷是影响蔬菜中各类营养物质合成的主要因素。蔬菜中的可溶性蛋白质、可溶性糖和Vc含有人体所需的氨基酸、供能物质、抗氧化物质等重要营养物质,它们的含量直接反映了蔬菜营养品质的好坏[24]。研究发现,增施氮、磷有利于植物可溶性蛋白含量的提高[25]。本试验中随着水体中氮、磷浓度的增加,水芹叶片中可溶性蛋白质含量呈升高趋势,这与蔡东升等[26]研究的番茄品质在不同氮磷水平下的变化结果相似。其原因可能是由于氮作为蛋白质的主要成分,直接影响着可溶性蛋白质的含量,磷则通过参与氮素代谢来促进可溶性蛋白质的合成[24]。而氮、磷对植物可溶性糖的影响较为复杂,氮、磷均有利于可溶性糖的积累,同时磷也可以促进植物对氮的吸收与利用,所以氮、磷的浓度及其比例共同影响着植物可溶性糖的积累[27]。水芹叶片可溶性糖含量在各浓度富营养化水体处理间无显著差异,说明本试验的氮、磷条件未对水芹可溶性糖的积累产生明显的促进。水芹叶片的Vc含量随着氮、磷浓度的增加先升高后降低,但都显著高于对照,说明适量的氮、磷供应有利于蔬菜Vc含量的提高[28]。蔬菜中的亚硝酸盐对人体存在潜在的危害,可诱发消化系统癌变等严重疾病,而蔬菜是人体摄入亚硝酸盐的主要来源[29],所以蔬菜中亚硝酸盐含量是蔬菜的重要品质和安全性指标之一。本研究中随着水体富营养化程度的增加,水芹的亚硝酸盐含量降低,且远低于国家标准(GB2762-2017)。这可能是因为随着水体中氮、磷含量的升高,促进了水芹氮素代谢,有利于降低水芹的亚硝酸盐含量[30]。可见,在富营养化水体中,水芹的营养品质与对照无显著差异,在本试验环境下水芹的营养品质较好,符合国家允许的食用标准。
综上所述,水芹可以在3种程度富营养化水体中正常生长,且氮、磷浓度增加有利于水芹的生长;在3种富营养化水体中,水芹的SOD和POD活性保持较高水平,电解质渗透率和MDA含量均维持在较低水平,并未发生膜脂过氧化伤害;在3种富营养化水体中,水芹营养品质较好,符合国家允许的食用标准。因此,水芹对富营养化水体具有较好的适应性,可用于富营养化水体的生态修复。
参考文献:
[1] 许其功, 曹金玲, 高如泰, 等. 我国湖泊水质恶化趋势及富营养化控制阶段划分[J]. 环境科学与技术, 2011,34(11):147-151.
XU Q G, CAO J L, GAO R T,etal. Trend of water quality deterioration and eutrophication control phases partition in China [J].EnvironmentalScience&Technology, 2011,34(11): 147- 151.
[2] 陈小锋, 揣小明, 杨柳燕. 中国典型湖区湖泊富营养化现状、历史演变趋势及成因分析[J]. 生态与农村环境学报, 2014,30(4): 438-443.
CHEN X F, CHUAI X M, YANG L Y. Status Quo, Historical evolution and causes of eutrophication in lakes in typical lake regions of China[J].JournalofEcologyandRuralEnvironment, 2014,30(4): 438-443.
[4] YANG CX, YANG JR, LI H. Improvement of inflow water quality of Dian Lake treated by constructed wetlands[J].AdvancedMaterialsResearch, 2013,613: 1 409-1 415.
[5] ZHAO F L, YANG W D, ZENG Z,etal. Nutrient removal efficiency and biomass production of different bioenergy plants in hypereutrophic water[J].BiomassandBioenergy, 2012,42(7): 212-218.
[6] 胡绵好, 袁菊红, 杨肖娥. 水生蔬菜对富营养化水体净化及资源化利用[J]. 湖泊科学, 2010,22(3): 416-420.
HU M H, YUAN J H, YANG X E. Eutrophication purification and resource utilization by aquatic vegetables[J].JournalofLakeScience, 2010,22(3): 416-420.
[7] 李发荣, 余 冬. 应用水培蔬菜净化滇池富营养水体分析研究[J]. 环境科学导刊, 2012,31(5): 32-34.
LI F R, YU D. A research on purification of the eutrophicated water by hydroponic vegetables in Dianchi Lake[J].EnvironmentalScienceSurvey, 2012,31(5): 32-34.
[8] 罗 竞, 柯卫东, 刘玉平, 等. 水生蔬菜在水体修复中的应用概况[J]. 长江蔬菜, 2013,18: 37-41.
LUO J, KE W D, LIU Y P,etal. Application of aquatic vegetables in water restoration[J].JournalofChangjiangVegetables, 2013,18: 37-41.
[9] 刘晓丹, 李 军, 龚一富, 等. 5种水培植物对富营养化水体的净化能力[J]. 环境工程学报, 2013,7(7): 2 607-2 612.
LIU X D, LI J, GONG Y F,etal. Purification of eutrophic water by five aqua-cultured plants[J].ChineseJournalofEnvironmentalEngineering, 2013,7(7): 2 607-2 612.
[10] HU M H, AO Y S, YANG X E,etal. Treating eutrophic water for nutrient reduction using an aquatic macrophyte (IpomoeaaquaticForsskal) in a deep flow technique system[J].AgriculturalWaterManagement, 2008,95(5): 607-615.
[11] 何 娜, 孙占祥, 张玉龙, 等. 不同水生植物去除水体氮磷的效果[J]. 环境工程学报, 2013,7(4): 1 295-1 300.
HE N, SUN Z X, ZHANG Y L,etal. Efficiency of removing nitrogen and phosphorus from simulated wastewater using hydrophytes[J].ChineseJournalofEnvironmentalEngineering, 2013,7(4): 1 295-1 300.
[12] 蔡庆生. 植物生理学实验[M]. 北京: 中国农业大学出版社, 2013: 41-42+ 166-179.
[13] 汤绍虎, 罗 充. 植物生理学实验教程[M]. 重庆: 西南师范大学出版社, 2012: 113-131.
[14] MA T T, CHRISTIE P, LUO Y M,etal. Physiological and antioxidant responses of germinating mung bean seedlings to phthalate esters in soil[J].Pedosphere, 2014,24(1): 107-115.
[15] 周晓红, 王国祥, 杨 飞. 两种富营养化水体对植物生长及光合荧光特性的影响[J]. 生态环境学报, 2011,20(2): 337-344.
ZHOU X H, WANG G X, YANG F. Effects of eutrophication water on plant growth and photosynthetic fluorescence characteristics[J].EcologyandEnvironmentalSciences, 2011,20(2): 337-344.
[16] FOYER C H, SHIGEOKA S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis[J].PlantPhysiology, 2011,155(1): 93-100.
[17] GILL S S, KHAN N A, TUTEJA N. Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulfur assimilation and antioxidant machinery in garden cress (LepidiumsativumL.)[J].PlantScience, 2012,182: 112-120.
[18] 和华龙, 黄 华, 薛建辉. 模拟酸雨和富营养化复合胁迫对水葫芦抗氧化酶的影响[J]. 江苏农业科学, 2015,43(11): 430-432.
HE H L, HUANG H, XUE J H. Effects of simulated acid rain and eutrophication on antioxidant enzymes ofEichhorniacrassipes(Mart.) Solms[J].JiangsuAgriculturalSciences, 2015,43(11): 430-432.
[19] 徐华林. 扬州地区挺水观赏植物资源及其利用价值研究[D]. 江苏扬州: 扬州大学, 2015.
[20] 贾永霞, 李 弦, 张长峰, 等. 羽衣甘蓝对镉的耐性和富集特征研究[J]. 西北植物学报, 2015,35(5): 971-977.
JIA Y X, LI X, ZHANG C F,etal. Cadmium tolerance and accumulation characteristics ofBrassicaoleraceaL. var.acephalaDC. [J].ActaBotanicaBoreali-OccidentaliaSinica, 2015,35(5): 971-977.
[21] 曹 特, 倪乐意. 金鱼藻抗氧化酶对水体无机氮升高的响应[J]. 水生生物学报, 2004,28(3): 299-303.
CAO T, NI L Y. Responses of antioxidases ofCeratophyllumdemersumto the increase of inorganic nitrogen in water column [J].ActaHydrobiologicaSinica, 2004,28(3): 299-303.
[22] 王 敏. 附植藻类对太湖流域常见沉水植物的影响研究[D]. 南京: 南京信息工程大学, 2013.
[23] 宋关玲, 侯文华, 汪群慧, 等. 适用于青萍修复的水体富营养化状况研究[J]. 哈尔滨工业大学学报, 2006,38(10): 1 793-1 796.
SONG G L, HOU W H, WANG Q H,etal. Suitable eutrophication level for restoration byLemnaminor[J].JournalofHarbinInstituteofTechnology, 2006,38(10): 1 793-1 796.
[24] 王正银. 蔬菜营养与品质[M]. 北京: 科学出版社, 2009: 2, 7, 68-70.
[25] 孙常青, 杨艳君, 郭志利, 等. 施肥和密度对杂交谷可溶性糖、可溶性蛋白及硝酸还原酶的影响[J]. 植物营养与肥料学报, 2015,21(5): 1 169-1 177.
SUN C Q, YANG Y J, GUO Z L,etal. Effects of fertilization and density on soluble sugar and protein and nitrate reductase of hybrid foxtail millet[J].JournalofPlantNutritionandFertilizer, 2015,21(5): 1 169-1 177.
[26] 蔡东升, 李建明, 樊翔宇, 等. 基质栽培营养液氮磷钾补充水平对番茄养分吸收及产量品质影响[J]. 东北农业大学学报, 2017,48(1): 7-14.
CAI D S, LI J M, FAN X Y,etal. Effect of nitrogen, phosphorus and potassium supplementation on nutrient uptake, yield and quality of tomato in substrate culture[J].JournalofNortheastAgriculturalUniversity, 2017,48(1): 7-14.
[27] 杨俊年, 李彦杰, 胡廷章, 等. 低氮磷胁迫对五个玉米品种幼苗的影响[J]. 湖北农业科学, 2017,56(12): 2 257-2 260.
YANG J N, LI Y J, HU T Z,etal. Effects of low nitrogen and phosphorus stress on five kinds varieties of corn seedling[J].HubeiAgriculturalSciences, 2017,56(12): 2 257-2 260.
[28] MOZAFAR A. Nitrogen fertilizers and the amount of vitamins in plants: A review[J].JournalofPlantNutrition, 1993,16(12): 2 479-2 506.
[29] CHANG A C, YANG T Y, RISKOWSKI G L. Changes in nitrate and nitrite concentrations over 24 h for sweet basil and scallions[J].FoodChemistry, 2013,136(2): 955-960.
[30] 王玉波, 李彩凤, 马凤鸣, 等. 氮磷钾对甜菜硝酸还原酶与亚硝酸还原酶的影响[J]. 核农学报, 2012,26(5): 803-808.
WANG Y B, LI C F, MA F M,etal. Effect of NPK on NR and NIR activity of sugar beet[J].JournalofNuclearAgriculturalSciences, 2012,26(5): 803-808.
猜你喜欢
海洋通报(2022年4期)2022-10-10
青春期健康(2022年9期)2022-05-16
新农业(2020年18期)2021-01-07
皮革制作与环保科技(2020年14期)2020-03-17
天然产物研究与开发(2019年1期)2019-03-01
中成药(2018年8期)2018-08-29
中国农业文摘-农业工程(2016年5期)2016-04-12
应用海洋学学报(2015年2期)2015-11-22
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
