
小黑杨和桑树叶片光合特性对不同氮素形态的响应
2018-04-20 09:58:36金微微张会慧滕志远张书博张秀丽孙广玉
中南林业科技大学学报 2018年5期
金微微 ,张会慧 ,李 鑫 ,滕志远 ,张书博,张秀丽,许 楠 ,孙广玉
(1.东北林业大学 生命科学学院,黑龙江 哈尔滨 150040;2.东北农业大学 资源与环境学院,黑龙江 哈尔滨 150030;3. 黑龙江省科学院自然与生态研究所,黑龙江 哈尔滨 150040)
氮素的供应不足是植物生长最普遍的限制因素之一,氮在植物的生命活动中占有重要的地位[1-2]。植物可以利用的氮素形态主要有硝态氮(NO3--N)、铵态氮(NH4+-N)和有机态氮(如尿素)等[3-4],但植物体内大多数的氮都是以NO-3-N或NH4+-N的形式吸收的[5],NO3--N和NH4+-N的性质不同,植物体对不同形态氮的吸收能力及利用能力也不相同[6],并且不同氮形态明显影响植物的生长及生理特性[7-8]。光亮桉Eucalyptus nitens[9]、美洲山核桃Carya illinoinensis和白云杉Picea glauca[10]等一些树种对NH4+-N的吸收作用大于NO3--N,但水青冈Fagus longipetiolataSeem和桑树Morus alba等一些树种则优先选择吸收NO3--N[11-12],一些植物在单一NH4+-N下还会表现出铵盐毒害现象[13]。因此,研究不同植物对不同氮素形态的生理响应对指导合理施肥具有重要的意义。
目前,关于不同形态的氮素对木本植物生理特性方面的研究相对农作物还比较薄弱。小黑杨是我国北方常见的造林和绿化树种[14],桑树为南桑北移后我国北方地区推广的主要经济树种之一,二者均具有极强的抗旱和盐碱能力[15-16]。除了绿化外,二者在退化生态地区均可作为植被恢复的先锋树种。在退化生态地区土壤中氮缺乏一直是限制植物恢复的重要限制因素之一[17-18],因此研究二者对不同氮素形态的生理响应具有重要意义。植物叶片中的氮大部分以酶的形式存在于叶绿体和线粒体中,特别是叶绿体内的氮大约占到总氮量的75%左右[19],为此,本文中分别研究了不同氮素形态下小黑杨和桑树幼苗叶片的光合气体交换特性、叶绿素荧光特性以及光合能量分配特点,以期为指导退化生态氮素缺乏地区植被恢复过程中的树种合理配置提供理论依据,并为研究树木的合理施肥提供一些基础数据。
1 材料与方法
1.1 试验材料及处理方法
试验于2012年3—6月在东北林业大学植物生理学实验室进行。供试材料为一年生小黑杨和桑树幼苗,2种材料的株高均为30 cm左右,利用水培方法将2种材料培养于装有1/2Hoagland完全营养液直径30 cm、高50 cm的不透光水培养箱内,每水培养箱分别定植5株。用电动气泵连续通气,每3 d更换一次营养液。待水培苗长势稳定后进行不同氮素试验处理。在1/2Hoagland完全营养液本身提供的NO3--N 浓度为 7.5 mmol·L-1的基础上(NO3--N处理),将Hoagland完全营养液中的Ca(NO3)2·4H2O以同等氮浓度的(NH4)2SO4进行替换(NH4+-N处理),并将NH4+-N处理中的Ca2+浓度以CaCl2补充到NO3--N处理中的Ca2+浓度水平。待不同氮形态处理30 d后进行各生理指标的测定。
1.2 参数的测定及其方法
光合气体交换参数的测定:在环境温度23 ℃、相对湿度 75%、CO2浓度 450 μmol·moL-1温室条件下,利用Li-6400XT光合仪(Licor公司,美国)固定光照强度 1000 μmol·m-2s-1,各处理选择 3 片(倒数第3或第4)片完全展开叶,测定其净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)。3次重复。
叶绿素荧光参数的测定:利用暗适应夹对小黑杨和桑树幼苗叶片进行0.5 h的暗适应,采用便携式脉冲调制荧光仪FMS-2(Hansatch公司,英国),参照Hu等[20]方法测定PSⅡ最大光化学效率(Fv/Fm)、潜在光化学活性(Fv/Fo)、实际光化学效率(ФPSⅡ)和电子传递速率(ETR)、光化学淬灭系数(qP)和非光化学淬灭(NPQ),并计算PSⅡ反应中心吸收光能分配参数,即:用于光化学反应的量子产额(YPSⅡ)、依赖于类囊体膜两侧质子梯度和叶黄素循环的量子产额(YNPQ)、基本的荧光量子产额和热耗散的量子产额(Yf,D)及失活PSⅡ反应中心的热耗散量子产额(YNF),计算方法参照Hendrickson和Zhou等[21-22]。3次重复。利用FMS-2的内置光源分别测定经0.5 h暗适应后小黑杨和桑树幼苗叶片的ФPSⅡ、ETR光响应曲线,方法参照张会慧等[23],FMS-2施加作用光强分别为100、200、400、600、800 和 1200 μmol·m-2s-1, 每个作用光强下作用3 min后测定ФPSⅡ和ETR等数据。
1.3 数据处理方法
运用Excel和DPS(7.05)软件对测定数据进行统计分析,文中数据为3次重复的平均值±标准差(SE),并采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
2 结果与分析
2.1 小黑杨和桑树幼苗叶片光合气体交换参数对不同氮素形态的响应
图1可以看出,NH4+-N和NO3--N分别作为唯一氮源时,小黑杨幼苗叶片的Pn、Gs、Tr和Ci之间均无明显差异(P>0.05),但不同氮形态下,桑树幼苗叶片的光合气体交换参数却明显不同,NO3--N处理下桑树幼苗叶片的Pn、Gs和Tr分别高于NH4+-N处理的38.72%(P<0.01)、103.70(P<0.01)和73.33%(P<0.01),均达极显著差异水平,但2种氮素形态处理下桑树幼苗叶片的Ci却无显著差异(P>0.05)。
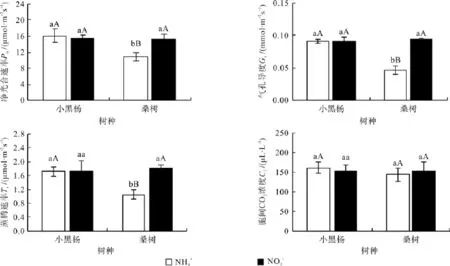
图1 小黑杨和桑树幼苗叶片光合气体交换参数对不同氮素形态的响应Fig.1 Responses of gas exchange parameters in leaves of Populus simonii×P. nigra and Morus alba seedlings to nitrogen form
2.2 小黑杨和桑树幼苗叶片叶绿素荧光特性对不同氮素形态的响应
由图2可以看出,NO3--N处理下小黑杨幼苗叶片ФPSⅡ、ETR、qP和NPQ分别高于NH4+-N处理的16.24%(P<0.05)、28.84%(P<0.01)、20.07%(P<0.01)和37.88%(P<0.05),但Fv/Fm和Fv/Fo未达显著差异水平。桑树幼苗叶片的各叶绿素荧光参数对2种氮形态的表现明显不同,其中NH4+-N处理下桑树幼苗叶片的Fv/Fm、Fv/Fo、ФPSⅡ、ETR和qP分 别 低 于 NO3--N处理的 10.16%(P< 0.01)、42.24%(P< 0.01)、55.53%(P< 0.01)、63.97%(P< 0.01) 和39.33%(P<0.01),而NPQ却高于NO3--N处理的100.46%(P<0.01),均达到极显著差异水平。
2.3 不同氮形态下小黑杨和桑树幼苗叶片叶绿素荧光参数的光响应曲线
由图3可以看出,随着光照强度的增加,2种氮形态下小黑杨和桑树幼苗叶片的ФPSⅡ均呈降低趋势,而ETR呈先增加后降低的趋势。不同光强下NH4+-N处理桑树幼苗叶片的ФPSⅡ均明显低于NO3--N 处理。100 ~ 200 μmol·m-2s-1弱光下,2 种氮形态下桑树幼苗叶片的ETR之间无明显差异,随着光强的增加,桑树幼苗叶片的ETR之间差异变得明显,当光强达 800和 1200 μmol·m-2s-1时,NH4+-N处理桑树幼苗叶片的ETR分别低于NO3--N处理的38.83%和55.30%。
2.4 小黑杨和桑树幼苗叶片PSⅡ反应中心光能分配参数对不同氮素形态的响应
由图4可以看出,2种氮素形态下小黑杨和桑树幼苗叶片的PSⅡ反应中心光能分配参数明显不同,其中小黑杨幼苗叶片PSⅡ反应中心光能分配参数在NO3--N和NH4+-N处理下差异较小,NH4+-N处理下的YPSⅡ低于NO3--N处理的8.19%,Yf,D高于NO3--N处理的7.77%,而YNPQ和YNF无明显差异。与小黑杨不同,桑树幼苗叶片PSⅡ反应中心光能分配参数对2种形态氮的响应明显不同,其中NH4+-N处理下的YPSⅡ低于NO3--N处理的29.56%,YNPQ和YNF分别高于NO3--N处理的20.35%和9.62%。
3 讨 论
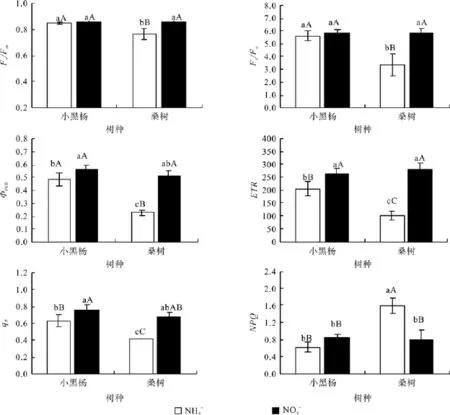
图2 小黑杨和桑树幼苗叶片叶绿素荧光特性对不同氮素形态的响应Fig.2 Responses of hlorophyll fl uorescence characteristics in leaves of Populus simonii×P.nigra and Morus alba seedlings to nitrogen forms
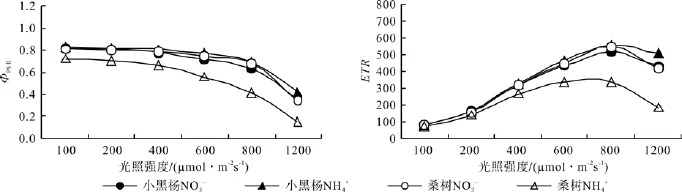
图3 不同氮形态下小黑杨和桑树幼苗叶片叶绿素荧光参数的光响应曲线Fig.3 Chlorophyll fl uorescence parameters response to illumination intensity in leaves of Populus simonii×P. nigra and Morus alba seedlings under different nitrogen forms
氮代谢和碳代谢是植物生命活动中最重要的两大代谢过程,二者常耦联在一起发挥作用[24]。光合作用是植物获取物质和能量的基础,氮肥的施用除了直接影响植物的生长及代谢过程外,还会影响到植物的光合作用,且不同形态氮的生理作用明显不同,有研究发现铁核桃Juglans sigillata实生苗在NO3--N处理下的光合色素含量以及净光合速率明显低于NH4+-N处理[25],并且在红松Pinus koraiensis幼苗的研究中也表现出相似的结果[26]。本试验结果发现,小黑杨幼苗叶片的各光合气体交换参数在NH4+-N和NO3--N处理下均无明显差异;桑树幼苗在NO3--N处理下叶片的Pn、Gs和Tr均明显高于NH4+-N处理,但2种氮处理下桑树幼苗叶片的Ci却无显著差异。说明2种氮源下小黑杨幼苗均能有效利用进行光合作用,而NH4+-N处理下桑树幼苗叶片的气孔因素不但限制了CO2进入叶肉细胞,而非气孔因素中叶肉细胞对CO2的利用能力也明显降低。这可能是由于当植物以NO3--N为唯一氮源时,若NO3--N量超过体内所需,多余的NO3--N会贮存在液泡中,并不影响植物的其他代谢过程;而以NH4+-N为唯一氮源时,NH4+容易扩散穿过生物膜,使膜去极化,阻碍质子驱动势的形成,进而影响同化力的形成,从而对CO2的固定产生抑制作用,降低光合速率[27]。以上结果表明在供应NO3--N的条件下桑树幼苗光合能力强于在供应NH4+-N的条件下。
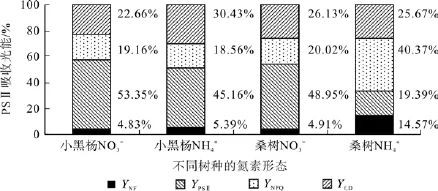
图4 小黑杨和桑树幼苗叶片PSⅡ反应中心光能分配参数对不同氮素形态的响应Fig.4 Responses of PS Ⅱ energy allocation pathway in leaves of Populus simonii×P.nigra and Morus alba seedlings to nitrogen forms
叶绿素荧光技术可分析植物光合机构的生理功能,目前已经广泛应用到植物生理学研究的各个方面,它弥补了光合气体交换参数在光反应研究中的不足。目前已有较多关于氮素形态对植物PSⅡ荧光特性的研究,张一利[28]发现相比NO3--N培养的黄瓜植株,NH4+-N处理下黄瓜植株叶片的ФPSⅡ、F′v/F′m以及光化学碎灭系数qP均有显著下降,但 2种氮素处理下水稻的ФPSⅡ、F′v/F′m、qP均没有明显差异。江力等[29]的研究指出,高氮水平下烟叶的光合放氧速率、光合电子传递活性都显著上升;低氮水平下烟草的光合放氧速率、光合电子传递活性显著下降。陈永亮等[30]的研究表明在NO3--N条件下红松幼苗叶PSⅡ、PSⅠ活性均明显高于NH4+-N处理。NO3--N处理的向日葵植株ФPSⅡ、qP和ETR都显著高于NH4+-N处理的植株;而非光化学猝灭系数则明显低于NH4+-N处理的植株[31]。本试验结果表明,在NO3--N条件下,小黑杨叶片的ФPSⅡ、ETR、qP显著高于NH4+-N处理,有利于碳同化力(ATP和NADPH)的生成,但由于光合作用受多种非气孔因素(如Rubisco活性等)双重影响,这种增高并未使小黑杨净光合速率的升高,且在不同光强下小黑杨叶片的ФPSⅡ、ETR、qP在2种氮形态处理下无明显差异,同时叶片PSⅡ反应中心光能分配参数在NO3--N、NH4+-N处理间差异较小,并结合2种氮形态下小黑杨叶片的Fv/Fm、Fv/Fm光能无显著差异,说明小黑杨在2种氮形态处理下PSⅡ反应中心的光化学效率相似。与小黑杨在2种不同氮形态处理条件下的响应不同,在NH4+-N处理下,与NO3--N处理相比,桑树的Fv/Fm、Fv/Fo、ФPSⅡ、ETR、qP显著降低,NPQ显著升高,这表明在NH4+-N处理下桑树PSⅡ光化学活性降低,而这种降低可能是由于氨毒害作用使光合磷酸化和光合电子传递解偶联[32],导致ATP的生成减少,碳同化所需的ATP供应进而减少,使碳同化速率减慢,导致碳同化利用的NADPH减少,造成NADPH的累积,没有充足的NADP+接受光合电子传递链传来的电子,光合电子传递链处于过度还原状态。PSⅡ反应中心激发的电子传递受阻,使PSⅡ反应中心关闭,使桑树叶片PSⅡ的光化学活性降低、过剩激发能积累增加,促进了桑树叶片PSⅡ激发能的非光化学耗散,这与对桑树PSⅡ反应中心光能分配参数的分析结果一致(YNPQ和YNF明显增加),导致YPSⅡ明显减少,而YPSⅡ减少反过来又会进一步减少同化力的形成,使桑树叶片的光合能力下降。而在桑树在NO3--N处理下,NO3--N对植物代谢活动不会产生不利影响,同时NO3-还原过程可利用由叶绿体中生成的苹果酸经双羧酸运转器运送到细胞质,再由苹果酸脱氢酶催化生成的NADH[33],同时直接利用还原型铁氧还蛋白(Fdred)传递的电子[11],Fdred向氮代谢分配的能量可以达光化学反应量子产额的20%~25%,在某些植物中甚至高达55%,这有助于增强桑树对光合激发电子的利用能力,使光合电子传递链正常运转。
4 结 论
在NO3--N、NH4+-N 2种不同的氮形态处理下,小黑杨光合特性总体无显著差异;桑树在NH4+-N处理下,与NO3--N处理相比,PSⅡ光化学活性明显下降,净光合速率显著下降,表明NH4+-N对桑树可能有毒害作用,NO3--N更有利于桑树的生长,碳同化受到抑制可能是NH4+-N处理下桑树净光合速率下降的主要原因。
参考文献:
[1] Burke I C, Lauenroth W K, Partow J. Regional and temporal variation in net primary production and nitrogen mineralization in grasslands[J]. Ecology, 1997, 78: 1330-1340.
[2] 童 琪, 李志辉, 童方平, 等. 南酸枣幼林配方施肥及叶片氮磷钾元素分布规律[J]. 中南林业科技大学学报, 2017, 37(5): 50-54.
[3] Kuelland K. Amino acid absorption by arctic plants: implications for plant nutrition and nitrogen cycling[J]. Ecology, 1994, 75:2373-2383.
[4] Chapin F S, Autumn K, Pugnaire F. Evolution of suites of traits in response to environmental stress[J]. The American Naturalist,1993, 142: 78-92.
[5] 王 磊, 隆小华, 郝连香, 等. 氮素形态对盐胁迫下菊芋幼苗PSⅡ光化学效率及抗氧化特性的影响[J]. 草业学报, 2012,21(1): 133-140.
[6] 张延春, 陈治锋, 龙怀玉, 等. 不同氮素形态及比例对烤烟长势、产量及部分品质因素的影响[J].植物营养与肥料学报,2005, 11(6): 787-792.
[7] Xie H L, Jiang R F, Zhang F S,et al. Effect of nitrogen form on the rhizosphere dynamics and uptake of cadmium and zinc by the hyperaccumulatorThlaspi caerulescens[J]. Plant Soil, 2009,318:205-21.
[8] 刘赵帆, 张国斌, 郁继华, 等. 氮肥形态及配比对花椰菜产量、品质和养分吸收的影响[J]. 应用生态学报,2013,24(7):1923-1930.
[9] Garnett T P, Segey N, Shabala P J,et al.Kinetics of ammonium and nitrate uptake by eucalypt roots and associated proton fl uxes measured using ion selective microelectrodes[J]. Functional Plant Biology, 2003, 30(11): 1165-1176.
[10] Kin T, Mills H A, Wetzstein H Y. Studies on effects of nitrogen form on growth, development, and nutrient uptake in pecan[J].Journal of Plant Nutrition, 2002, 25(3): 497-508.
[11] 许 楠, 张会慧, 朱文旭, 等. 氮素形态对饲料桑树幼苗生长和光合特性的影响[J]. 草业科学, 2012, 29(10): 1568-1572.
[12] 许 楠, 张会慧, 谷思玉, 等. 增施硝态氮对Na2CO3胁迫下桑树幼苗叶片PSⅡ功能的影响[J].草业科学,2017,34(1):67-74.
[13] 邹 娜, 李保海, 强晓敏, 等. 高铵胁迫对拟南芥根系向重性的影响及机制初探[J]. 土壤, 2013, 45(1): 166-171.
[14] 宋 扬, 张潆心, 郭 娜. 低温胁迫后不同光强对小黑杨幼苗叶片叶绿素荧光和能量分配的影响[J]. 安徽农业科学, 2013,41(10): 4421-4423.
[15] 张会慧, 张秀丽, 胡彦波, 等. 中碱钠盐胁迫对桑树幼苗生长及光合特性的影响 [J]. 南京林业大学学报(自然科学版),2013, 37(1):55-60.
[16] 张会慧, 张秀丽, 朱文旭, 等. 桑树叶片光系统Ⅱ对NaCl和Na2CO3胁迫的响应[J]. 北京林业大学学报, 2011,33(6):15-20.
[17] 原俊凤, 田长彦, 冯 固, 等. 硝态氮对盐胁迫下囊果碱蓬幼苗根系生长和耐盐性的影响[J]. 植物营养与肥料学报, 2009,15(4): 953- 959.
[18] 王常慧, 邢雪荣, 韩兴国. 草地生态系统中土壤氮素矿化影响因素的研究进展[J]. 应用生态学报, 2004, 15(11): 2184-2188.
[19] Shangguan Z P, Shao M A, Dyckmans J. Effect of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat[J]. Journal of Plant Physiology, 2000, 156(1): 46-51.
[20] Hu Y B, Sun G Y, Wang X C. Induction characteristics and response of photosynthetic quantum conversion to changes in irradiance in mulberry plants[J]. Journal of Plant Physiology,2007, 164: 959-968.
[21] Hendrickon L, Furbank R T, Chow W S. A simple alternative approach to assessing the fate of absorbed light energy using chlorophyll fl uorescence[J]. Photosynthesis Research, 2004,82: 73-81.
[22] Zhou Y H, Lam H M, Zhang J H. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice[J]. Journal of Experimental Botany, 2007, 58(5): 1207-1217.
[23] 张会慧, 张秀丽, 胡彦波, 等. 高粱-苏丹草杂交种的生长特性和光合功能研究[J]. 草地学报, 2012, 20(5): 881-887.
[24] 田纪春, 王学臣, 刘广田. 植物的光合作用与光合氮、碳代谢的耦联及调节[J]. 生命科学, 2001, 13(4): 145-147.
[25] 樊卫国, 葛慧敏, 吴素芳, 等. 不同形态氮素及其配比对铁核桃实生苗叶片光合特性和养分含量的影响[J]. 果树学报,2013, 30(3): 437-443.
[26] 陈永亮, 刘明河, 李修岭. 不同形态氮素配比对红松幼苗光合特性的影响[J]. 南京林业大学学报, 2005, 29(3): 77-80.
[27] 张福锁.土壤与植物营养研究新动态[M].北京:中国农业出版社,1993:42-75.
[28] 张一利.不同氮素形态对黄瓜和水稻光合电子传递及叶黄素循环的影响[D].杭州:浙江大学,2007.
[29] 江 力, 张荣铣. 不同氮钾水平对烤烟光合作用的影响[J]. 安徽农业大学学报, 2000, 27(4): 328-331.
[30] 谭万能, 李秧秧. 不同氮素形态对向日葵生长和光合功能的影响[J]. 西北植物学报, 2005, 25(6): 1191-1194.
[31] Raab K T, Teer Y N. Nitrogen source regulation of growth and photosynthesis inBeta vulgarisL.[J].Plant Physiology, 1994,105(1): 159-166.
[32] Pearson J R, Stewart G. The deposition of atmospheric ammonia and its effeets on plants[J].New phytologist, 1993, 125: 283-305.
[33] Robinson J M. Carbon dioxide and nitrite photo assimilatory processes do not intercompete for reducing equivalents in spinach and soybean leaf chloroplasts[J]. Plant Physiology, 1986,80: 676-684.
猜你喜欢
音乐教育与创作(2022年1期)2022-04-26 02:21:20
云南化工(2021年9期)2021-12-21 07:44:10
今日农业(2020年16期)2020-12-14 15:04:59
小学生作文(中高年级适用)(2018年3期)2018-04-18 01:24:40
学生天地(2016年16期)2016-05-17 05:46:06
影像科学与光化学(2014年5期)2014-03-11 16:03:23
无机化学学报(2014年3期)2014-02-28 17:30:42
现代检验医学杂志(2014年1期)2014-02-06 01:29:25
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
