
西藏自治区冷杉林生物量分布格局及模型研建
2018-04-20 09:58:37
中南林业科技大学学报 2018年5期
(国家林业局 中南林业调查规划设计院,湖南 长沙 410014)
森林是陆地上面积最大的生态系统,其生物量是全球碳循环的重要组成部分,在减缓气候变化方面发挥了积极的作用[1-2]。西藏位于地球“第三极”青藏高原的西南部,自然条件复杂,森林资源丰富,是我国森林碳储量最大的省份之一[3-6]。西藏乔木林中,冷杉林面积、蓄积最大,分别占乔木林面积和蓄积的30%、50%左右[7],储存了大量的有机碳。开展冷杉林生物量、碳储量的研究,了解其在时间、空间上的变化规律,对于加强西藏森林资源保护与管理、发挥高原生态安全屏障功能提供了重要的数据支撑。
鉴于森林资源连续清查数据具有系统性、连续性和可比性强的特点,本研究在充分利用清查数据对全区森林资源碳密度及分布规律研究[8]的基础上,将冷杉林生物量与植被的生长阶段(林龄)、森林结构(胸径、树高、株数密度)等林分因子及经度、纬度、海拔、坡度、坡向、坡位等地理因子结合在一起,探寻地理因子与林分因子对森林生物量的协同影响。试图进行:(1)分析冷杉林生物量变化的驱动因子是什么,这些因子在多大程度上影响着生物量的变化;(2)探索冷杉林生物量的分布格局,即在地理空间上的分布状况及随林龄变化的消长规律;(3)在提高多因子林分生物量密度模型的可靠性方面进行探索;(4)鉴于特殊的自然地理条件导致西藏自治区森林资源连续清查布设的固定样地中有一定数量的遥感影像判读样地和目测调查样地[9],建立适合该两类样地的生物量模型,实现全区范围的冷杉林生物量估算。
1 研究区概况
西藏自治区位于祖国的西南边陲,青藏高原 西 南 部, 地 处 东 经 78°15′~ 99°07′、 北 纬26°50′~ 36°29′之间,东西长约 2020 km,南北宽约1060 km,陆地国境线长约4000 km。北抵昆仑山、唐古拉山,与新疆维吾尔自治区及青海省毗邻,东隔金沙江与四川省相望,东南与云南省山水相连,西临克什米尔地区,南与印度、尼泊尔、锡金、不丹、缅甸接壤。西藏自治区土地面积约占全国土地总面积的八分之一,平均海拔4000 m以上,地势高峻,地理位置特殊,素有“世界屋脊”和“地球第三极”之称。这里不仅是南亚、东南亚地区的“江河源”和“生态源”,还是中国乃至东半球气候的“启动器”和“调节区”。
2 研究方法
2.1 数据来源
基础数据为2011年第八次全国森林资源连续清查西藏自治区复查实测样地数据[10],实测样地1462个,其中冷杉有林地共90个样地。按地区分布情况为:林芝地区79个样地,昌都地区3个样地,日喀则地区4个样地,山南地区4个样地。数据包括样地基本记录和每木检尺记录。样地数据包括样地高斯投影坐标、海拔高度、坡度、坡向、坡位、林分起源、优势树种、林分平均年龄、龄组、平均胸径、平均树高、郁闭度、公顷株数等。样木数据包括样地中样木的立木类型、检尺类型、树种、胸径、材积等。
2.2 数据处理
2.2.1 实测样地生物量的计算
利用样地的胸径每木检尺数据,结合样地平均胸径、平均树高,利用相对树高曲线估测单株树高[11-12],进而推算单株生物量[13],合并计算样地生物量。
2.2.2 生物量密度影响因子的筛选
地理因子中,经度、纬度、海拔反映了光照、水分及温度条件的变化。坡度影响了植株分布密度,同时通过影响土壤厚度等因子间接影响生物量密度。地貌、坡位、坡向3个因子为定性变量,取值非连续性分布。其中坡向按照光照强弱和时长对坡向进行赋值,分南、东南、西南、东、西、西北、东北和北分别赋予权重1.0、0.9、0.9、0.8、 0.8、0.7、0.7、0.6,无坡向赋值1.0[14]。地貌、坡位2个因子可以通过海拔得到体现,故在影响因子筛选时将其剔除[15]。
平均胸径、平均树高、郁闭度与林木生长密切相关,反映了林木材积和生物量状况;林龄反映了林木的生长发育阶段;在西藏天然林为主体的森林结构中,株数密度反映了林分的自我发育状况与自疏程度。
2.3 模型的构建
使用SPSS17.0软件,以样地数据库为分析对象,以林分生物量密度为因变量(Y),以地理因子和林分因子为自变量(Xi,i=1、2、3、…)进行分析,构建多元线性模型或非线性模型,探讨自然环境和林分生物学特性对生物量密度的影响[15]。
2.3.1 模型形式的选择
线性:y=β0+β1x2+β2x2+…+βpxp+ε。
式中:y为因变量(生物量密度);xp为自变量;β0为截距,即常量;β1、β2、…、βp为偏回归系数,表明自变量对因变量的影响程度;ε为其它随机因素引起的y的变化部分。
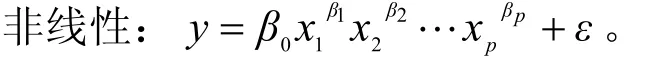
式中:y为因变量(生物量密度);xp为自变量;β0、β1、β2、…、βp为模型参数,表明自变量对因变量的影响程度;ε为其它随机因素引起的y的变化部分。
2.3.2 自变量的选择
利用SPSS17.0软件,使用多元线性回归的逐步回归法,根据设定的判据重复进行自变量的引入和剔除,直至回归方程中的自变量均符合进入模型的判据,即所有引入的自变量对因变量都有显著影响而不需要剔除,而模型外的自变量都不符合进入模型的判据。
2.3.3 模型评价指标
采用决定系数(R2)、估计值的标准差(SEE)、总相对误差(TRE)、平均系统误差(MSE)、平均预估误差(MPE)和平均百分标准误差(MPSE)等6项指标作为模型的评价指标[16],其计算公式如下:


式中:yi为实测观测值;为模型预估值;为样本平均值;n为样本单元数;p为参数个数;tα为置信水平α时的t值。这些指标中,R2和估计值的标准差是回归模型的最常用指标,反映了模型的拟合优度;总相对误差和平均系统误差是反映拟合效果的重要指标,二者都应该控制在一定范围内(如±3%或±5%),趋于0时效果最好;平均预估误差是反映平均生物量估计值的精度指标;平均百分标准误差是反映平均单个样地林分生物量估计值的精度指标。
3 研究结果
以林分生物量密度为因变量,通过不同林分因子和地理因子的组合进行多元线性逐步回归,结果表明:按照Probability ofF<= 0.05引入变量、Probability ofF>=0 .10剔除变量的原则,最终选用的变量为平均胸径、郁闭度,生物量密度随平均胸径和郁闭度的增加而增加。将平均胸径、郁闭度作为自变量进行非线性拟合,构建的线性和非线性模型的拟合结果见表1。
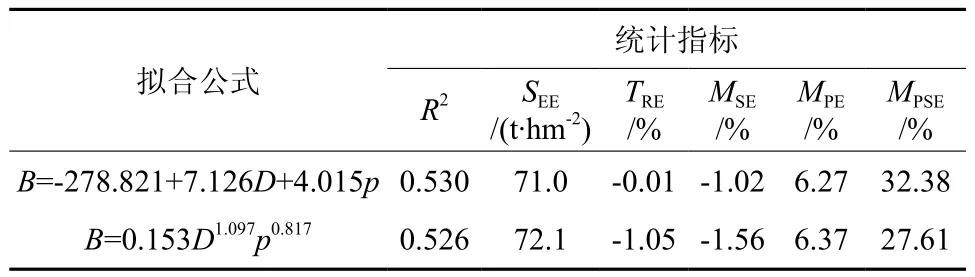
表1 生物量密度模型的拟合结果及其统计指标†Table 1 Fitting results of biomass density model and its statistical indicators
从表1的各项统计指标可以看出,线性和非线性模型的拟合结果均较为理想,拟合模型的R2分别为0.530和0.526,说明胸径和郁闭度解释了生物量变动的50%以上;估计值的标准差分别为71.0和72.1 t/hm2,这是计算预估误差的基础信息;总相对误差和平均系统误差均控制在±2%以内,表明模型拟合效果良好;平均预估误差分别为6.27%和6.37%,表明模型的平均预估精度达到93%以上;平均百分标准误差反映了单个样地生物量估计误差的平均水平。2种模型比较来看,线性模型的R2、估计值的标准差、总相对误差、平均系统误差、平均预估误差均优于非线性模型,但由于线性模型各个自变量对目标变量的贡献是按照线性累加的,很难避免出现不合理的预估结果,比如生物量估计值出现负值[17]。由表1线性模型的公式可以看出,当胸径和郁闭度较小时,生物量密度会出现负值。这也导致了线性模型的平均百分标准误差高于非线性模型,即线性模型对单个样地生物量估计误差要大于非线性模型,因此建议在进行冷杉林生物量估计时使用非线性模型。
因胸径在很大程度上反映了林木材积和生物量状况,在通过逐步回归法筛选自变量时可能将其它影响生物量密度的因子掩盖;同时,在遥感和目测样地或小班中,准确地估计胸径也存在一定的技术限制,因此在引入自变量时尝试对胸径进行了剔除,探寻生物量密度在时间和空间上的变化规律。重新以林分生物量密度为因变量,将剔除胸径后的林分因子和地理因子作为自变量,进行多元线性逐步回归,最终选用的变量为林龄、海拔、郁闭度,其中生物量密度随林龄和郁闭度的增加而增加,随海拔的升高而减少。以上述3个自变量为基础建立非线性模型,构建的线性和非线性模型的拟合结果见表2。
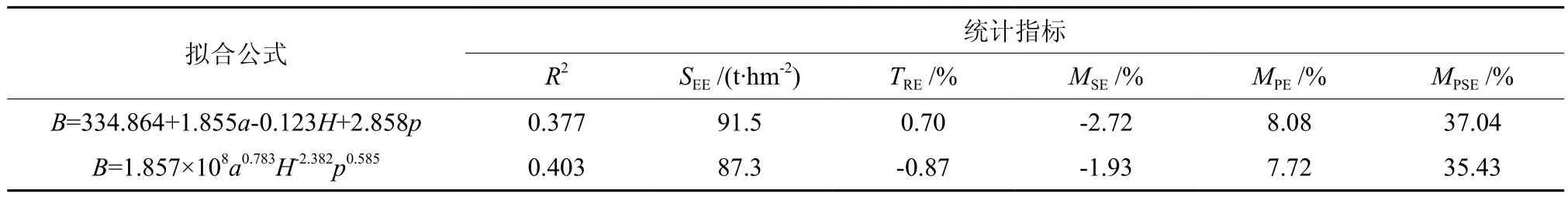
表2 生物量密度模型的拟合结果及其统计指标†Table 2 Fitting results of biomass density model and its statistical indicators
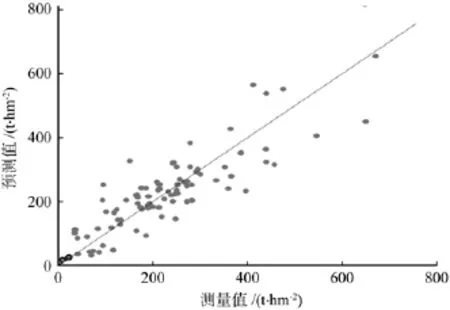
图1 样地预测值和测量值的比较(以胸径和郁闭度为自变量)Fig.1 Comparison of predicted and measured values of the plots (with DBH and canopy closureas independent variables)
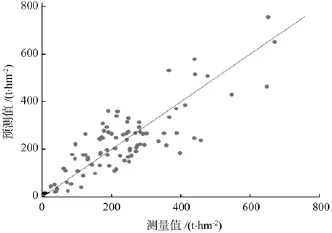
图2 样地预测值和测量值的比较(以林龄、郁闭度和海拔为自变量)Fig.2 Comparison of predicted and measured values of the plots (with forest age, canopy closure andaltitude as independent variables)
对比表1与表2,剔除胸径而选用林龄、海拔、郁闭度作为自变量后,方程的拟合优度和生物量估计精度均有一定程度的下降,但模型的平均预估精度仍可达到92%左右,预估精度下降有限。为了进一步确认模型的拟合效果和通用性,绘制了生物量测量值与模型预估值的散点分布图(见图1、图2)。由图1和图2可以看出,以林龄、郁闭度和海拔为自变量建立的模型与以胸径和郁闭度为自变量建立的模型相比,预测值的分布略为分散,但各个预测值都基本均匀分布在y=x的直线两侧,即模型的残差近似随机分布,不存在明显的系统偏差,这对于保证模型的通用性是至关重要的。在估算区域冷杉林生物量或碳储量时,可优先采用以胸径、郁闭度为自变量建立的模型;对于目测和遥感样地,由于难以准确获取平均胸径数值,可采用以林龄、海拔、郁闭度为自变量建立的模型。同时,若需进行某个时间段内冷杉林生物量变化或碳汇的估算,并了解生物量在地理空间上的分配规律,需要引入林龄、海拔作为自变量,即使用以林龄、海拔、郁闭度为自变量建立的模型。
4 讨 论
4.1 海拔的影响
在同一地区,暗针叶林的生产量与海拔高度有着密切的联系。冷杉林的生产量在海拔3500 m附近最高,随着海拔的增加和水分条件的减少,冷杉树高变矮,干形趋于尖削,且枝桠横生[18]。李文华等[22]通过对不同海拔高度冷杉生长过程的对比,发现各测树指标的生长量随海拔的增加而降低,特别是在接近森林上限附近,海拔的微小差异都会引起生长状况的急剧变化。马和平等[19]在色季拉山的调查发现,在3700~4300 m 海拔范围内冷杉林郁闭度随海拔高度的增加而减小。刘新圣等[20]在色季拉山沿海拔梯度选择了具有相似林龄(150~200 a)的冷杉林设置调查样地,表明随着海拔的升高,林分郁闭度、林木密度和胸高断面积均呈降低趋势,从而导致冷杉群落地上生物量呈现相应的下降趋势,其降低的平均幅度为73.1 t/(hm2·100 m)。从亚高山森林到林线,逐渐严酷的环境条件直接影响到冷杉个体的生长发育乃至整个群落的结构,导致林分生物量随海拔的升高而降低。决定暗针叶林分布最重要的热量指标是最热月的温度,其分布上限处为10 ℃,下限处为15℃(19℃)[21],因此冷杉生物量随海拔的变化规律可能与亚高山地带垂直方向温度的变化有关。从根据气象台站数据建立的热量指标与海拔高度的方程来看,年平均气温、最热月平均气温和最冷月平均气温与海拔高度的相关系数均在0.9以上,且三者均随海拔的升高而呈直线递减,海拔每升高100 m的递减值为0.57~0.61 ℃。在同等降水分布水平上,随着温度的降低,林分生产力呈指数递减,从而产生了冷杉林生物量随海拔升高而递减的分布规律。
4.2 林分特征因子的影响
李文华等[22]在对全国冷杉林生物生产力格局的研究中发现,随着林龄的增长,生物量递增,而生产力则递减。本研究对生物量密度与林龄的模型显示,幂函数的拟合效果好于线性函数,表明随着林龄的增加,冷杉林生物量增加速度加快,这可能是由于西藏湿润热带和亚热带的亚高山地带独特的自然地理环境,产生了暗针叶林独特的高产机制与经久性生长机制,数量成熟期很长,生长持续增长的潜力很大。对西藏冷杉生长特性的研究发现,亚高山地带冷杉具有很长的生长持续期,所研究的大量解析木中,绝大多数林木材积生长量仍处于持续的上升阶段,极少发现材积平均生长量下降的情况[18]。在相似的自然条件与林分年龄的情况下,立地条件差异对树木生长的影响可以通过平均胸径、郁闭度等因子体现,生物量密度与胸径、郁闭度的正相关关系研究较多,在此不再赘述。
5 结 论
(1)利用西藏自治区森林资源连续清查冷杉林样地数据,分析了影响冷杉林生物量密度的因子,包括胸径、郁闭度、林龄、海拔。生物量密度随胸径、林龄和郁闭度的增加而增加,随海拔的升高而减少。
(2)估算冷杉林生物量或碳储量时,实测样地可使用胸径、郁闭度建立的生物量模型,即B=0.153D1.097p0.817;对于目测或遥感样地,可使用林龄、海拔、郁闭度为自变量建立的模型,即B=1.857×108a0.783H-2.382p0.585;在了解生物量或碳汇随时间、空间的变化规律时,建议使用林龄、海拔、郁闭度为自变量建立的模型,即B=1.857×108a0.783H-2.382p0.585。
参考文献:
[1] Dixon R K, Brown S, Houghton R A,et al.Carbon pools and fl ux of global forest ecosystems[J]. Science, 1994, 263: 185-190.
[2] Scurlock J M O, Cramer W, Olson R J,et al.Terrestrial NPP:Towards a consistent data set for global model evaluation[J].Ecological Applications, 1999(9): 913-919.
[3] 李海奎,雷渊才,曾伟生.基于森林清查资料的中国森林植被碳储量[J].林业科学,2011,47(7):7-12.
[4] 吴庆标,王效科,段晓男,等. 中国森林生态系统植被固碳现状和潜力[J].生态学报,2008,28(2):517-524.
[5] 赵 敏,周广胜.中国森林生态系统的植物碳贮量及其影响因子分析[J].地理科学,2004,24(1):50-54.
[6] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16.
[7] 贺东北,柯善新,杨 宁,等.西藏自治区第二次森林资源规划设计调查成果报告[R].2014.
[8] 桂来庭,张 蓓,吴南飞,等. 西藏自治区森林碳密度及分布规律研究[J].中南林业调查规划,2014,33(3):34-36.
[9] 曾伟生,廖志云.西藏天然林直径分布模型及其应用研究[J].中南林业调查规划,2008,27(3):4-7.
[10] 国家林业局中南森林资源监测中心,西藏自治区林业厅.第八次全国森林资源清查西藏自治区森林资源清查成果[R].2012.
[11] 曾伟生. 西藏自治区相对树高曲线模型的研制[J].林业资源管理,2002(5):27-30.
[12] 李海奎,法 蕾. 基于分级的全国主要树种树高-胸径曲线模型 [J].林业科学,2011,47(10):83-90.
[13] 李海奎,雷渊才,曾伟生.基于森林清查资料的中国森林植被碳储量[J].林业科学,2011,47(7):7-12.
[14] 肖兴威. 影响亚热带东部森林结构的因子分析[J]. 东北林业大学学报,2004,32(5):19-20,34.
[15] 齐建文,张 蓓,刘金山. 湖南杉木林生物量密度的模拟与预测[J]. 中南林业科技大学学报,2014,34(8):15-18.
[16] 曾伟生,唐守正.立木生物量方程的优度评价和精度分析[J].林业科学,2011,47(11):106-113.
[17] 曾伟生,肖前辉,胡 觉,等. 中国南方马尾松立木生物量模型研建[J]. 中南林业科技大学报,2010,30(5):50-56.
[18] 李文华. 西藏森林[M].北京:科学出版社,1985.
[19] 马和平,郭其强,刘合满,等.藏东南色季拉山西坡土壤有机碳库研究[J].生态学报,2013,33(10) : 3122-3128.
[20] 刘新圣,张 林,孔高强,等. 藏东南色季拉山急尖长苞冷杉林线地带地上生物量随海拔的变化特征[J].山地学报,2011,29(3): 362-368.
[21] 李文华, 周沛村. 暗针叶林在欧亚大陆分布的基本规律及其数学模型的研究[J]. 自然环境,1979,1(1): 21-34.
[22] 李文华,罗天祥. 中国云冷杉林生物生产力格局及其数学模型[J].生态学报,1997,17(5):511-518.
猜你喜欢
吉林林业科技(2023年6期)2023-11-20 02:13:16
内蒙古林业调查设计(2021年5期)2022-01-05 02:50:40
防护林科技(2020年10期)2020-12-22 11:46:42
福建林业(2020年5期)2020-03-18 08:23:02
热带林业(2019年4期)2019-03-05 09:53:58
森林工程(2018年3期)2018-06-26 03:40:46
林业勘察设计(2017年4期)2017-07-06 08:46:12
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
