
我国地表臭氧生态环境效应研究进展
2018-04-19 01:20冯兆忠袁相洋姜立军代碌碌
生态学报 2018年5期
冯兆忠,李 品,袁相洋,高 峰,姜立军,代碌碌
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085 2 中国科学院大学资源环境学院,北京 101408
地表臭氧(O3)特指距离地球表面15km范围内的对流层O3,是地表大气光化学烟雾及温室气体的主要成分,具有强氧化性,能严重危害地表农作物和自然植物生长,被称为“坏O3”[1]。地表O3除少量来自平流层的大气传输过程(动力下传)[2]外,绝大部分来自大气中甲烷(CH4)、一氧化碳(CO)、氮氧化物(NOx=NO+NO2)及挥发性有机化合物(VOCs)等一次污染物在强烈紫外辐射下发生复杂光化学反应生成的产物。依据前体物的不同,O3的形成主要通过两个过程同时进行:① NO2在强烈光照射下直接发生光解反应,释放出游离氧原子(O·),不稳定的O·和空气中的氧分子(O2)结合生成O3;② 空气中的O2光解产生的自由基可将VOCs等前体物氧化为过氧化物自由基(RO2·)和羟基自由基(OH·),这些活性自由基进一步促进大气中NO向NO2转化,从而提供O3形成的NO2源[3]。
地表O3的生成受气象条件(光照、大气温湿度及风速等)、排放源分布和土地利用类型等影响,其浓度高峰值一般出现在前体物浓度较高的夏季午后[4]。其光化学形成、区域传输、还原氧化及沉降分解等过程共同决定着某一地区的地表O3浓度状况。基于经验动力学模型(Empirical Kinetics Modeling Approach,EKMA)和已有的观测数据显示,O3浓度与其前体物浓度(尤其是NOx与VOCs)呈显著的非线性响应关系[5-6]。从NOx浓度较高的中心城区到VOCs浓度较高的近郊远郊地区,存在由市中心的VOCs控制逐渐转变为近郊的VOCs和NOx共同控制,进而过渡到远郊及乡村的NOx控制为主的移动特征[7]。地表O3形成后一方面随大气环流进行跨区域、长距离的传输,另一方面与大气污染物的氧化产物OH·及HO2·等发生反应还原为O2,或直接向地表沉降并最终分解。
1 我国地表臭氧污染水平
20世纪90年代,中国大部分地区地表O3浓度显著低于美国、欧洲等发达国家[8]。但是,随着我国工业化和城市化进程的加快,O3前体物尤其是NOx排放量显著增高,导致国内大部分地区地表O3浓度显著升高[9-11]。据统计数据显示,在过去20年间我国NOx排放总量以每年5%的速度递增,截止到2010年已到达21.9—26.1Tg[12-14];并且排放量高的地区大部分集中在东部及东南沿海经济发达、人口稠密的华北平原、长江三角洲及珠江三角洲等地[3]。中国环保部发布的2014—2015年《中国环境公告》显示,2013年到2015年间全国74个城市的白天8h平均O3浓度从69.5nmol/mol增加到75nmol/mol。高浓度的地表O3已经成为中国大多数城市夏季最主要的空气污染物,尤其北京、上海、济南、杭州、香港和广州等发达城市[3,12]。从2015年全国1497个空气质量监测站点数据分析结果(图1)来看,我国年平均地表O3浓度从南到北呈现出明显地域差异,4—9月生长季白天12h(8:00—20:00)的O3浓度平均值(M12)已超过40nmol/mol,最高可达70nmol/mol。M12超过50nmol/mol的区域主要集中在东北平原南部、华北平原、长江三角洲地区及中南部分地区,西南及西北等低纬度地区的O3浓度相对较低(图1)。此外,O3浓度还表现出明显的季节波动特征,在中国北部及中部地区,高浓度O3一般出现在夏季;但在南部地区,其高值通常出现在秋季,夏季呈现略低的趋势[12]。基于月平均值的观测数据显示,长江三角洲地区的地表O3浓度高值出现在5月,华北平原地区出现在7月,珠江三角洲地区出现在10月[15]。而且,受到大气传输过程的影响,处于下风向地区的O3浓度往往偏高,比如2010年北京地区夏季白天小时平均O3浓度,处于下风向的郊区(67±27)nmol/mol比城区高近20nmol/mol[16]。因此,综合大气化学模型模拟和实际O3浓度监测数据的结果,都表明目前中国大部分地区出现高浓度的地表O3污染,并且主要集中在夏季。中国必将成为全球高浓度O3污染危害与研究的热点区域[17]。
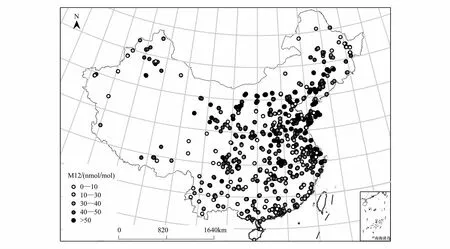
图1 中国1497个监测站点2015年4—9月生长季12小时(8:00—20:00)O3浓度平均值示意图(M12)Fig.1 The 12-hour (08:00—20:00) mean O3 concentration index (M12) of 2015 (April—September) from 1497 monitoring stations in China
2 地表臭氧对植物的影响机制
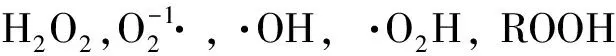
其次,进入细胞的大量O3将诱导抗氧化系统启动解毒和修复的非气孔防御响应[18,21]。具体来说,抗氧化系统中的质外体抗氧化剂(主要是抗坏血酸(AsA))首先进行初级解毒防御,将ROS部分降解;过剩的ROS进入共质体后,启动次级解毒响应[22-24],大量的抗氧化剂(AsA,酚类化合物,谷胱甘肽(GSH)等)和抗氧化酶类(超氧化物歧化酶(SOD), 过氧化物酶(POD),过氧化氢酶(CAT)及抗坏血酸过氧化物酶(APX)等)被诱导生成,参与诸如抗坏血酸-谷胱甘肽(AsA-GSH)循环等一些氧化还原过程,从而对ROS进行解毒。一旦ROS的累积量超过了初级和次级解毒防御响应的阈值,整个防御体系将崩溃,促发细胞程序性死亡(programmed cell death, PCD),表现出来的特征为叶片出现可见伤害症状(如斑点、坏死斑块、早衰、提前脱落)等。
另外,气孔防御启动时关闭的气孔阻挡了CO2吸收和水汽交换,从而抑制光合和蒸腾作用;在抗氧化系统解毒过程中将消耗大量的能量并对光合反应系统造成不同程度的损伤,导致羧化速率和电子传递速率的降低,影响光合速率和代谢反应[25- 26],最终导致植物生长缓慢,生物量降低。
3 地表臭氧对植物叶片的表观影响
暴露在O3污染下的敏感性植物(如田间作物和绿化树木)通常会出现叶片可见伤害症状,典型的症状表现为叶片上表面的叶脉之间均匀地散布着形状、大小规则的细密点状缺绿斑,呈黄/红褐色或棕色,老叶比新叶症状严重,叶片的叶脉以及下表面正常,无明显虫害和霉斑;随着受害时间的延长,有的坏死斑点会增大并连成大的斑块,导致叶片呈现黑褐色灼伤状条状干枯斑纹[1,27-28]。
可见伤害症状的评估列入了欧洲林业组织如ICP Forests和ICP Vegetation[29- 31]和北美的一些森林健康监测项目中[32],用于指示当前环境O3浓度是否对植物造成胁迫及伤害程度[31]、筛选本地O3胁迫指示物种和评估城市及区域的O3风险[33]。尽管我国许多地区的空气O3浓度已经超过了敏感植物受害的临界浓度阈值(40nmol/mol),但开展野外调查O3对植物胁迫伤害症状的研究非常有限,仅见近两三年在北京及附近区域的数篇报道[16,34-36]。本文作者联合欧洲科学家分别在2013和2014年的植物生长季期间,在北京及其周边地区(河北、天津等地)调查发现至少有28种植物(包括树木和农作物)叶片呈现出典型的O3受害症状[35]。由于O3的累积效应,O3伤害症状多出现在植物生长旺季末期8月底至9月初。对于中国温带地区而言,臭椿、木槿和豆类作物可作为表征地表O3污染的关键指示物种[35]。
4 地表臭氧对植物光合固碳能力的影响
短期高浓度的O3暴露会造成叶片出现明显可见伤害症状[37-38],而长期低浓度的O3暴露则会影响植物的光合作用、生长发育、产量/生物量以及果实、籽粒的品质[25- 26,39]。环境中地表O3浓度具有明显日间波动和季节性波动特征,急性、慢性损害可在同一植物的不同生长时期出现。并且,植物在O3暴露下可能不出现可见症状但生长发育受阻、早衰和减产/生物量降低。下面就O3对我国主要农作物和森林树木光合和固碳能力的影响分别从光合作用、农作物产量和品质、以及树木生长和生物量三方面进行详述。
4.1 光合作用
我国利用开顶式气室(OTCs)进行O3熏蒸实验,证实O3显著降低了树木的净光合速率(Asat)、气孔导度(gs)和叶绿素含量,并使光合电子传递链受阻。供试植物包括一些温带落叶阔叶树如银杏(Ginkgobiloba)、蒙古栎(Quercusmongolica)、臭椿(Ailanthusaltissima)和法桐(Platanusorientalis)等[40-41];亚热带落叶针叶树水杉(Metasequoiaglyptostroboides)[42]和落叶阔叶树鹅掌楸(Liriodendronchinense)、枫香(Liquidambarformosana)[37,43]等,以及亚热带常绿阔叶树香樟(Cinnamomumcamphora)和青冈(Cyclobalanopsisglauca)[37,44]等。
O3对植物光合作用的影响因物种、品种或基因型和生长阶段(幼苗或成树)而异。Meta分析结果显示,当O3浓度平均升高到73nmol/mol和62nmol/mol时,分别能使小麦和水稻的光合速率降低20%和28%[25,45]。Pang等[46]研究发现,高浓度O3引起敏感性水稻品种SY63和抗性品种WYJ3旗叶Asat分别降低23.1%和9.4%,并且SY63比WYJ3的响应显著提前29d,叶绿素荧光参数(PSII实际光量子效率(ФPSII)和光化学猝灭系数(qP))也显示出相同的趋势。O3对小麦品种Yangmai16(Y16)光合作用的影响仅在籽粒灌浆的后期表现出显著差异[47]。高浓度O3显著降低了油松(Pinustabulaeformis)上一年生枝条叶的Asat和ФPSII,但对当年生枝条叶影响不显著[48]。
O3对植物光合作用的影响也因其他环境因子的变化而不同。在O3和CO2浓度同时升高的情况下,邵在胜等[49]研究表明水稻叶片光合速率的改变与对照相比并无显著差异;然而Yan等[50]发现CO2浓度升高缓解了O3对蒙古栎(Quercusmongolica)幼苗光合速率降低的负效应。当加入干旱胁迫后,干旱加重了O3对元宝枫(Acertruncatum)叶片气体交换的影响,两者交互作用对Asat和gs的降低幅度较O3单因子胁迫分别增加23.7%和15.5%[51]。在氮添加(即增加有效氮含量)的条件下,O3对小麦和水稻叶片光合速率和叶绿素含量的负效应有所缓解[52-53];然而添加氮肥(60 kg hm-2a-1)并没有改变O3对香樟(Cinnamomumcamphora)叶片Asat、羧化作用、PSII量子产量和光合色素含量的降低程度[44]。
4.2 农作物产量与品质
相比于国外,我国在O3污染对作物影响方面的研究起步较晚,但发展迅速。从20世纪90年代末开始,利用OTCs、开放式O3浓度增加(Free Air O3Concentration Enrichment, O3-FACE)系统和田间原位施用化学防护剂,研究了我国主要粮食作物和蔬菜对O3浓度升高的响应,包括冬小麦[47,52,54- 56]、水稻[46,49,53, 57- 59]、大豆[60-63]、玉米[64]、油菜[65- 67]和菠菜[68- 69]等。
暴露在O3污染下的农作物均出现不同程度的减产[59,70-71],这在浙江嘉兴和江苏江都地区分别利用OTC和O3-FACE对水稻和冬小麦长达5年的研究中得到证实。O3浓度增加使得4种水稻(杂交品种WJ15和YD6,自交品种SY63和LYPJ)的平均产量降低12%,其中杂交品种WJ15和YD6分别降低17.5%和15%,但对自交品种产量损失影响不显著[59]。O3浓度在当前环境浓度的基础上增加25%,可使冬小麦籽粒重显著降低,从而引起20%的产量损失[71]。佟磊等[72]利用O3暴露量AOT40(小时O3浓度超过40nmol/mol的累积值)与产量的响应关系,对我国从北到南5个地区(北京、定兴、江都、嘉兴、东莞)的水稻和冬小麦的O3敏感性进行了比较分析,结果表明水稻对O3的敏感性从北到南呈逐渐增加的趋势,但冬小麦对O3的敏感性并无明显的地域变化规律。张巍巍等[62]通过对东北地区广泛种植的9种大豆品种的研究发现,O3浓度升高使得大豆平均产量降低40%,其中O3对大豆鼓粒期的影响是造成大豆减产的主要原因。
O3污染也影响籽粒品质。随着O3浓度的升高,冬小麦籽粒中的蛋白质含量、氨基酸含量、K、Ca、Mg、P、Mn、Cu和Zn元素含量,以及水稻籽粒中的Mg、K、Mn和Cu的含量都呈现增加趋势,但是蛋白质积累量、直链淀粉积累量、支链淀粉积累量和总淀粉积累量则分别降低4.5%—22.7%,6.3%—19.4%,14.10%—27.5%,12.7%—26.1%[73]。此外,高浓度O3还降低了小麦籽粒的容重、硬度、出粉率和加工品质[73]。
国内学者对O3污染造成的农作物损失进行了区域评估。O3污染造成长三角地区的冬小麦和水稻分别减产59.9万t和66.9万t,造成的经济损失分别为9.36亿元和5.39亿元[58];造成长江三角洲地区主要粮食作物(水稻和小麦)的平均产量损失率为5.88%,总经济损失为13.44亿元[74];O3导致油菜平均损失率为5.92%, 产量总损失量为11万t,经济损失2.61亿元[74]。耿春梅等[75]利用O3暴露量AOT40与产量的响应关系,估算出全国范围内由于O3造成水稻、小麦、玉米、豆类、薯类及油菜籽分别减产5.2%—18.4%,10.5%—37.3%,1.80%—6.42%,5.30%—18.90%,2.94%—10.48%,3.17%—11.32%。Aunan等[76]利用全球三维光化学跟踪/输送模型(CTM)对我国主要农作物在未来O3浓度不断升高的情景下的产量变化进行了预测。预计到2020年,O3浓度的升高将导致全国范围内主要农作物的产量损失将比当前大幅增加,其中春小麦、大豆、冬小麦、玉米和水稻的产量损失将分别升至29.3%、28%—35%、13.4%、7.2%和5%。Tang等[77]利用高分辨率的化学输送模型及亚洲区域排放清单,结合实测数据得到的O3剂量响应关系,估算到2020年O3造成的小麦减产率将增加8.1%—9.4%。
4.3 树木生长和生物量
图2显示了目前在我国OTCs对61种木本植物进行O3熏蒸实验的46篇文献整合结果。O3浓度升高平均达到109nmol/mol时,相比于对照O3浓度(34nmol/mol),树木的净光合作用下降28%;总生物量降低14%(升高O3浓度116nmol/mol vs. 对照浓度21nmol/mol),茎生物量降低8%,根生物量降低13%。其它生长指标,如树高、基径和叶面积,都出现不同程度的降低(图2)。O3引起树木光合固碳能力的降低,一方面是由于O3胁迫造成气孔部分关闭,抵御O3进入细胞的同时也降低了光合原材料CO2的摄入,从而导致光合速率降低、生物量降低。另一方面,O3进入植物体破坏了光合作用系统[41],植物叶片在解毒修复过程中对碳的需求增加从而降低了植物叶片同化物向其余部分的转移,导致树木非叶器官(茎和根)的碳固定降低;同时,受损伤的器官将储藏物质以补偿性方式转移供给新叶生长,抑制了茎的生长、侧枝生长、单叶大小[78- 79],植物因此将变得更细[80- 81]。并且,O3引起根系碳分配减少[82],使细根周转率增加、根长缩短、生物量降低,从而改变碳在地上与地下的分配格局[26]。
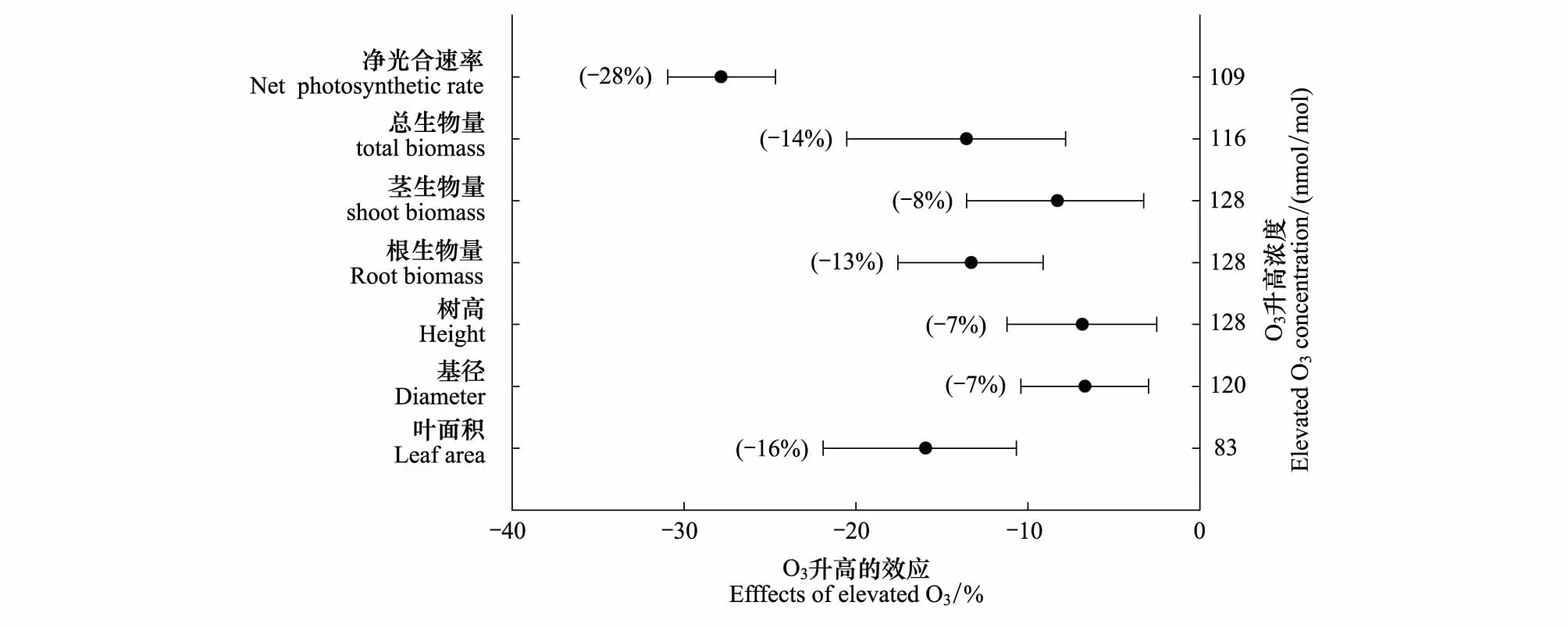
图2 O3浓度升高对中国木本植物的光合、生长和生物量的影响Fig.2 Effects of elevated O3 concentrations on net photosynthetic rate, growth index and biomass of woody plants in China
5 地表臭氧对植物释放BVOCs的影响
植物源挥发性有机化合物(BVOCs)是植物的次生代谢产物之一。它既是地表O3形成的重要前体物,又可与O3发生反应而消耗O3。因此BVOCs的释放与地表O3胁迫之间有着复杂的交互作用[83]。BVOCs在植物应对生物与非生物胁迫中扮演重要的保护角色,O3浓度升高可刺激BVOCs的释放以保护植物光合器官免受氧化伤害[84]。
目前,关于我国地表O3胁迫对BVOCs影响的研究只有少数文献,而结果也无统一定论。Li等[85]观察到短期30天O3熏蒸(白天8:00—17:00, 平均O3浓度80nmol/mol)显著增加中国北方银杏(Ginkgobiloba)夏季单萜类及总BVOCs的释放。Xu等[86]通过一个生长季的O3熏蒸实验(平均O3浓度66.1nmol/mol),发现熏蒸30d和90d都显著增加4年生油松(Pinustabuliformis)BVOCs的释放。然而,与以上研究结果不同,Yuan等[87]发现高浓度O3(80.3nmol/mol)显著降低杨树中部叶片异戊二烯(Isoprene)的释放速率,但对上部叶片的影响较小。O3胁迫对BVOCs释放产生不一致的结论可能与O3浓度、植物品种、熏蒸时间及BVOCs种类的差异有关[88]。此外,Yuan等[87]还发现杨树叶片Isoprene的释放存在“激效剂量反应”(hormetic dose-response),即随着O3熏蒸时间的延长,出现熏蒸前期Isoprene的释放速率增加,后期(当O3积累量达到一定阈值后)Isoprene释放速率降低的现象;并且,在O3熏蒸50d及90d后,Isoprene释放速率都与AOT40和POD(气孔O3吸收通量)呈显著负相关[89]。
尽管国内关于O3胁迫对BVOCs释放的研究仍有大量工作待做,但根据现有报道反映的信息来看,短期低O3浓度能刺激BVOCs释放,尤其是单萜类物质;而长期高O3浓度对BVOCs有着明显抑制作用,其中Isoprene比单萜类物质受到的抑制更明显。这种抑制作用可能与植物光合速率、叶片色素含量及BVOCs合成酶密切相关[84,90]。现有的结果表明,在未来地表O3浓度不断升高的情景下,O3浓度升高能显著降低杨树Isoprene的释放速率,考虑到Isoprene在BVOCs占据几乎50%的总量,这种抑制作用可能有助于减少O3-VOCs限制区域内大气O3及次生气溶胶(SOA)的减轻,减轻因全球温度升高或其他环境因素变化造成的全球BVOCs释放量增高而产生的负面影响。
6 地表臭氧对土壤微生物的影响
国内学者利用OTCs或者O3-FACE,研究了O3浓度升高对土壤微生物生物量、活性、群落结构及功能多样性等的影响,但因实验手段、研究对象、环境条件、臭氧水平、暴露时间等的差异,影响结果存在争议[91-93]。一般来讲,作物到成熟期收割后,能够持续响应O3胁迫的主要是地下部分,因此O3对土壤微生物的影响具有长期累积效应[94]。O3浓度升高通过减少光合产物对根系的分配、改变根系分泌物的量和组成种类等方式间接影响土壤微生物生物量和群落结构及功能多样性。目前,我国有关O3浓度升高对陆地生态系统的影响研究主要集中在地上部分,而对地下土壤微生物影响的研究有限,仅限于在农田生态系统中的若干研究[91-92,95-97]。
O3浓度升高增加了稻麦轮作农田土壤氨氧化细菌和反硝化细菌的数量,但抑制了土壤氨氧化细菌和反硝化细菌的代谢活性[98-99],表明细菌活性与数量的响应机制存在不同;并且,土壤中的固氮微生物数量和群落种类减少[98- 99]。吴芳芳等[97]通过4a的熏气实验发现O3浓度升高降低了麦田土壤微生物量氮、氨氧化细菌的数量和硝化强度,但增加了土壤反硝化细菌数量和反硝化强度,表明随着O3作用时间的累积,农田土壤N2O排放的风险将增加。
O3浓度升高显著降低了小麦和水稻土壤根际微生物功能多样性,并且主要影响根际土壤微生物而对非根际土壤微生物影响不大[95,100-101]。Feng等[102]研究发现O3浓度升高显著降低了水稻田土壤中非产氧光合细菌(AnPPB)的丰度和遗传多样性;并且,对于不同水稻品种,土壤细菌群落组成对O3的响应存在显著差异[91-92]。
7 地表臭氧对土壤温室气体排放的影响
地表O3浓度升高导致地下部分干物质的分配降低[103],根呼吸作用及土壤微生物活性改变[104-106],从而影响土壤的碳氮循环过程,最终导致土壤中二氧化碳(CO2)、氧化亚氮(N2O)及甲烷(CH4)等主要温室气体的释放量变化[107]。目前,我国有关O3对土壤温室气体排放的影响仅限于农田生态系统的一到两年短期的若干研究[107-111]。
O3浓度升高降低了冬小麦和水稻田土壤CO2的排放[108-109,111],但Kou等[107]发现O3浓度升高增加了农田土壤CO2的排放。并且,在植物不同的生长发育阶段,植物-土壤系统中所释放的CO2量也不同;反映出O3对土壤呼吸作用的影响存在着明显的累积效应[107]。O3浓度升高降低了冬小麦土壤N2O的释放。与过滤空气(对照)相比,O3浓度升高到100nmol/mol和150nmol/mol时,N2O累积排放量在冬小麦返青期分别降低8.8%和37.8%,拔节-孕穗期分别降低15.0%和39.1%[111]。
基于稻田土壤的研究表明,O3浓度升高抑制了CH4排放[108-109]。与对照相比,O3浓度升高到75nmol/mol和130nmol/mol时,CH4排放量分别降低42.4%和48.7%。CH4平均释放量与AOT40呈显著负相关,与水稻产量、地上部生物量及地下部生物量呈显著正相关。Tang等[108]利用O3-FACE研究发现O3浓度升高显著降低了水稻(扬稻6号和II-优084)分蘖和开花期的CH4释放量,平均降低量为29.6%。O3引起的CH4的释放量的降低与总生物量、根生物量及最大分蘖数呈正相关关系[108-109]。
基于这些为数不多的研究来看,O3浓度升高不同程度地影响农田土壤CO2、N2O、CH4的排放,一定程度上可以减轻由土壤温室气体排放造成的全球气候变暖的趋势。但考虑到国内相关研究才刚刚起步,而且国外的相关研究发现O3对土壤温室气体排放有抑制、增加或者无影响的不一致结果,因此,这方面仍有大量工作待做。
8 减少臭氧生态环境效应的管理措施
地表O3是二次污染物,因此要降低地表O3浓度,必须控制其前体物(NOx和VOCs)的排放。具体措施如提高汽油品质,在全国大中城市实施欧- 5排放标准,以减少汽油车、柴油车等尾气排放的NOx量;另外,在大城市大力发展和提倡使用公共交通,如地铁、无轨电车、共享单车/电动汽车;并通过增加油价、征收拥堵费、增收停车费等手段减少私家车的使用[12]。本节重点阐述减轻O3对植物伤害的措施以及植物对降低O3浓度的贡献。
(1)不同植物以及同一植物不同品种对O3的敏感性存在差异,因此选育对O3具有较高抗性的优良物种/品种,可预防和减轻植物(农作物和树木)因O3胁迫而带来的负面效应,减少农作物产量及森林固碳的损失。
(2)通过合理的栽培管理措施,如推广喷施O3化学防护剂EDU[69,112-113]、外源抗坏血酸[114],以及接种丛枝菌根(AM)真菌[115],都能有效减轻O3对植物的胁迫伤害。说明此类措施在控制或缓解O3伤害中具有一定的应用前景。
(3)O3通过气孔和角质层进入植物体内,植物吸收O3的同时也降低了空气中O3浓度。这种植物净化污染空气的功能对城市园林绿化、生态环境规划和建设具有直接的指导意义和应用价值[116]。但是值得注意的是,植物排放的BVOCs也是参与O3合成的前体物。因此,筛选高效的净化植物,发挥其在治理O3污染中的积极作用,是植物修复大气污染物的重要应用领域。
9 研究展望
尽管国内已经开展了大量O3对农田生态系统和绿化树种影响的研究工作,但今后还需要在以下几个方面进行重点研究。
(1)监测站点要多在郊区设置。虽然我国已经在各个城市建立了空气质量监测网,目前有国控点、省控点和市控点等2000多个监测点。但这些监测点大部分都分布在各个不同级别的城市中,而对农田和森林区的监测点则非常少。通常来讲,郊区的O3浓度要高于城区,因此当前的空气监测网显然限制了对农区和林区的O3评估精度。今后需要在中国生态系统监测网络(CERN)各站点增加空气质量的监测,使之成为气象监测同等重要的部分。
(2)加强地表O3对地下生态过程的影响研究。目前地下生态过程对地表O3浓度升高的响应机制尚不明确,无法将土壤微生物过程(如微生物生物量、活性、群落组成、多样性、功能等)与土壤关键生物地球化学循环过程,以及与植物地上生态过程关联起来;并且,现有研究主要针对农田生态系统,缺乏对其他生态系统(如森林、草地)的研究。未来的研究应加强O3对地下过程累积效应的长期定位研究,关注土壤-植物相互作用过程,并运用新技术(如同位素技术、分子生物学技术等)将研究不断扩大和深入。
(3)国内开展了大量的O3单因子实验研究,但缺乏多因子同时存在的交互作用研究。目前仅有少量研究O3和CO2增加或N 添加或水分胁迫对农作物和绿化植物的影响。由于自然界是多种环境因子共同作用于生态系统,因此在全球变化的背景下,亟需考虑多个因子(如O3、CO2、水分、N沉降等)同时存在对生态系统服务功能的复合影响研究,进而为陆地生态系统模型(如DLEM等)提供优化参数,提高模型的模拟预测能力。
(4)当前开展全国尺度的O3区域评估大都是根据O3浓度累计值如AOT40得到的剂量-效应(生物量或产量)关系,然后根据大气化学模型模拟的O3浓度来估算区域尺度的O3生态效应。然而,近些年来随着气孔O3吸收量概念的提出,越来越多的研究对不同农作物和绿化植物进行气孔导度参数化,开发不同植物的气孔导度模型来量化气孔O3吸收量—生物量或产量的剂量响应关系。这种评估方法充分考虑了各个区域的环境气象因子和生物因素,在理论上更具有科学性。目前利用气孔O3吸收量评估方法对我国冬小麦进行了全国尺度的O3风险评价[77]。今后需要加强对其他作物和森林树种开展此类研究,科学评估当前环境O3对我国主要粮食作物产量和森林生产力的区域影响。
参考文献(References):
[1] 郑有飞, 吴荣军. 地表臭氧变化特征及其作物响应. 北京: 气象出版社, 2012.
[2] Junge C E. Global ozone budget and exchange between stratosphere and troposphere. Tellus, 1962, 14(4): 363- 377.
[3] Wang T, Xue L K, Brimblecombe P, Lam Y F, Li L, Zhang L. Ozone pollution in China: a review of concentrations, meteorological influences, chemical precursors, and effects. Science of the Total Environment, 2017, 575: 1582- 1596.
[4] Blande J D, Holopainen J K, Li T. Air pollution impedes plant-to-plant communication by volatiles. Ecology Letters, 2010, 13(9): 1172- 1181.
[5] Dentener F, Stevenson D, Ellingsen K, Van Noije T, Schultz M, Amann M, Atherton C, Bell N, Bergma D, Bey I, Bouwman L, Butler T, Cofala J, Collins B, Drevet J, Doherty R, Eickhout B, Eskes H, Fiore A, Gauss M, Hauglustaine D, Horowitz L, Isaksen I S A, Josse B, Lawrence M, Krol M, Lamarque J F, Montanaro V, Müller J F, Peuch V H, Pitari G, Pyle J, Rast S, Rodriguez J, Sanderson M, Savage N H, Shindell D, Strahan S, Szopa S, Sudo K, Van Dingenen R, Wild O, Zeng G. The global atmospheric environment for the next generation. Environmental Science & Technology, 2006, 40(11): 3586- 3594.
[6] 漏嗣佳, 朱彬, 廖宏. 中国地区臭氧前体物对地面臭氧的影响. 大气科学学报, 2010, 33(4): 451- 459.
[7] 王占山, 李云婷, 陈添, 张大伟, 孙峰, 王欣, 郇宁, 潘丽波. 北京城区臭氧日变化特征及与前体物的相关性分析. 中国环境科学, 2014, 34(12): 3001- 3008.
[8] Chan L Y, Chan C Y, Qin Y. Surface ozone pattern in Hong Kong. Journal of Applied Meteorology, 1998, 37(10): 1153- 1165.
[9] Luo C, St John J C, Zhou X J, Lam K S, Wang T, Chameides W L. A nonurban ozone air pollution episode over eastern China: observations and model simulations. Journal of Geophysical Research, 2000, 105(D2): 1889- 1908.
[10] Wang X K, Manning W, Feng Z W, Zhu Y G. Ground-level ozone in China: distribution and effects on crop yields. Environmental Pollution, 2007, 147(2): 394- 400.
[11] Wang T, Wei X L, Ding A J, Poon C N, Lam K S, Li Y S, Chan L Y, Anson M. Increasing surface ozone concentrations in the background atmosphere of Southern China, 1994- 2007. Atmospheric Chemistry and Physics, 2009, 9(16): 6217- 6227.
[12] Feng Z Z, Hu E Z, Wang X K, Jiang L J, Liu X J. Ground-level O3pollution and its impacts on food crops in China: a review. Environmental Pollution, 2015, 199: 42- 48.
[13] Tian H Z, Hao J M. Current status and future trend of nitrogen oxides emissions in China. Preprint Papers-American Chemical Society, Division of Fuel Chemistry, 2003, 48(2): 764- 765.
[14] Zhao B, Wang S X, Liu H, Xu J Y, Fu K, Klimont Z, Hao J M, He K B, Cofala J, Amann M. NOxemissions in China: historical trends and future perspectives. Atmospheric Chemistry and Physics, 2013, 13(19): 9869- 9897.
[15] Wang Y, Zhang Y, Hao J, Luo M. Seasonal and spatial variability of surface ozone over China: contributions from background and domestic pollution. Atmospheric Chemistry and Physics, 2011, 11(7): 3511- 3525.
[16] 万五星, 夏亚军, 张红星, 王娇, 王效科. 北京远郊区臭氧污染及其对敏感植物叶片的伤害. 生态学报, 2013, 33(4): 1098- 1105.
[17] Avnery S, Mauzerall D L, Liu J F, Horowitz L W. Global crop yield reductions due to surface ozone exposure: 2. Year 2030 potential crop production losses and economic damage under two scenarios of O3pollution. Atmospheric Environment, 2011, 45(13): 2297- 2309.
[18] Wieser G, Tegischer K, Tausz M, Häberle K H, Grams T E E, Matyssek R. Age effects on Norway spruce (Piceaabies) susceptibility to ozone uptake: a novel approach relating stress avoidance to defense. Tree Physiology, 2002, 22(8): 583- 590.
[19] Sun G E, McLaughlin S B, Porter J H, Uddling J, Mulholland P J, Adams M B, Pederson N. Interactive influences of ozone and climate on streamflow of forested watersheds. Global Change Biology, 2012, 18(11): 3395- 3409.
[20] Hoshika Y, Watanabe M, Inada N, Koike T. Ozone-induced stomatal sluggishness develops progressively in Siebold′s beech (Faguscrenata). Environmental Pollution, 2012, 166: 152- 156.
[21] Tausz M, Grulke N E, Wieser G. Defense and avoidance of ozone under global change. Environmental Pollution, 2007, 147(3): 525- 531.
[22] Noctor G, Foyer C H. Ascorbate and glutathione: keeping active oxygen under control. Annual Review of Plant Physiology and Plant Molecular Biology, 1998, 49: 249- 279.
[23] Sanmartin M, Drogoudi P D, Lyons T, Pateraki I, Barnes J, Kanellis A K. Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta, 2003, 216(6): 918- 928.
[24] Asada K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiologist, 2006, 141(2): 391- 396.
[25] Feng Z Z, Kobayashi K, Ainsworth E A. Impact of elevated ozone concentration on growth, physiology, and yield of wheat (TriticumaestivumL.): a meta-analysis. Global Change Biology, 2008, 14(11): 2696- 2708.
[26] Wittig V E, Ainsworth E A, Naidu S L, Karnosky D F, Long S P. Quantifying the impact of current and future tropospheric ozone on tree biomass, growth, physiology and biochemistry: a quantitative meta-analysis. Global Change Biology, 2009, 15(2): 396- 424.
[27] Innes J L, Skelly J M, Schaub M. Ozone and Broadleaved Species. A Guide to the Identification of Ozone-Induced Foliar Injury. Bern, Switzerland: Paul Haupt Publishers, 2001: 136- 136.
[28] Manning W J. Detecting plant effects is necessary to give biological significance to ambient ozone monitoring data and predictive ozone standards. Environmental Pollution, 2003, 126(3): 375- 379.
[29] De Vries W, Reinds G J, Posch M, Sanz M J, Krause G H M, Calatayud V, Renaud J P, Dupouey J L, Sterba H, Vel E M, Dobbertin M, Gundersen P, Voogd J C H. Intensive Monitoring of Forest Ecosystems in Europe. Technical Report 2003. Brussels, Geneva: United Nations, Economic Commission for Europe and European Commission, 2003: 163- 163.
[30] Schaub M, Calatayud V. Assessment of visible foliar injury induced by ozone//Ferretti M, Fisher R, eds. Forest Monitoring, Methods for Terrestrial Investigations in Europe with an Overview of North America and Asia, Developments in Environmental Science. Oxford: Elsevier, 2013: 205- 221.
[31] Hayes F, Mills G, Harmens H, Norris D. Evidence of Widespread Ozone Damage to Vegetation in Europe (1990- 2006). Bangor, UK: Programme Coordination Centre for the ICP Vegetation, Centre for Ecology & Hydrology, 2007.
[32] Campbell S J, Wanek R, Coulston J W. Ozone Injury in West Coast Forests: 6 Years of Monitoring. Gen. Tech. Rep. PNW-GTR- 722. Portland, OR: U. S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, 2007: 53- 53.
[33] Schaub M, Calatayud V, Ferretti M, Brunialti G, Lövblad G, Krause G, Sanz M J. Assessment of ozone injury//Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. Hamburg: UNECE, ICP Forests Programme Co-ordinating Centre, 2010.
[34] 张红星, 孙旭, 姚余辉, 万五星, 肖扬, 孙滨峰, Manning W J, 韩春萌, 郜世奇, 高付元, 徐卫华, 冯兆忠, 欧阳志云, 王效科. 北京夏季地表臭氧污染分布特征及其对植物的伤害效应. 生态学报, 2014, 34(16): 4756- 4765.
[35] Feng Z Z, Sun J S, Wan W X, Hu E Z, Calatayud V. Evidence of widespread ozone-induced visible injury on plants in Beijing, China. Environmental Pollution, 2014, 193: 296- 301.
[36] Wan W X, Manning W J, Wang X K, Zhang H X, Sun X, Zhang Q Q. Ozone and ozone injury on plants in and around Beijing, China. Environmental Pollution, 2014, 191: 215- 222.
[37] Zhang W W, Feng Z Z, Wang X K, Niu J F. Responses of native broadleaved woody species to elevated ozone in subtropical China. Environmental Pollution, 2012, 163: 149- 157.
[38] Li P, Calatayud V, Gao F, Uddling J, Feng Z Z. Differences in ozone sensitivity among woody species are related to leaf morphology and antioxidant levels. Tree Physiology, 2016, 36(9): 1105- 1116.
[39] Wittig V E, Ainsworth E A, Long S P. To what extent do current and projected increases in surface ozone affect photosynthesis and stomatal conductance of trees? A meta-analytic review of the last 3 decades of experiments. Plant, Cell & Environment, 2007, 30(9): 1150- 1162.
[40] Yan K, Chen W, He X Y, Zhang G Y, Xu S, Wang L L. Responses of photosynthesis, lipid peroxidation and antioxidant system in leaves ofQuercusmongolicato elevated O3. Environmental and Experimental Botany, 2010, 69(2): 198- 204.
[41] Gao F, Calatayud V, García-Breijo F, Reig-Armiana J, Feng Z Z. Effects of elevated ozone on physiological, anatomical and ultrastructural characteristics of four common urban tree species in China. Ecological Indicators, 2016, 67: 367- 379.
[42] Zhang W W, Feng Z Z, Wang X K, Niu J F. Impacts of elevated ozone on growth and photosynthesis ofMetasequoiaglyptostroboidesHu et Cheng. Plant Science, 2014, 226: 182- 188.
[43] Zhang W W, Niu J F, Wang X K, Tian Y, Yao F F, Feng Z Z. Effects of ozone exposure on growth and photosynthesis of the seedlings ofLiriodendronchinense(Hemsl.) Sarg, a native tree species of subtropical China. Photosynthetica, 2011, 49(1): 29- 36.
[44] Feng Z Z, Niu J F, Zhang W W, Wang X K, Yao F F, Tian Y. Effects of ozone exposure on sub-tropical evergreenCinnamomumcamphoraseedlings grown in different nitrogen loads. Trees, 2011, 25(4): 617- 625.
[45] Ainsworth E A. Rice production in a changing climate: a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration. Global Change Biology, 2008, 14(7): 1642- 1650.
[46] Pang J, Kobayashi K, Zhu J G. Yield and photosynthetic characteristics of flag leaves in Chinese rice (OryzasativaL.) varieties subjected to free-air release of ozone. Agriculture Ecosystems & Environment, 2009, 132(3/4): 203- 211.
[47] Feng Z Z, Pang J, Kobayashi K, Zhu J G, Ort D R. Differential responses in two varieties of winter wheat to elevated ozone concentration under fully open-air field conditions. Global Change Biology, 2011, 17(1): 580- 591.
[48] Xu S, He X Y, Chen W, Su D Y, Huang Y Q. Elevated CO2ameliorated the adverse effect of elevated O3in previous-year and current-year needles ofPinustabulaeformisin urban area. Bulletin of Environmental Contamination and Toxicology, 2014, 92(6): 733- 737.
[49] 邵在胜, 赵轶鹏, 宋琪玲, 贾一磊, 王云霞, 杨连新, 王余龙. 大气CO2和O3浓度升高对水稻‘汕优63’叶片光合作用的影响. 中国生态农业学报, 2014, 22(4): 422- 429.
[50] Yan K, Chen W, Zhang G Y, Xu S, Liu Z L, He X Y, Wang L L. Elevated CO2ameliorated oxidative stress induced by elevated O3inQuercusmongolica. Acta Physiologiae Plantarum, 2010, 32(2): 375- 385.
[51] Li L, Manning W J, Tong L, Wang X K. Chronic drought stress reduced but not protected Shantung maple (AcertruncatumBunge) from adverse effects of ozone (O3) on growth and physiology in the suburb of Beijing, China. Environmental Pollution, 2015, 201: 34- 41.
[52] 陈娟, 曾青, 朱建国, 刘钢, 曹际玲, 谢祖彬, 唐昊冶, 小林和彦. 施氮肥缓解臭氧对小麦光合作用和产量的影响. 植物生态学报, 2011, 35(5): 523- 530.
[53] 罗克菊, 朱建国, 刘钢, 唐昊冶, 李春华, 曾青. 臭氧胁迫对水稻的光合损伤与施氮的缓解作用. 生态环境学报, 2012, 21(3): 481- 488.
[54] 郭建平, 王春乙, 白月明, 温民, 霍治国, 刘江歌, 李雷. 大气中臭氧浓度变化对冬小麦生理过程和籽粒品质的影响. 应用气象学报, 2001, 12(2): 255- 256.
[55] 王亮, 曾青, 冯兆忠, 朱建国, 唐昊冶, 陈曦, 谢祖彬, 刘钢, Kobayashi K. 开放式臭氧浓度升高对2个冬小麦品种光合损伤的研究. 环境科学, 2009, 30(2): 527- 534.
[56] 佟磊, 冯宗炜, 苏德·毕力格, 王琼, 耿春梅, 逯非, 王玮, 殷宝辉, 王效科. 冬小麦气孔臭氧通量拟合及通量产量关系的比较分析. 生态学报, 2012, 32(9): 2890- 2899.
[57] 金明红, 冯宗炜, 张福珠. 臭氧对水稻叶片膜脂过氧化和抗氧化系统的影响. 环境科学, 2000, 21(3): 1- 5.
[58] Feng Z W, Jin M H, Zhang F Z, Huang Y Z. Effects of ground-level ozone (O3) pollution on the yields of rice and winter wheat in the Yangtze River Delta. Journal of Environmental Sciences, 2003, 15(3): 360- 362.
[59] Shi G Y, Yang L X, Wang Y X, Kobayashi K, Zhu J G, Tang H Y, Pan S T, Chen T, Liu G, Wang Y L. Impact of elevated ozone concentration on yield of four Chinese rice cultivars under fully open-air field conditions. Agriculture, Ecosystems &Environment, 2009, 131(3/4): 178- 184.
[60] 杨连新, 王云霞, 赵秩鹏, 朱建国, Sun J D, 王余龙. 自由空气中臭氧浓度升高对大豆的影响. 生态学报, 2010, 30(23): 6635- 6645.
[61] 李彩虹, 李勇, 乌云塔娜, 吴光磊, 蒋高明. 高浓度臭氧对大豆生长发育及产量的影响. 应用生态学报, 2010, 21(9): 2347- 2352.
[62] 张巍巍, 王光华, 王美玉, 刘晓冰, 冯兆忠. 东北春大豆品种东生 1 号对臭氧胁迫的响应. 环境科学, 2014, 35(4): 1473- 1478.
[63] 赵天宏, 曹艳红, 王岩, 戴震, 刘轶欧, 刘波. 臭氧胁迫对大豆根系形态和活性氧代谢的影响. 大豆科学, 2012, 31(1): 52- 57.
[64] 孙加伟, 赵天宏, 付宇, 胡莹莹, 徐玲, 赵艺欣, 史奕. 臭氧浓度升高对玉米活性氧代谢及抗氧化酶活性的影响. 农业环境科学学报, 2008, 27(5): 1929- 1934.
[65] 白月明, 王春乙, 郭建平, 温民. 油菜产量响应臭氧胁迫的试验研究. 农业环境科学学报, 2003, 22(3): 279- 282.
[66] 冯兆忠, 王效科, 郑启伟, 冯宗炜, 谢居清, 陈展. 油菜叶片气体交换对O3浓度和熏蒸方式的响应. 生态学报, 2006, 26(3): 823- 829.
[67] 郑启伟, 王效科, 冯兆忠, 冯宗炜, 欧阳志云, 谢居清. 不同臭氧熏气方式对油菜光合速率、生物量和产量的影响. 生态毒理学报, 2006, 1(4): 323- 329.
[68] 白月明, 王春乙, 温民, 郭建平. 臭氧浓度和熏气时间对菠菜生长和产量的影响. 中国农业科学, 2004, 37(12): 1971- 1975.
[69] 姚芳芳, 王效科, 冯兆忠, 郑飞翔, 冯宗炜, 欧阳志云. EDU对臭氧胁迫下菠菜急性伤害症状和光合特性的影响. 生态环境学报, 2007, 16(5): 1399- 1405.
[70] Wang X K, Zhang Q Q, Zheng F X, Zheng Q W, Yao F F, Chen Z, Zhang W W, Hou P Q, Feng Z Z, Song W Z, Feng Z W, Lu F. Effects of elevated O3concentration on winter wheat and rice yields in the Yangtze River Delta, China. Environmental Pollution, 2012, 171: 118- 125.
[71] Zhu X K, Feng Z Z, Sun T F, Liu X C, Tang H Y, Zhu J G, Guo W S, Kobayashi K. Effects of elevated ozone concentration on yield of four Chinese cultivars of winter wheat under fully open-air field conditions. Global Chang Biology, 2011, 17(8): 2697- 2706.
[72] 佟磊, 王效科, 肖航, 黄仲文. 我国近地层臭氧污染对水稻和冬小麦产量的影响概述. 生态毒理学报, 2015, 10(3): 161- 169.
[73] 郭文善, 朱新开, 赵正, 高春艳, 张如标, 胡海娟, 刘晓成, 孙陶芳, 朱建国, Kobayashi K. 大气O3浓度升高对小麦籽粒产量和品质影响的生理机制 // 第十五次中国小麦栽培科学学术研讨会论文集. 北京: 中国作物学会, 2012.
[74] 姚芳芳, 王效科, 逯非, 冯宗炜, 欧阳志云. 臭氧对农业生态系统影响的综合评估: 以长江三角洲为例. 生态毒理学报, 2008, 3(2): 189- 195.
[75] 耿春梅, 王宗爽, 任丽红, 王燕丽, 王琼, 杨文, 白志鹏. 大气臭氧浓度升高对农作物产量的影响. 环境科学研究, 2014, 27(3): 239- 245.
[76] Aunan K, Berntsen T K, Seip H M, 闵庆文. 中国的地面臭氧及其对农作物产量的可能影响. AMBIO-人类环境杂志, 2000, 29(6): 294- 301.
[77] Tang H Y, Takigawa M, Liu G, Zhu J G, Kobayashi K. A projection of ozone-induced wheat production loss in China and India for the years 2000 and 2020 with exposure-based and flux-based approaches. Global Change Biology, 2013, 19(9): 2739- 2752.
[78] Bortier K, De Temmerman L, Ceulemans R. Effects of ozone exposure in open-top chambers on poplar (Populusnigra) and beech (Fagussylvatica): a comparison. Environmental Pollution, 2000, 109(3): 509- 516.
[79] Matyssek R, Sandermann Jr H. Impact of ozone on trees: an ecophysiological perspective//Esser K, Lüttge U, Beyschlag W, Hellwig F, eds. Progress in Botany. Heidelberg: Springer-Verlag, 2003, 64: 349- 404.
[80] Pretzsch H, Dieler J, Matyssek R, Wipfler P. Tree and stand growth of mature Norway spruce and European beech under long-term ozone fumigation. Environmental Pollution, 2010, 158(4): 1061- 1070.
[81] Karlsson P E, Selldén G, Pleijel H. Establishing Ozone Critical Levels II. UNECE Workshop Report. Gothenburg, Sweden: Swedish Environmental Research Institute, 2003.
[82] Karnosky D F, Pregitzer K S, Zak D R, Kubiske M E, Hendrey G R, Weinstein D, Nosal M, Percy K E. Scaling ozone responses of forest trees to the ecosystem level in a changing climate. Plant, Cell & Environment, 2005, 28(8): 965- 981.
[83] Pinto D M, Blande J D, Souza S R, Nerg A M, Holopainen J K. Plant volatile organic compounds (VOCs) in ozone (O3) polluted atmospheres: the ecological effects. Journal of Chemical Ecology, 2010, 36(1): 22- 34.
[84] Loreto F, Velikova V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiology, 2001, 127(4): 1781- 1787.
[85] Li D W, Chen Y, Shi Y, He X Y, Chen X. Impact of elevated CO2and O3concentrations on biogenic volatile organic compounds emissions fromGinkgobiloba. Bulletin of Environmental Contamination and Toxicology, 2009, 82(4): 473- 477.
[86] Xu S, Chen W, Huang Y Q, He X Y. Responses of growth, photosynthesis and VOC emissions ofPinustabulaeformisCarr. exposure to elevated CO2and/or elevated O3in an urban area. Bulletin of Environmental Contamination and Toxicology, 2012, 88(3): 443- 448.
[87] Yuan X Y, Calatayud V, Gao F, Fares S, Paoletti E, Tian Y, Feng Z Z. Interaction of drought and ozone exposure on isoprene emission from extensively cultivated poplar. Plant, Cell & Environment, 2016, 39(10): 2276- 2287.
[88] Loreto F, Schnitzler J P. Abiotic stresses and induced BVOCs. Trends in Plant Science, 2010, 15(3): 154- 166.
[89] Yuan X Y, Feng Z Z, Liu S, Shang B, Li P, Xu Y S, Paoletti E. Concentration- and flux-based dose-responses of isoprene emission from poplar leaves and plants exposed to an ozone concentration gradient. Plant Cell and Environment, 2017, DOI: 10.1111/pce.13007.
[90] Lichtenthaler H K, Schwender J, Disch A, Rohmer M. Biosynthesis of isoprenoids in higher plant chloroplasts proceeds via a mevalonate-independent pathway. FEBS Letters, 1997, 400(3): 271- 274.
[91] Feng Y Z, Yu Y J, Tang H Y, Zu Q H, Zhu J G, Lin X G. The contrasting responses of soil microorganisms in two rice cultivars to elevated ground-level ozone. Environmental Pollution, 2015, 197: 195- 202.
[92] Zhang J W, Tang H Y, Zhu J G, Lin X G, Feng Y Z. Divergent responses of methanogenic archaeal communities in two rice cultivars to elevated ground-level O3. Environmental Pollution, 2016, 213: 127- 134.
[93] 陈展, 王效科, 冯兆忠, 郑启伟, 欧阳志云. 臭氧对生态系统地下过程的影响. 生态学杂志, 2007, 26(1): 121- 125.
[94] Andersen C P. Source-sink balance and carbon allocation below ground in plants exposed to ozone. New Phytologist, 2003, 157(2): 213- 228.
[95] 陈展, 王效科, 段晓男, 冯兆忠, 吴庆标. 臭氧浓度升高对盆栽小麦根系和土壤微生物功能的影响. 生态学报, 2007, 27(5): 1803- 1808.
[96] 石春红, 郑有飞, 吴芳芳, 刘宏举, 赵泽, 胡程达. 大气中臭氧浓度增加对根际和非根际土壤微生物的影响. 土壤学报, 2009, 46(5): 894- 898.
[97] 吴芳芳, 郑有飞, 吴荣军, 李萍, 王锦旗. 四年O3熏气对小麦根际土壤氮素微生物转化的影响. 生态学报, 2013, 33(24): 7679- 7689.
[98] 李全胜, 林先贵, 胡君利, 张晶, 余永昌, 沈标, 朱建国. 近地层臭氧浓度升高对麦田土壤氨氧化与反硝化细菌活性的影响. 生态与农村环境学报, 2010, 26(6): 524- 528.
[99] 李全胜, 林先贵, 胡君利, 张晶, 余永昌, 沈标, 朱建国. 近地层臭氧浓度升高对稻田土壤氨氧化与反硝化细菌活性的影响. 生态环境学报, 2010, 19(8): 1789- 1793.
[100] Chen Z, Wang X K, Feng Z Z, Xiao Q, Duan X N. Impact of elevated O3on soil microbial community function under wheat crop. Water, Air, and Soil Pollution, 2009, 198(1/4): 189- 198.
[101] Chen Z, Wang X K, Shang H. Structure and function of rhizosphere and non-rhizosphere soil microbial community respond differently to elevated ozone in field-planted wheat. Journal of Environmental Sciences, 2015, 32: 126- 134.
[102] Feng Y Z, Lin X G, Yu Y C, Zhu J G. Elevated ground-level O3changes the diversity of anoxygenic purple phototrophic bacteria in paddy field. Microbial Ecology, 2011, 62(4): 789- 799.
[103] Kanerva T, Palojärvi A, Rämö K, Ojanperä K, Esala M, Manninen S. A 3-year exposure to CO2and O3induced minor changes in soil N cycling in a meadow ecosystem. Plant and Soil, 2006, 286(1/2): 61- 73.
[104] Mörsky S K, Haapala J K, Rinnan R, Tiiva P, Saarnio S, Silvola J, Holopainen T, Martikainen P J. Long-term ozone effects on vegetation, microbial community and methane dynamics of boreal peatland microcosms in open-field conditions. Global Change Biology, 2008, 14(8): 1891- 1903.
[105] Chen Z, Wang X K, Yao F F, Zheng F X, Feng Z Z. Elevated ozone changed soil microbial community in a rice paddy. Soil Science Society of America Journal, 2010, 74(3): 829- 837.
[106] Islam K R, Mulchi C L, Ali A A. Interactions of tropospheric CO2and O3enrichments and moisture variations on microbial biomass and respiration in soil. Global Change Biology, 2000, 6(3): 255- 265.
[107] Kou T J, Cheng X H, Zhu J G, Xie Z B. The influence of ozone pollution on CO2, CH4, and N2O emissions from a Chinese subtropical rice-wheat rotation system under free-air O3exposure. Agriculture, Ecosystems & Environment, 2015, 204: 72- 81.
[108] Tang H Y, Liu G, Zhu J G, Kobayashi K. Effects of elevated ozone concentration on CH4and N2O emission from paddy soil under fully open-air field conditions. Global Change Biology, 2015, 21(4): 1727- 1736.
[109] Zheng F X, Wang X K, Lu F, Hou P Q, Zhang W W, Duan X N, Zhou X P, Ai Y P, Zheng H, Ouyang Z Y, Feng Z W. Effects of elevated ozone concentration on methane emission from a rice paddy in Yangtze River Delta, China. Global Change Biology, 2011, 17(2): 898- 910.
[110] 吴杨周, 胡正华, 李岑子, 陈书涛, 谢燕, 肖启涛. 地表臭氧浓度升高对旱作农田N2O排放的影响. 环境科学, 2015, 36(2): 636- 643.
[111] 胡正华, 李岑子, 陈书涛, 李涵茂, 杨燕萍, 申双和. 臭氧浓度升高对土壤-冬小麦系统CO2排放的影响. 环境科学, 2011, 32(1): 46- 50.
[112] Yuan X Y, Calatayud V, Jiang L J, Manning W J, Hayes F, Tian Y, Feng Z Z. Assessing the effects of ambient ozone in China on snap bean genotypes by using ethylenediurea (EDU). Environmental Pollution, 2015, 205: 199- 208.
[113] Xin Y, Yuan X Y, Shang B, Manning W J, Yang A Z, Wang Y N, Feng Z Z. Moderate drought did not affect the effectiveness of ethylenediurea (EDU) in protectingPopuluscathayanafrom ambient ozone. Science of the Total Environment, 2016, 569- 570: 1536- 1544.
[114] 郑启伟, 王效科, 谢居清, 冯兆忠, 冯宗炜, 倪雄伟, 欧阳志云. 外源抗坏血酸对臭氧胁迫下水稻叶片膜保护系统的影响. 生态学报, 2006, 26(4): 1131- 1137.
[115] 王曙光, 冯兆忠, 王效科, 冯宗炜. 大气臭氧浓度升高对丛枝菌根(AM)及其功能的影响. 环境科学, 2006, 27(9): 1872- 1877.
[116] 陶雪琴, 卢桂宁, 周康群, 刘晖, 党志. 大气化学污染的植物净化研究进展. 生态环境, 2007, 16(5): 1546- 1550.
猜你喜欢
煤气与热力(2021年10期)2021-12-02
今日农业(2020年19期)2020-12-14
红领巾·萌芽(2017年5期)2017-06-23
中学生数理化·八年级物理人教版(2017年12期)2017-04-18
爆笑show(2016年7期)2017-02-09
中国蔬菜(2016年8期)2017-01-15
中国科技信息(2015年2期)2015-11-16
少儿科学周刊·儿童版(2015年10期)2015-11-07
少儿科学周刊·少年版(2015年1期)2015-07-07
少儿科学周刊·儿童版(2015年1期)2015-07-07
