
不同生境条件下滨海芦苇湿地C、N、P化学计量特征
2018-04-19 01:20屈凡柱付战勇孙景宽刘京涛宋爱云
生态学报 2018年5期
屈凡柱, 孟 灵, 付战勇, 孙景宽,*, 刘京涛 , 宋爱云
1 滨州学院山东省黄河三角洲生态环境重点实验室, 滨州 256600 2 山东农业大学林学院, 泰安 271018
所有的生命有机体都是由多种化学元素按照自然存在的一定比例构成。而碳(C)、氮(N)和磷(P)是按照不同比率存在于生物体中的3个主要的生源要素,它们之间的比率的动态变化决定着生物和生态系统的主要特性[1- 3]。在植物生长过程中,C是构成植物体干物质的最主要元素;N在氨基酸、蛋白质和核酸等物质的生物合成、提高植物的光合作用能力等方面起着重要作用;P是核酸和酶的重要组成部分,是生命有机体组织的基本元素。同时,植物N/P值可以作为判断植物生长对营养供给适应的指标[4]。在生态学研究领域,化学计量学通常应用于生物体的主要元素组成研究,特别是C、N和P[5]。1958年,Redfield通过研究发现海洋浮游生物体内C、N和P的原子个数比存在与大洋海水相似的固定比率,为106∶16∶1,这一值被称为Redfield比率。同时研究发现,这一比率受到环境和生物的相互调节作用。Redfield比率的实用性激发了生态学家在陆地生态系统中寻找类似的比率模式和相互关系,进而产生了一个新的学科——生态化学计量学,试图用它来解释生态相互作用的多种化学元素平衡关系[6]。生态化学计量学已经被证明在理解营养物质之间相互作用和养分循环方面有重要价值;同时,生态化学计量理论结合热力学第一定律,生物进化的自然选择、分子生物学的中心法则[7-8],已经在不同尺度上,从分子到生物,从生态系统到生物圈进行了各种研究[9-11]。
我国关于生态化学计量学的系统研究起步较晚,2004年“Ecology”杂志和2005年“Oikos”杂志先后出版了生态化学计量学特刊或专题,集中报导了生态化学计量学这一生态学研究热点,引起了国内学者的广泛关注。随后曾德慧和陈广生以“生态化学计量学:复杂生命系统奥秘的探索”为题对生态化学计量学的概念、历史起源、基本理论、研究进展和应用前景进行了综述[12]。任书杰等对中国东部南北样带654种植物进行研究,发现叶片中N和P含量与纬度、年均温度间存在极显著的相关关系,随着纬度升高和年均温度的降低,叶片中N和P含量极显著地增加(P<0.001),N∶P与纬度和年均温度的相关性较弱(P=0.386和P=0.342)[13],与全球尺度的研究结果比较发现我国的陆地植物N∶P明显偏低,这说明我国陆地植物相对于其他国家更缺P。贺金生等研究了内蒙古、新疆和青藏高原地区的213种植物,发现3个地区的植物叶片的C∶N没有差异,生活型和种类的不同导致70%的变异,温度和降水只解释了3%的变异[14]。庾强等在内蒙古锡林郭勒草原生态系统国家野外科学观测研究站进行了两年的NP添加实验,同时结合一个1200km的样带实验和草原站27年的长期监测数据,从时间和空间尺度充分证明了内稳性高的物种具有较高的优势度和稳定性,内稳性高的生态系统具有较高的生产力和稳定性;研究结果表明,通过调节生物对环境因子的响应,化学计量内稳性成为生态系统结构、功能和稳定性维持的重要机理[15]。2010年《植物生态学报》集中报导了国内生态化学计量学方面的研究,展示了近年来中国生态化学计量学的研究进展。但总体上,我国的生态化学计量学研究主要集中于陆地生态系统。从生态化学计量学角度探讨滨海湿地植物群落演变机理,评价河口滨岸土壤养分限制状况及其对生态系统潜在的环境效应的相关研究鲜见报道。
芦苇(Phragmitesaustralis),在近30年得到了广泛的关注,它作为一种适应性广、抗逆性强、生物量高的禾本科植物,在世界范围内广泛分布。在适宜的条件下,单株芦苇可以在两年内繁殖覆盖0.05hm2的土壤和水面,拥有极强的繁殖能力、对其他植物光线的遮盖能力和产生大量腐殖质的能力。其适宜生长的地区为热带和温带地区的低海拔湿地或浅水,黄河三角洲滨海湿地属于此类地区。黄河三角洲滨海湿地处在陆地-海洋相互作用活跃的地带,是世界上陆地面积增长最快的地区,也是我国暖温带地区最年轻、最广阔的湿地生态系统[16- 17]。黄河三角洲国家级自然保护区内芦苇湿地集中分布面积达2700hm2,20世纪,区外大面积芦苇湿地曾被开垦成棉田,为响应退耕还湿工程,退耕芦苇湿地也大面积存在[18]。目前,我们尚未发现任何关于生境条件对湿地土壤和植物C、N、P化学计量学特征影响的相关研究,本研究选择新生湿地和退耕湿地作为研究对象,旨在为正在进行的黄河三角洲湿地保护和恢复工作提供借鉴。
1 材料与方法
1.1 研究区概况
研究区位于黄河三角洲国家级自然保护区(图1)入海口和中国科学院黄河三角洲滨海湿地生态试验站内。黄河三角洲国家级自然保护区于1992年经国务院批准建立,是东北亚和环西太平洋鸟类的越冬栖息地和中转站以及典型新生河口湿地。历史上黄河经历多次改道,现入海口为1996年改道形成。入海口处新增土地面积约1300hm2/a,属于典型的新生滨海湿地。中国科学院黄河三角洲滨海湿地生态试验站位于自然保护区大汶流管理站西侧,成立于2011年5月,试验站占地约40hm2,站内土地原为棉田,为了生态试验站建设于2009年开展了退耕还湿工作,站内湿地均属于退耕湿地。两个研究区均有芦苇优势种群的自然植被覆盖区。研究区属温带大陆性季风气候,年降水量为551.6mm,绝大部分降水集中于7—8月,多形成季节性淹水。年平均气温为12.1℃,无霜期为196d,土壤为隐域性潮土和盐土。

图1 黄河三角洲地理位置图和采样点分布Fig.1 Location of the sampling sites in the coastal wetland in the Yellow River Delta
1.2 样品采集
2015年9月17日,在黄河三角洲国家级自然保护区与中国科学院黄河三角洲滨海湿地生态试验站内分别选择3个30m×30m群落高度相近芦苇样地,每个样地内采取蛇形取样,选取7—10个50cm×50cm芦苇植物生物量大致相当的小样方,芦苇地上部分采用收割法进行采集,清除地表枯落物,用剪刀沿地面剪下植物的地上部分带回实验室,实验室内摘除枯死物。地下部分采样时采用挖掘法,将对应的50cm×50cm样方范围的根全部挖出,带回实验室,用水冲洗至无泥,分拣活根、腐根。地上、地下植物体活体分别于105℃杀青2h,80℃下烘干至恒重。采集芦苇植物样品的同时采集深度为50cm的土壤样品,每层10cm。将土壤样品放入聚乙烯袋,带回实验室自然风干,捡出植物残体,研磨过100目土壤筛后装袋待测。
1.3 样品分析与数据处理
土壤pH值和电导率(EC)测定按照土样:去离子水(水土比=5∶1)浸提,分别用pH值和电导率仪进行测定。盐度测定采用差量法。土壤和植物TC、TN采用大进样量元素分析仪(Vario MACRO cube,德国Elmentar)进行分析测定,TP的测定按照常规农化分析手册采用钼蓝比色法测定。所有样品的测定均在滨州学院-山东省黄河三角洲生态环境重点实验室内进行。数据的统计和比较分析采用SAS v8.1(SAS Inc., Cary, NC)软件,采用单因方差分析(One-way ANVOA)比较不同类型芦苇湿地pH、EC、盐度和养分含量比值差异是否显著。并采用Duncan法进行多重比较,以T检验,判断测定指标差异是否显著。采用Pearson相关分析,检验土壤植物TC、TN和TP含量及比值关系以及三者之间可能存在的关系。用SPSS 17.0软件对所得实验数据进行统计分析,表中数据均采用平均值±标准差(SD)。实验数据作图采用OriginPro 8.5 软件。
2 结果
2.1 滨海芦苇湿地土壤和植物C、N、P的含量与相关性
黄河三角洲滨海芦苇湿地土壤基本理化性质见表1。研究区域内新生芦苇湿地和退耕芦苇湿地的土壤均为碱性土壤。退耕湿地土壤含盐量的平均值为12.7‰,显著高于新生湿地土壤含量6.1‰。退耕湿地土壤TC和TN平均含量分别为14.7g/kg和485.8mg/kg,显著高于新生湿地8.7g/kg和385.2mg/kg,二者差异水平显著。但退耕湿地TP平均含量略高于新生湿地,分别为531.5mg/kg和523.9mg/kg,二者不存在显著差异,为进行生境条件对滨海芦苇湿地土壤C、N、P的化学计量特征影响的研究提供了可行性。通过对两种类型滨海湿地的土壤和植物TC、TN和TP的相关分析可以发现,新生湿地和退耕湿地样地的土壤TC、TN与TP存在相关关系,其中新生湿地TN和TP,退耕湿地TC、TN和TP显著相关(表2)。退耕湿地植物体TC、TN和TP三者相关性显著;但新生湿地植物样本的相关性不显著。

表1 黄河三角洲滨海芦苇湿地土壤基本理化性质
EC:电导率,electrical conductivity; SSC:土壤含盐量,soil salt content; TC:总碳,total carbon; TN:总氮,total nitrogen; TP:总磷,total phosphorus
表2芦苇湿地土壤(n=20, 25)和植物体(n=12, 16)C、N、P含量的相关分析
Table2CorrelationmatrixofTC,TNandTPinreed-dominatedwetlandssoils(n=20, 25)andplants(n=12, 16)intheYellowRiverDelta
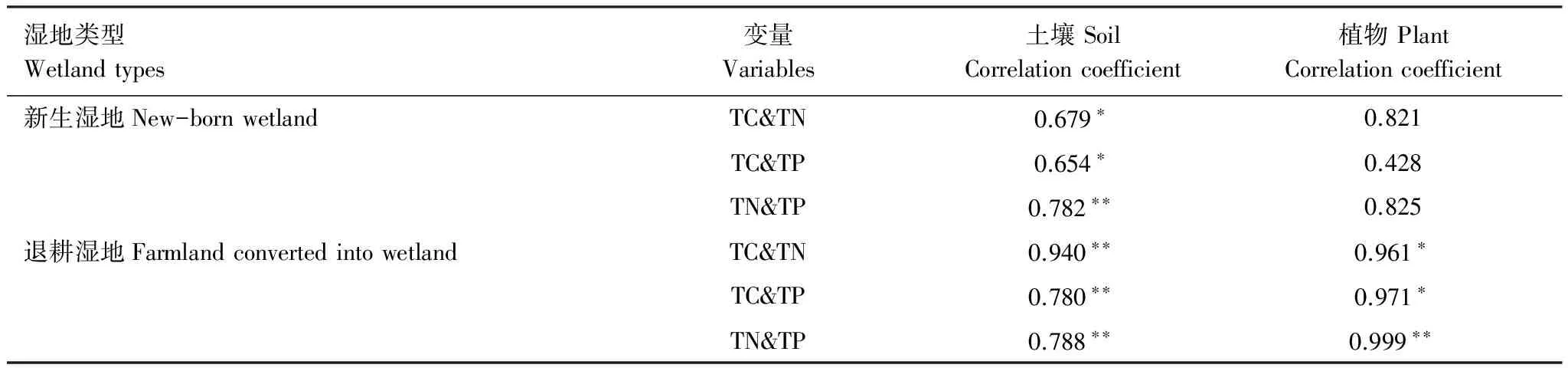
湿地类型Wetlandtypes变量Variables土壤SoilCorrelationcoefficient植物PlantCorrelationcoefficient新生湿地New-bornwetlandTC&TN0.679∗0.821TC&TP0.654∗0.428TN&TP0.782∗∗0.825退耕湿地FarmlandconvertedintowetlandTC&TN0.940∗∗0.961∗TC&TP0.780∗∗0.971∗TN&TP0.788∗∗0.999∗∗
n: 样本数; *P<0.05; **P<0.01
2.2 滨海芦苇湿地土壤C、N、P的化学计量值对比
黄河三角洲滨海芦苇湿地土壤RCN、RCP和RNP的剖面分布如图2。在新生湿地土壤剖面中,RCN和RCP变化剧烈,RCN、RCP最小值出现在20—30cm土层,呈“V”型分布。RNP值表层最大,随深度的增加而减小。在退耕湿地土壤剖面中,RCN值变化剧烈且随深度的增加而变大;RCP和RNP值的剖面分布特征一致,随深度的增加而减小,且底层30—40cm和40—50cm土层变化不明显。
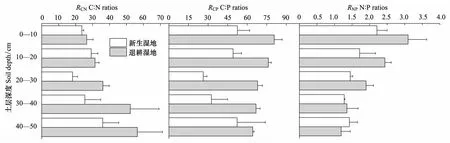
图2 新生湿地和退耕湿地土壤RCN,RCP和RNP的对比Fig.2 The distribution of RCN, RCP and RNP in soil profile in the new-born wetland and in the farmland converted into wetland
新生湿地和退耕湿地土壤RCNP分别为42.6∶1.6∶1、71.2∶2.0∶1。RCN、RCP和RNP的平均值分别为:26.5、42.6、1.6;40.5、71.2、2.0(表3)。退耕湿地土壤RCN、RCP和RNP值均高于新生湿地。两种湿地类型RCN、RCP和RNP之间具有显著性差异(RCN:F=3.75,P=0.047;RCP:F=4.24,P=0.007;RNP:F=3.97,P=0.039)。
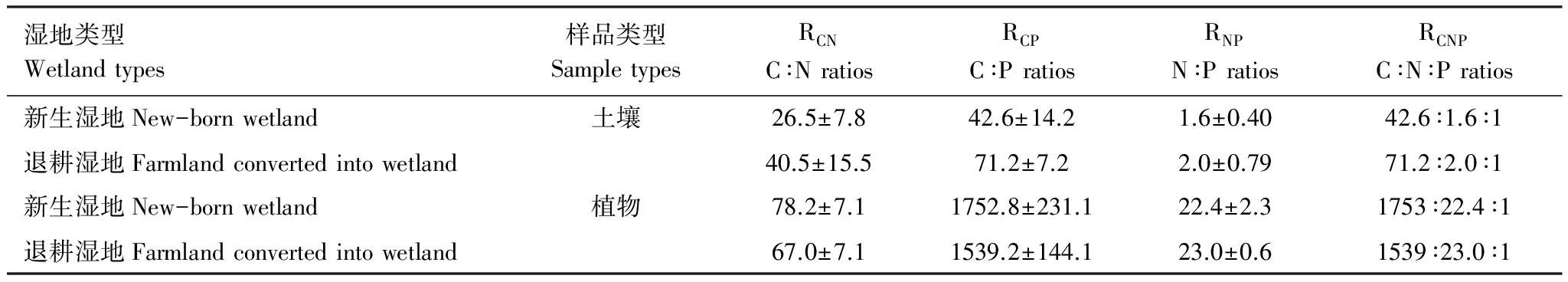
表3 黄河三角洲滨海芦苇湿地土壤和植物体C、N、P化学计量值对比
RCN、RCP、RNP和RCNP所代表的C∶N比值、C∶P比值、N∶P比值和C∶N∶P比值均为元素的摩尔质量比
2.3 滨海芦苇湿地植物C、N、P的化学计量值对比
图3为黄河三角洲滨海芦苇植物RCN、RCP和RNP的对比图。两种湿地芦苇地上部分RCN值变化不大,新生湿地芦苇地下部分RCN值高于退耕湿地,两者之间的差异性不明显(P=0.149)。退耕湿地芦苇地上部分RCP和RNP值高于新生湿地,二者的差异性均不显著(P=0.193;P=0.315);新生湿地芦苇地下部分RCP和RNP低于退耕湿地,二者RCP、RNP值之间差异显著(P=0.021;P=0.008)。新生湿地和退耕湿地芦苇植物整株RCN、RCP和RNP平均值分别为78.2、1752.8、22.4;67.0、1539.2、23.0(表3)。虽然新生湿地芦苇植物整株RCN和RCP值均稍高于退耕湿地,但二者差异不显著。
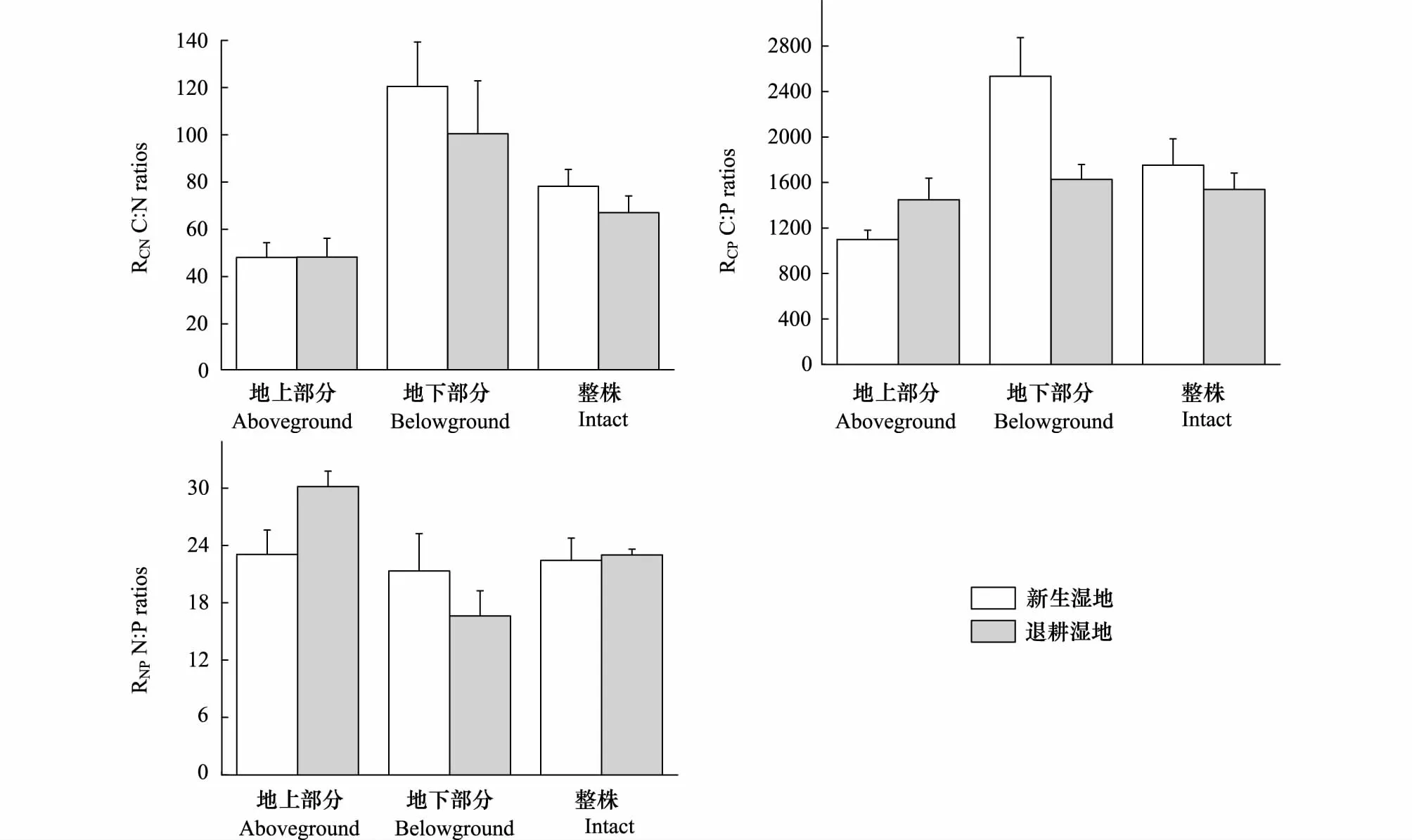
图3 新生湿地和退耕湿地中芦苇植物体R CN , RCP 和RNP 的对比Fig.3 The RCN, RCP and RNP in plant tissue of the new-born wetland and the farmland converted into wetland
3 讨论
3.1 不同尺度生态系统C、N、P的化学计量特征
从Redfield比率开始提出,化学计量特征的研究逐渐广泛应用于海洋和淡水浮游植物研究;同时,也为陆地生态系统的相关研究开启了一扇大门。2007年,Cleveland和Liptzin[10]研究发现土壤和土壤中微生物群落的C∶N∶P值与海洋浮游植物类似,虽然在全球范围内土壤中微生物群落的个体亲缘组浓度有所不同,但微生物群落的C∶N∶P值的平均值被较好地约束在一个范围内,土壤和土壤中微生物的C∶N∶P值分别为186∶13∶1和60∶7∶1(表4)。Tian等[20]研究发现,中国范围内土壤C∶N∶P值为60∶5∶1;由于土壤表层0—10cm有机质含量丰富,有机体与环境相互作用活跃,C∶N∶P值约为134∶9∶1。同时,相关研究发现表明:在陆地生态系统中元素的化学计量特征较海洋变化剧烈且不固定,但固定的比率很有可能存在于与海洋相似的营养丰富、环境相似的植物群落或森林生态系统中[21- 23]。
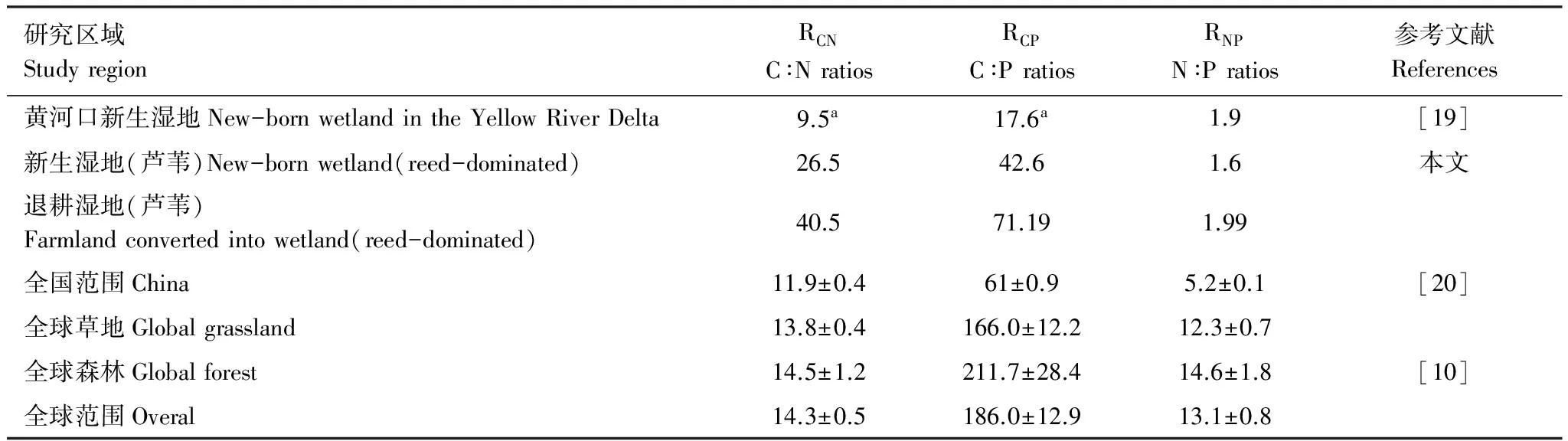
表4 不同尺度土壤C、N、P化学计量特征的对比
a为RCN和RCP比值计算中使用的是有机碳
3.2 生境对土壤C、N、P的化学计量特征的影响
在陆地和海洋生态系统中,C和N的生物地球化学循环是紧密耦合的[5,10]。影响土壤C∶N∶P值的因素较多,主要受区域水热条件和成土作用的控制。由于气候、地貌、成土母质、年代、植被和土壤生物等土壤形成因子和人类活动的影响,土壤C、N和P总量变化很大,也使得土壤C∶N∶P值的空间变异性较大。对若尔盖湿地沼泽土、草甸化沼泽土和草甸土研究发现,土壤C∶N值分别为40.38,31.70,23.26,C∶P值分别为409.52,247.46,113.07,N∶P值分别为10.43,7.90,5.02,土壤C/N、C/P、N/P均随湿地退化而减小,较高的C/P与N/P<14揭示氮磷元素均是影响植物生长的限制性因素,且受氮素限制高于磷素[24]。曾全超等通过对黄土高原陕北地区不同纬度下5个典型植被区的34个典型土样的养分及生态化学计量指标研究发现,随着植被的恢复,相对于高纬度地区,低纬度地区更容易缺磷;高纬度地区的植被更容易受到N含量的限制[25]。刘兴华等对黄河三角洲未利用地开发过程中的农耕区、未利用地以及二者之间的过渡区土壤与优势物种碱蓬和芦苇的碳、氮、磷化学计量特征进行研究发现,未利用地开发利用过程中,土壤的C∶N和C∶P值范围分别为:7.11—20.96和3.16—21.04,N∶P值范围在0.44—1.66之间[26]。本研究中退耕芦苇湿地TC和RCN显著高于新生滨海芦苇湿地,表明农业开垦可以显著提高滨海湿地土壤有机物的含量。同时,黄河三角洲滨海芦苇湿地土壤中C∶N值(RCN=26.5—40.5)显著高于全球范围和全国范围内C∶N值的平均值,N∶P比值(RNP=1.61—1.99)显著低于全球范围(13.1)和全国范围内N∶P值的平均值(5.2),可以发现黄河三角洲滨海芦苇湿地土壤处于N限制状态。大量关于土壤固定过程C∶N比值研究表明,植物是陆地生态系统土壤C、N的主要来源,而P含量主要取决于土壤母质和岩石风化。本研究结果表明黄河三角洲芦苇湿地土壤中C∶N∶P值并不存在与Redfield比率相似的比值,可以看出滨海湿地生态系统的养分变化剧烈及其脆弱性。
3.3 生境对植物C、N、P的化学计量特征的影响
植物的元素含量不是一个固定值,每月、每天甚至每小时以及不同的植物不同的部分之间都会变化[27]。植物C、N、P化学计量特征受到经纬度、海拔、年均温度和年降水等环境因子的影响。姜沛沛等对陕西省刺槐、辽东栎和油松林3种乔木研究发现,不同树种乔木N、P含量与土壤N、P含量均没有显著相关性,均大致表现出随着年均温度和年降水的增加而增加,随着经纬度的增加而降低的趋势[28]。相关研究表明:在中国东部南北样带森林生态系统中,优势植物叶片C∶N∶P质量比为313.9∶11.5∶1,摩尔比为810.9∶25.4∶1。与全球尺度的研究结果相比,叶片C/N明显偏高,N/P明显偏低,C/P差异不显著[29]。在荒漠极端环境下,土壤表现出严重的N缺乏,植物叶片N、P含量及N∶P值之间均有显著差异[30]。罗艳等对塔里木盆地南缘克里雅河流域旱生芦苇生态化学计量特征进行研究发现,芦苇的生长主要受N限制,同时组织器官对C、N以及C∶N和N∶P的影响最大,生境对P及C∶P的影响最大[31]。土地利用方式的改变打破森林生态系统原有的碳氮磷平衡,从而显著地影响森林生态系统的生物地球化学循环过程。周正虎等研究发现土地利用变化显著改变土壤碳-氮-磷的耦合关系,不同利用方式的东北温带幼龄林土壤碳-氮(C/N)之间存在极显著(P<0.001)的线性关系[32],结果与滨海湿地芦苇湿地土壤相似。本研究新生湿地和退耕湿地中芦苇植株C∶N∶P值分别为1753∶22.4∶1和1539∶23.0∶1。虽然C∶N∶P值没有限制在一定范围,但两种湿地类型芦苇植物体的N∶P值被较好的约束在23左右,造成这种情况原因很大可能由土壤N限制及P来源的稳定性导致。
4 结论
在农业开垦的影响下,芦苇湿地土壤TC、TN的含量明显增加,TP的含量变化不大。新生湿地和退耕湿地土壤RCNP分别为42.6∶1.6∶1、71.2∶2.0∶1,RNP远低于全球平均水平(13.1)和我国平均水平(5.2),黄河三角洲滨海芦苇湿地土壤表现为N限制。新生湿地土壤剖面中,RCN和RCP变化剧烈,RCN、RCP值最小值出现在20—30cm土层,呈“V”型分布;RNP值表层最大,随深度的增加而减小。而退耕还湿后湿地土壤RCN值规律性较好,RCN随深度的增加而变大,RCP和RNP值随深度的增加而减小。新生湿地和退耕湿地中芦苇植物整株RCN、RCP和RNP平均值分别为78.2、1752.8、22.4;67.0、1539、23.0。开垦活动可以降低芦苇植物体RCN和RCP值,但对RNP值的影响不大,芦苇植物体本身对RNP的约束性较高,且RNP约为23.0。本研究表明,开垦活动可以使土壤剖面C、N、P的化学计量特征的变化显得有序,而植物体的C、N、P化学计量特征却受土壤养分、开垦活动和植物本身固定作用的多重制约。
参考文献(References):
[1] Sterner R W, Elser J J. Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere. Princetion, NJ: Princeton University Press, 2002.
[2] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论. 植物生态学报, 2010, 34(1): 2- 6.
[3] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937- 3947.
[4] 王维奇, 徐玲琳, 曾从盛, 仝川, 张林海. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征. 生态学报, 2011, 31(23): 7119- 7124.
[5] Michaels A F. The ratios of life. Science, 2003, 300(5621): 906- 907.
[6] Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L J. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540- 550.
[7] Elser J J, Sterner R W, Galford A E, Chrzanowski T H, Findlay D L, Mills K H, Paterson M J, Stainton M P, Schindler D W. Pelagic C: N: P stoichiometry in a eutrophied lake: responses to a whole-lake food-web manipulation. Ecosystems, 2000, 3(3): 293- 307.
[8] Zhang Z S, Song X L, Lu X G, Xue Z S. Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: influences of vegetation coverage, plant communities, geomorphology, and seawalls. Journal of Soils and Sediments, 2013, 13(6): 1043- 1051.
[9] Sardans J, Rivas-Ubach A, Peuelas J. The C: N: P stoichiometry of organisms and ecosystems in a changing world: a review and perspectives. Perspectives in Plant Ecology, Evolution and Systematics, 2012, 14(1): 33- 47.
[10] Cleveland C C, Liptzin D. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 2007, 85(3): 235- 252.
[11] Hessen D O, Elser J J, Sterner R W, Urabe J. Ecological stoichiometry: an elementary approach using basic principles. Limnology and Oceanography, 2013, 58(6): 2219- 2236.
[12] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007- 1019.
[13] 任书杰, 于贵瑞, 陶波, 王绍强. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 2007, 28(12): 2665- 2673.
[14] He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Geng Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(1): 115- 122.
[15] Yu Q, Chen Q S, Elser J J, He N P, Wu H H, Zhang G M, Wu J G, Bai Y F, Han X G. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecology Letters, 2010, 13(11): 1390- 1399.
[16] 于君宝, 王永丽, 董洪芳, 王雪宏, 栗云召, 周迪, 高永军. 基于景观格局的现代黄河三角洲滨海湿地土壤有机碳储量估算. 湿地科学, 2013, 11(1): 1- 6.
[17] Yu JB, Fu YQ, Li YZ, Han GX, Wang YL, Zhou D, Sun W D, Gao Y J, Meixner F X. Effects of water discharge and sediment load on evolution of modern Yellow River Delta, China, over the period from 1976 to 2009. Biogeosciences, 2011, 8(9): 2427- 2435.
[18] Qu F Z, Yu J B, Du S Y, Li Y Z, Lv X F, Ning K, Wu H F, Meng L. Influences of anthropogenic cultivation on C, N and P stoichiometry of reed-dominated coastal wetlands in the Yellow River Delta. Geoderma, 2014, 235- 236: 227- 232.
[19] 于君宝, 陈小兵, 孙志高, 谢文军, 毛培利, 吴春发, 董洪芳, 牟晓杰, 栗云召, 管博, 单凯. 黄河三角洲新生滨海湿地土壤营养元素空间分布特征. 环境科学学报, 30(4): 855- 861.
[20] Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C: N: P ratios in China′s soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139- 151.
[21] Hedin L O. Global organization of terrestrial plant-nutrient interactions. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 10849- 10850.
[22] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001- 11006.
[23] McGroddy M E, Daufresne T, Hedin L O. Scaling of C: N: P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology, 2004, 85(9): 2390- 2401.
[24] 叶春, 蒲玉琳, 张世熔, 王贵胤, 王奥博, 王丹, 贾永霞, 徐小逊. 湿地退化条件下土壤碳氮磷储量与生态化学计量变化特征. 水土保持学报, 2016, 30(6): 181- 187, 192.
[25] 曾全超, 李鑫, 董扬红, 李娅芸, 程曼, 安韶山. 陕北黄土高原土壤性质及其生态化学计量的纬度变化特征. 自然资源学报, 2015, 30(5): 870- 879.
[26] 刘兴华, 陈为峰, 段存国, 王凯, 王海翠. 黄河三角洲未利用地开发对植物与土壤碳、氮、磷化学计量特征的影响. 水土保持学报, 2013, 27(2): 204- 208.
[27] Kalra Y P. Handbook of Reference Methods for Plant Analysis. Boca Raton, FL: CRC Press, 1998.
[28] 姜沛沛, 曹扬, 陈云明, 赵一娉. 陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征. 生态学报, 2017, 37(2): 443- 454.
[29] 任书杰, 于贵瑞, 姜春明, 方华军, 孙晓敏. 中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征. 应用生态学报, 2012, 23(3): 581- 586.
[30] 陶冶, 张元明. 古尔班通古特沙漠4种草本植物叶片与土壤的化学计量特征. 应用生态学报, 2015, 26(3): 659- 665.
[31] 罗艳, 贡璐. 塔里木盆地南缘不同生境下芦苇生态化学计量特征. 生态学杂志, 2016, 35(3): 684- 691.
[32] 周正虎, 王传宽, 张全智. 土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响. 生态学报, 2015, 35(20): 6694- 6702.
猜你喜欢
世界科学技术-中医药现代化(2021年5期)2021-11-05
世界科学技术-中医药现代化(2021年12期)2021-04-19
华人时刊(2020年21期)2021-01-14
现代装饰(2019年12期)2020-01-19
建材发展导向(2019年11期)2019-08-24
宝藏(2018年12期)2019-01-29
戏曲研究(2018年3期)2018-03-19
创新科技(2015年9期)2015-12-15
华东经济管理(2015年7期)2015-11-14
太空探索(2015年2期)2015-07-12
