
干旱胁迫对日本荚蒾幼苗生理生化特性的影响
2018-04-23 12:53李瑞姣岳春雷李贺鹏陈献志
西北林学院学报 2018年2期
李瑞姣,岳春雷,李贺鹏*,王 珺,郭 亮,陈献志,杨 乐
(1.浙江农林大学 林业与生物技术学院,浙江 临安 311300; 2.浙江省林业科学研究院,浙江 杭州 310000; 3.浙江省台州市椒江区林业科技推广总站,浙江 台州 318000; 4.浙江省临海市林业技术推广总站,浙江 临海 317000)
在大气温室气体增加所导致的全球气候变化下,水资源匮乏日益严重,干旱半干旱地区范围增加,因此选择兼具良好观赏性和抗旱节水型的园林绿化植物具有十分重要的意义。干旱影响着植物形态、生理生化代谢及地理分布范围,是植物最容易受到的胁迫之一[1-2]。植物在干旱条件下会引起膜伤害和膜透性的增加,而体内抗氧化酶协同作用以及渗透调节物质,能够抵御或减少干旱胁迫对细胞的伤害[3-5]。此外,大多数植物在干旱条件下组织相对含水量都会有所降低[6-7],生物量的积累及分配格局也会发生变化[8],但不同植物对干旱逆境的响应机制和适应能力都有差异。因此,从多方面系统研究,明确植物对不同强度干旱胁迫的响应和适应机制,对其进行资源保护及绿化应用都十分重要。
日本荚蒾(Viburnumjaponicum)为忍冬科(Caprifoliaceae)荚蒾属(Viburnum)常绿灌木,叶革质,浓绿而光亮,花密集白色,果实熟时红色,经冬不凋,是优良的观赏花木。日本荚蒾在我国仅在浙江省部分海岛发现有分布记录[9-10],但海岛地区土壤贫瘠,降雨量偏少,植物生存环境恶劣,野生植株数量十分稀少,已被列入浙江省重点保护野生植物名录(第一批)和浙江省极小种群拯救保护物种。虽然目前已经有对日本荚蒾开展人工引种驯化,在南京城市公园绿化建设也有种植[11],但若进一步进行园林绿化栽植或沿海防护林建设等推广应用,必须了解其抗旱能力及抗旱机理,但目前对这方面的研究尚未见报道。因此,本试验采用盆栽控水模拟不同程度的土壤水分状况,通过测定持续干旱胁迫下日本荚蒾幼苗的生物量分配、叶片相对含水量、抗氧化活性和渗透调节物质等变化特点,分析并讨论对干旱环境的适应能力及对策,为日本荚蒾的迁地保护及推广应用提供参考理论依据。
1 材料与方法
1.1 试验设计
试验在浙江省林业科学研究院苗圃大棚内进行。选用生长健壮、无病虫害、长势基本一致的2年生日本荚蒾实生幼苗作为供试材料。试验采用盆栽法,盆钵规格为21 cm×16 cm×21 cm(上径×下径×高),每盆土重约5 kg,栽培基质为苗圃熟土,有机质含量为38.26 g/kg,速氮0.16 g/kg,速磷0.061 g/kg,速钾0.014 g/kg,试验前用环刀法测得基质的最大田间持水量为31.9%。水分梯度划分为4个土壤水分水平:正常供水(Normal water supply,CK)、轻度干旱(Mild water stress,LS)、中度干旱(Moderate water stress,MS)、重度干旱(Severe water stress,SS),其土壤含水量分别为田间最大持水量的80%、60%、40%、20%,即土壤含水量分别为25.52%、19.14%、12.76%、6.38%。2016年6月将盆栽苗移入大棚内缓苗,期间正常供水。7月下旬不浇水待土壤水分分别自然消耗至设定标准,每天傍晚采用感量为千分之一的电子称称重补水,使各处理的土壤含水量维持在设定的水平,每个土壤水分处理设置6个重复。
1.2 测定指标与方法
1.2.1 生理生化指标的测定 各处理于胁迫后第15、30、45、60、75天分别进行采样,叶片取样在早晨8:00-10:00进行,测定生理生化特征参数。叶片相对含水量(RWC)采用烘干法[12]测定;丙二醛(MDA)采用硫代巴比妥酸法测定[12];可溶性蛋白采用考马斯亮蓝染色法测定[12];可溶性糖采用蒽酮比色法测定[13];超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法测定[12];过氧化物酶(POD)活性采用愈创木酚比色法测定[13]。
1.2.2 生物量的测定 在胁迫处理75 d后,每个水分处理选3株,全株挖起,清洗干净、吸干水分,并将根、茎、叶分开,放入恒温箱中105℃杀青30 min,80℃烘至恒重并分别称重,计算总生物量、根冠比、根生物量比、茎生物量比、叶生物量比等。
1.2.3 数据分析 采用SPSS18.0软件对数据进行单因素方差(one-way ANOVA)分析,并用最小显著法(Duncan's)进行多重比较。用Origin 8.5软件绘图。
2 结果与分析
2.1 干旱胁迫对日本荚蒾幼苗叶片相对含水量(RWC)的影响
由图1可知,随着土壤含水量的减少日本荚蒾幼苗叶片的RWC逐渐降低,LS与CK处理在胁迫期间均无显著差异(P>0.05),MS处理仅在胁迫60 d、75 d与CK有显著差异(P>0.05),分别降低了16.88%和11.84%。SS在整个处理过程中叶片逐渐萎蔫、枯萎,RWC比CK分别降低了10.75%、24.94%、22.61%、35.43%,且差异均显著(P<0.05),在胁迫75 d,植株全部干枯死亡。除SS外,其他处理RWC随胁迫时间延长逐渐上升。
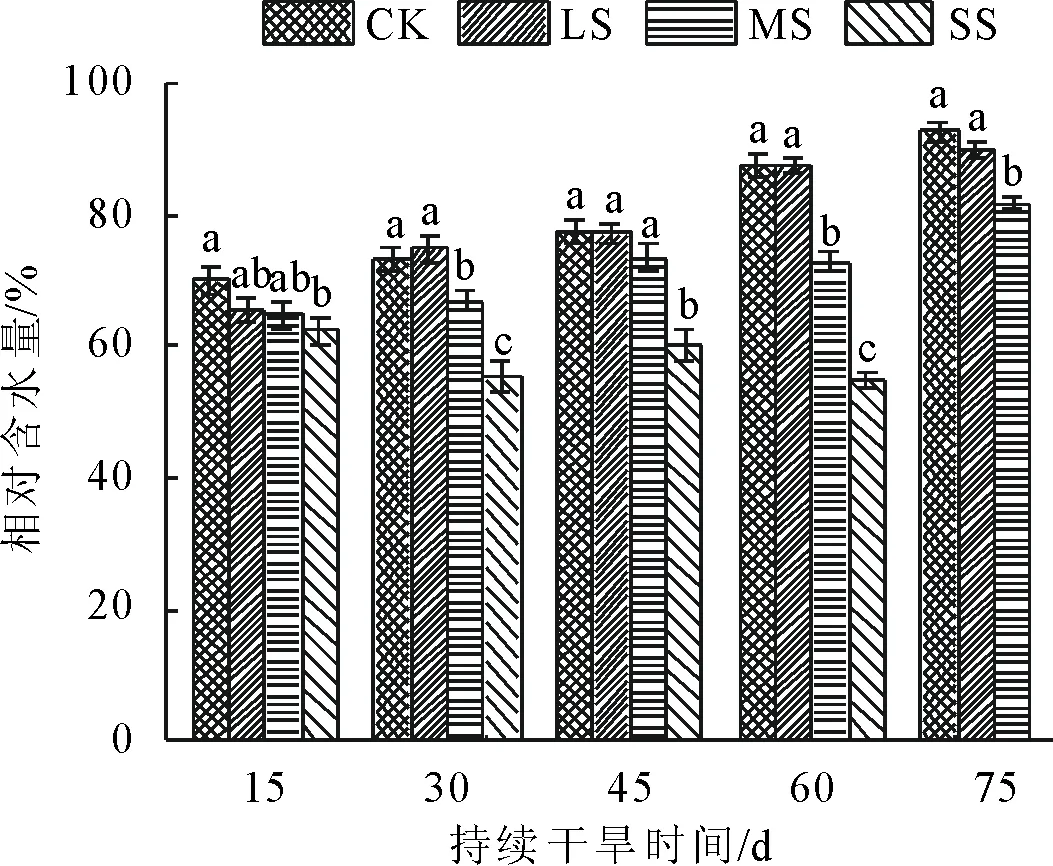
注:不同小写字母表示各处理在0.05水平有显著差异。下同。
图1不同水分处理下日本荚蒾幼苗叶片相对含水量的变化
Fig.1 Relative water content ofV.japonicumseedlings under different soil water treatments
2.2 干旱胁迫对日本荚蒾幼苗叶片丙二醛(MDA)含量的影响
由图2可知,日本荚蒾幼苗在受到干旱胁迫时叶片丙二醛(MDA)含量均比CK有所增加,且干旱胁迫越重MDA含量增加的越多,LS在胁迫期间与CK相比增幅为1.4%~23.8%,MS增幅为7.8%~38.8%,SS增幅为27.7%~64.8%,且SS与CK间差异均显著(P<0.05)。

图2 不同水分处理下日本荚蒾幼苗MDA含量的变化
2.3 干旱胁迫对日本荚蒾幼苗叶片渗透调节物质的影响
由图3可知,各干旱处理下日本荚蒾叶片的可溶性蛋白含量均高于CK,在胁迫45 d及以前,随着土壤含水量的减少可溶性蛋白含量逐渐上升,SS处理比CK增加了52.3%、75.5%、102.9%,但在胁迫60 d,LS、MS仍逐渐升高,SS虽高于CK但比MS有所减小。可溶性糖含量随土壤含水量的减少逐渐升高,LS、MS在胁迫过程中与CK均有显著性差异(P<0.05),SS处理在胁迫60 d有所降低,与CK差异不显著(P>0.05)。

图3 不同水分处理下日本荚蒾幼苗可溶性蛋白、可溶性糖含量的变化
2.4 干旱胁迫对日本荚蒾幼苗叶片抗氧化酶活性的影响
SOD、POD是生物体内重要的抗氧化酶。由图4可知,LS、MS处理在胁迫过程中的SOD活性均比CK有所增加,且随土壤水分含量的减少而增加,在胁迫45 d及以后,CK、LS、MS之间差异不显著(P>0.05)。SS处理的SOD活性在胁迫15 d时达到最大值,比CK增加了61.5%,之后值逐渐降低,胁迫60 d比CK减少了19.6%。LS、MS处理下日本荚蒾叶片的POD活性除在胁迫15 d、75 d与CK有显著差异外(P<0.05),其他阶段变化均不明显。SS处理叶片POD活性在胁迫15 d比CK有所降低,但在30 d、45 d大幅增加,分别增加至CK的2.23倍和3.10倍,在胁迫60 d又比CK降低了75.1%。
2.5 干旱胁迫对日本荚蒾幼苗生物量积累及分配的影响
由表1可知,日本荚蒾幼苗的生物量积累在不同的水分梯度下发生了明显改变,随胁迫程度的加强总生物量逐渐减小,且MS、SS与 CK处理间差异显著(P>0.05),SS较CK降低了33.78%。生物量的分配方式也存在着明显的不同,3种干旱处理下茎生物量比均低于CK且差异明显(P<0.05);LS、MS的叶生物量比较CK有所上升,但SS下降了14.63%;而MS、SS的根生物量比则逐渐升高且与CK差异均明显(P<0.05)。生物量分配的这些变化直接导致不同强度干旱胁迫下日本荚蒾根冠比的差异,LS与CK间无明显差异(P>0.05),而MS和SS则分别提高了11.08%和42.12%,差异均显著(P<0.05)。

图4 不同水分处理下日本荚蒾幼苗SOD、POD活性的变化

处理总生物量/g根冠比根生物量比茎生物量比叶生物量比CK15.372a0.406c0.289c0.376a0.335bLS14.998a0.407c0.289c0.348b0.363aMS12.889b0.451b0.311b0.350b0.339bSS10.180c0.577a0.366a0.348b0.286c
注:不同小写字母分别表示各处理之间有显著差异(P<0.05)。
2.6 干旱胁迫下日本荚蒾幼苗叶片各个生理生化指标相关性分析
由表2可知,日本荚蒾幼苗叶片MDA含量与与可溶性蛋白呈极显著正相关关系,与总生物量呈极显著负相关关系,与相对含水量呈负相关关系。可溶性糖含量与可溶性蛋白含量呈极显著正相关。相对含水量与总生物量呈显著正相关,而与根冠比呈极显著负相关。总生物量与根冠比呈显著负相关关系。
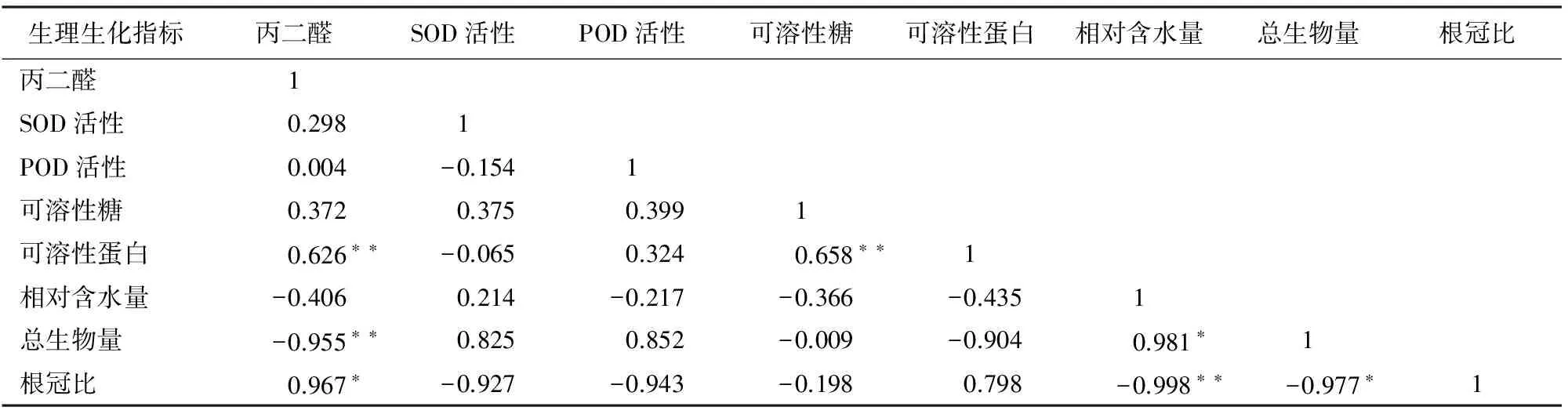
表2 干旱胁迫下日本荚蒾幼苗叶片各生理生化指标的相关系数
注:“-”表示负相关,“*”表示显著相关(P<0.05),“**”表示极显著相关(P<0.01)。
3 结论与讨论
干旱胁迫时植物正常的生理代谢会受到抑制,使形态特征、生理生化等方面发生一系列的变化,这是植物对环境胁迫的反应以及对环境改变的适应结果[14]。干旱胁迫对植物的伤害首先表现为膜脂过氧化[15],MDA是脂质过氧化的产物之一,其积累是活性氧毒害作用的表现。本研究中,随着土壤含水量的减少,日本荚蒾叶片的MDA含量逐渐升高,表明在干旱胁迫下,植物叶片代谢紊乱发生膜脂过氧化,质膜遭到损害,且随着胁迫程度的加重受伤害程度加深。轻度与中度干旱与正常供水处理相比叶片相对含水量(RWC)变化相对较小,但重度干旱在处理30 d便有大幅下降,说明植物在轻度、中度干旱环境下,通过减缓RWC的下降速度防止叶片萎蔫,且RWC与MDA呈负相关,保持较高的叶片含水量而减轻细胞受损程度,这是植物适应逆境生长的表现[16-17]。但长期的重度干旱导致植物叶片保水力严重下降,造成叶片枯萎、卷曲,质膜受到严重破坏。此外,除重度干旱外,其他处理RWC随胁迫时间延长逐渐升高,这可能是因为气温逐渐降低,叶片蒸发量减小,叶肉细胞自身的保水力上升所致[18]。
植物在抵御和适应不良环境时,自身还会形成一些防御机制,如启动抗氧化系统、进行渗透调节等[19]。渗透调节是植物适应干旱环境的一种重要生理机制,植物通过积累细胞内溶质浓度降低渗透势,维持膨压,从而使体内各种与膨压有关的生理过程正常进行[8]。轻度和中度干旱处理下,日本荚蒾可溶性蛋白含量均随含水量的减少而增加,这与干旱胁迫下欧洲鹅耳枥[20]、栓皮栎[21]的研究结果一致,可能是因为土壤干旱胁迫促进某些特殊种类蛋白质的合成如“逆境蛋白”或“胁迫蛋白”或使原有蛋白质含量增加,提高细胞渗透调节能力[5]。此外可溶性糖含量也比正常供水有所增加,植物通过增加细胞原生质浓度共同抵御逆境胁迫[14]。随着土壤水分的减少,重度干旱处理叶片可溶性糖虽有增加但增幅逐渐减少,且胁迫60 d可溶性蛋白、可溶性糖含量均低于MS,可能是因为长期严重干旱导致蛋白质、可溶性糖合成代谢受阻或降解增加[20,22-23],植物渗透调节能力降低或不再增加[24],膜脂过氧化加剧,细胞结构和功能受到严重破坏。胁迫过程中可溶性蛋白变化幅度比可溶性糖更明显,且相关性分析中可溶性蛋白含量与MDA呈极显著正相关,说明可溶性蛋白比可溶性糖发挥着更大的渗透调节作用。
植物在逆境时维持较高的抗氧化酶活性,有利于清除活性氧,减少膜脂过氧化程度,从而减轻质膜损伤[19]。SOD、POD是细胞内清除活性氧的主要保护酶,彼此协调综合作用才能发挥保护酶系统的防御能力。本研究中,不同的水分处理下,日本荚蒾叶片酶活性变化有所不同。干旱胁迫初期,轻度和中度处理POD酶活性较SOD变化更为明显,说明POD对迅速轻度、中度干旱环境更敏感,随着胁迫时间延长,这两种酶均保持较高的活性,减缓植物受伤害程度。重度干旱处理下,胁迫15 d叶片SOD大幅升高POD则比CK明显降低,之后SOD酶逐渐降低而POD则大幅升高。表明重度干旱初期,叶片主要依赖SOD消除伤害,而重度胁迫中期使SOD酶活性下降而POD发挥主要作用,协同作用提高植物的防御能力。植物存在一个耐受胁迫的极限,随胁迫程度的加强酶活性逐渐降低[25-26],胁迫60 d重度干旱处理SOD、POD均大幅下降,表明重度干旱胁迫下自由基产生和清除失衡,超出了防御系统的清除能力,膜系统发生过氧化反应破坏膜结构[5,27]。此外本研究发现,结合生理指标的变化,重度干旱处理在胁迫60 d下MDA保持较高,RWC、POD均为最低,SOD、可溶性蛋白、可溶性糖下降至较低,植物防御机制能力均处于下降状态,说明在土壤水分含量保持6.38%情况持续60 d时,日本荚蒾生理生化特性发生紊乱,对干旱环境无抵抗能力和适应性,达到植物维持生长的阈值,当胁迫持续下去植物逐渐死亡,但如果对植物进行复水,增加土壤含水量,其是否能够恢复生长还有待进一步研究。
干旱胁迫大多抑制植物的生长,而且生物量的降低幅度和不同器官之间生物量分配与植物干旱适应能力密切相关[28-30]。本研究中,轻度、中度干旱下日本荚蒾保持较高的叶生物量比例维持光合作用的正常进行,保证足够的生物量积累[31]。重度干旱下为了满足植物对水分的需求,将生长中心向根系转移,根生物量比和根冠比显著上升,试图从土壤中吸收更多的水分[8],但土壤中可用水量过分减少,植物叶片萎蔫、焦枯,叶生物量所占比例大幅降低,导致光合作用严重降低,植物生长受到严重影响,总生物量明显下降。
综合结果表明,随土壤含水量的减少,日本荚蒾幼苗受胁迫程度逐渐增加,但在轻中度干旱环境中植物具有较强的抗氧化活性和渗透调节能力,叶片相对含水量下降缓慢,同时保持较高的叶生物量比维持植物的光合能力,增强对干旱逆境的耐受性和适应性。重度干旱环境下,植物防御机制发生紊乱,细胞膜遭到严重破坏,叶片干枯,生长受到严重影响,且当在土壤含水量为6.38%的情况持续60 d,植物达到抵抗胁迫的极限,生命力转向负值。相较于其他常用绿化植物,香樟在土壤含水量长期低于7%会受到明显伤害[32],普陀樟在土壤含水量为12.21%~14.25%下有50%的死亡率[33],日本荚蒾对干旱具有相当甚至更强的抵抗能力,因此该植物在浙江地区园林绿化和沿海造林中具有一定的推广价值。
参考文献:
[1] 李霞,阎秀峰,于涛.水分胁迫对黄檗幼苗保护酶活性及脂质过氧化作用的影响[J].应用生态学报,2005,16(12):2353-2356.
LI X,YAN X F,YU T.Effects of water stress on protective enzyme activities and lipid peroxidation inPhellodendronamurenseseedlings[J].Chinese Journal of Applied Ecology,2005,16(12):2353-2356.(in Chinese)
[2] 李毅,屈建军,张伟民,等.白刺对干旱胁迫的生理生化反应[J].中国农学通报,2012,28(1):24-29.
[3] REDDY A R,CHAITANYA K V,VIVEKANANDAN M.Drought induced responses of photosynthesis and antioxidant metabolism in higher plants[J].Journal of Plant Physiology,2004,161(11 ):1189-1202.
[4] WHITE D A,TUMER N C,GALBRAITH J H. Leaf water relations and stomatal behavior of four allopatrie Eucalyptus species planted in Mediterranean Southwestern Australia[J].Tree Physiology,2000, 20 (17 ):1157-1165.
[5] 范苏鲁,苑兆和,冯立娟,等.干旱胁迫对大丽花生理生化指标的影响[J].应用生态学报,2011,22(3):651-657.
FAN S L,YUAN Z H,FENG L J,etal.Effects of drought stress on physiological and biochemical parameters ofDahliapinnata[J].Chinese Journal of Applied Ecology,2011,22(3):651-657.(in Chinese)
[6] 李彦瑾,赵忠,孙德祥,等.干旱胁迫下柠条锦鸡儿的水分生理特征[J],西北林学院学报,2008,23(3):1-4.
LI Y J,ZHAO Z,SUN D X,etal.Hydrological physiological characteristics ofCaraganakorshinskiiunder water stress[J].Journal of Northwest Forestry University,2008,23(3):1-4.(in Chinese)
[7] 张凤良,毛常丽,李小琴,等.橡胶树优树无性系对干旱胁迫的生理响应[J].西北林学院学报,2016,31(4):67-72.
ZHANG F L,MAO C L,LI X Q,etal.Physiological responses ofHeveabrasiliensisplus tree clones to drought stress[J].Journal of Northwest Forestry University,2016,31(4):67-72.(in Chinese)
[8] 安玉艳,梁宗锁,郝文芳.杠柳幼苗对不同强度干旱胁迫的生长与生理响应[J].生态学报,2011,31(3):716-725.
AN Y Y,LIANG Z S,HAO W F.Growth and physiological responses of thePeriplocasepiumBunge seedlings to drought stress[J].Acta Ecologica Sinica,2011,31(3):716-725.(in Chinese)
[9] 裘宝林,陈征海,张晓华.见于浙江的中国及中国大陆新记录植物[J].云南植物研究,1994,16(3):231-234.
[10] 朱弘,葛斌杰,叶喜阳.浙江舟山东福山岛种子植物区系初探[J].浙江农林大学学报,2015,32(1):150-155.
ZHU H,GE B J,YE X Y.Seed plant flora of Dongfushan Island in Zhoushan,Zhejiang Province[J].Journal of Zhejiang A&F University,2015,32(1):150-155.(in Chinese)
[11] 张凤凤,王静,郝日明.忍冬科植物在南京园林中的应用及其前景[J].江苏林业科技,2009,36(2):48-51.
[12] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[13] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[14] 苏慧敏,何丙辉,蔡兴华,等.水分胁迫对太阳扇扦插苗形态和生理特征的影响[J].生态学杂志,2011,30(10):2185-2190.
[15] 彭远英,颜红海,郭来春,等.燕麦属不同倍性种质资源抗旱性状评价及筛选[J].生态学报,2011,31(9):2478-2491.
PENG Y Y,YAN H H,GUO L C,etal.Evaluation and selectionon drought-resistance of germplasm resources ofAvenaspecies with different types of ploidy[J].Acta Ecologica Sinica,2011,31(9):2478-2491.(in Chinese)
[16] 胡义,胡庭兴,胡红玲,等.干旱胁迫对香樟幼树生长及光合特性的影响[J].应用与环境生物学报,2014,20(4):675-682.
HU Y,HU T X,HU H L,etal.Effects of drought stress on growth and photosynthetic characteristics ofCinnamomumcamphoraSaplings[J].Chinese Journal of Applied and Environmental Biology,2014,20(4):675-682.(in Chinese)
[17] 欧毅,王进,吴天强,等.水分胁迫对桃形李叶片含水量、质膜透性和抗氧化酶活性的影响[J].西南农业学报,2007,20(5):982-985.
OU Y,WANG J,WU T Q,etal.Effects of water stress on water content,relative membrance permeability and the activity of antioxidant enzymes of plum leaves[J].Southwest China Journal of Agricultural Sciences,2007,20(5):982-985.(in Chinese)
[18] 张娜,赵宝平,郭若龙,等.水分胁迫对不同抗旱性燕麦品种生理特性的影响[J].麦类作物学报,2012,32(1):150-156.
ZHANG N,ZHAO B P,GUO R L,etal.Effect of water stress on physiological characteristics of different oat cultivars[J].Journal of T riticeae Crops,2012,32(1):150-156.(in Chinese)
[19] 王兴顺.黄芩(Scutellariabaicalensis)幼苗对干旱胁迫的生理适应性反应[J].西北林学院学报,2014,29(1):55-59.
WANG X S.Physiological responses ofScutellariabaicalensisseedlings under drought stress[J].Journal of Northwest Forestry University,2014,29(1):55-59.(in Chinese)
[20] 王飒,周琦,祝遵凌.干旱胁迫对欧洲鹅耳枥幼苗生理生化特征的影响[J].西北植物学报,2013,33(12):2459-2466.
WANG S,ZHOU Q,ZHU Z L.Physiological and biochem ica l characteristics ofCarpinusbetulusseedlings under drought stress[J].Acta Botanica Boreali-Occidentalia Sinica,2013,33(12):2459-2466.(in Chinese)
[21] 崔豫川,张文辉,王校锋.栓皮栎幼苗对土壤干旱胁迫的生理响应[J].西北植物学报,2013,33(2):364-370.
[22] 张旭东,王智威,韩清芳,等.玉米早期根系构型及其生理特性对土壤水分的响应[J].生态学报,2016,36(10):2969-2977.
ZHANG X D,WANG Z W,HAN Q F,etal.Effects of water stress on the root structure and physiological characteristics of early-stage maize[J].Acta Ecologica Sinica,2016,36(10):2969-2977.(in Chinese)
[23] 朱万泽,王金锡,薛建辉..台湾桤木和四川桤木种源苗木对水分胁迫的生理响应[J].西北植物学报,2005,25(10):1969-1975.
[24] 鲍思伟,谈锋,廖志华.土壤干旱对蚕豆叶片渗透调节能力的影响[J].西南农业大学学报,2001,23(4):353-355,359.
BAO S W,TAN F,LIAO Z H.Effects of soil drought on the osmoregulating ability ofViciafabaleaves[J].Journal of Southwest Agricultural University,2001,23(4):353-355,359.(in Chinese)
[25] 张刚,魏典典,邬佳宝,等.干旱胁迫下不同种源文冠果幼苗的生理反应及其抗旱性分析[J].西北林学院学报,2014,29(1):1-7.
ZHANG G,WEI D D,WU J B,etal..Effects of drought stress on physiological characteristics ofXanthocerassorbifoliafrom different provenances and analysis on drought-resistance in seedling stage[J].Journal of Northwest Forestry University,2014,29(1):1-7.(in Chinese)
[26] 任磊,赵夏陆,许靖,等.4种茶菊对干旱胁迫的形态和生理响应[J].生态学报,2015,35(15):5131-5139.
REN L,ZHAO X L,XU J,etal.Varied morphological and physiological responses to drought stress among four teaChrysanthemumcultivars[J]. Acta Ecologica Sinica,2015, 35 (15):5131-5139. (in Chinese)
[27] 周宝利,孟兆华,李娟,等.水分胁迫下嫁接对茄子生长及其生理生化指标的影响[J].生态学杂志,2012,31(11):2804-2809.
[28] 桑子阳.红花玉兰花部性状多样性分析与抗旱性研究[D].北京:北京林业大学,2011.
[29] YIN C Y,WANG X,DUAN B L,etal.Early growth,dry matter allocation and water use efficiency of two sympatricPopulusspeciesas affected by water stress[J].Environmental and Experimental Botany,2005,53:315-322.
[30] 邱娟,谭敦炎,樊大勇.准噶尔荒漠早春短命植物的光合特性及生物量分配特点[J].植物生态学报,2007,31(5):883-891.
QIU J,TAN D Y,FAN D Y.Characteristics of photosynthesis and biomass allocation of spring ephemerals in the Junggar Desert[J].Acta Phytoecologica Sinica,2007,31(5):883-891.(in Chinese)
[31] 关保华,葛滢,樊梅英,等.华荠苧响应不同土壤水分的表型可塑性[J].生态学报,2003,23(2):259-263.
GUAN B H,GE Y,FAN M Y,etal.Phenotypic plasticity of growth and morphology inMoslachinensisresponds to diverse relative soil water content[J].Acta Ecologica Sinica,2003,23(2):259-263.(in Chinese)
[32] 胡义,胡庭兴,陈洪,等.干旱胁迫及复水对香樟幼树生理特性及生长的影响[J].西北植物学报,2015,35(2):294-301.
HU Y,HU T X,CHEN H,etal.Physiological properties and growth ofCinnamomumcamphorasaplings under drought stress and rewatering[J].Acta Botanica Boreali-Occidentalia Sinica,2015,35(2):294-301.(in Chinese)
[33] 陈闻,赵颖,叶正钱,等. 干旱胁迫对5个海岛树种生长及生理特性的影响[J].浙江农林大学学报,2013,30(4):490-498.
CHEN W,ZHAO Y,YE Z Q,etal.Growth and physiological characteristics of five island tree species with drought stress[J].Journal of Zhejiang A & F University,2013,30(4):490-498.(in Chinese)
猜你喜欢
今日农业(2020年19期)2020-12-14
音乐研究(2019年5期)2019-11-22
湖南饲料(2019年4期)2019-10-17
种子(2019年8期)2019-09-11
中国生物医学工程学报(2019年5期)2019-07-16
Coco薇(2017年5期)2017-06-05
中国饲料(2016年12期)2016-11-07
中国眼镜科技杂志(2016年18期)2016-11-03
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
