
甜樱桃PacAMR1基因的克隆及表达分析
2018-04-19 08:37:07李京霞吕秀兰
浙江农业学报 2018年4期
李京霞,夏 惠,2,吕秀兰,2,王 进,2,梁 东,2,*
(1.四川农业大学 园艺学院,四川 成都 611130; 2.四川农业大学 果蔬研究所,四川 成都 611130)
抗坏血酸(ascorbic acid,AsA)又名维生素C(vitamin C,Vc),是生物体内重要的抗氧化剂和许多酶的辅因子。AsA不仅在植物生长发育及抗逆过程中具有重要作用[1],且与人类的健康密切相关。因此,研究AsA在植物中的代谢调控机制具有重要意义。植物体内AsA代谢涉及自身合成、运输、循环和降解等几个方面。多项研究结果表明,无论是高AsA含量的猕猴桃(Actinidia)[2-4]或刺梨(Rosaroxburghii)[5]等,还是低AsA含量的苹果(Malusdomestics)[6]和番茄(Lycopersiconesculentum)[7]等,自身合成是植物体内AsA含量的决定因子,而且还发现绝大多数植物都是通过L-半乳糖合成AsA,其他几种途径(L-古洛糖途径、D-半乳糖醛酸途径和肌醇途径)可能只在一些植物中发挥辅助作用[8-9]。
L-半乳糖途径是植物合成AsA最重要的途径,也是目前研究的最为透彻的途径。该途径中涉及的所有基因也已在多种植物中获得,如拟南芥(Arabidopsisthaliana)[10]、苹果[4]、刺梨[5]、猕猴桃[4]、柑橘(Citrus)[11]、番茄[12]、蓝莓(Vacciniumcorymbosum)[13]和枣(Ziziphusjujuba)[14]等。随着植物体内AsA的合成途径及相关基因的鉴定和功能验证,目前关于AsA研究的热点已经转变为AsA在植物体内合成的调控机制方面,即研究有哪些因子可以调控L-半乳糖途径,及其通过何种方式进行调控。
一些研究表明,F-box类蛋白往往可以作为SCF(Skp1-Cullin-F-box)类E3泛素连接酶复合体的一部分,通过泛素化调控下游靶基因的表达[15-16]。2009年,Zhang等[17]从拟南芥臭氧敏感型突变体中克隆得到AMR1(ascorbic acid mannose pathway regulator 1)基因,功能位点分析发现该基因编码的氨基酸N端和C端分别有F-box功能域和由48个氨基酸残基构成的DUF295功能域。研究进一步发现,AMR1基因的表达在拟南芥中能够抑制L-半乳糖合成途径中GME(GDP-D-mannose-3',5'-epimerase)、GMP(GDP-D-mannose pyrophosphorylase)、GGP(GDP-L-galactose phosphorylase)和GPP(L-galactose-1-phosphate phosphatase)等基因的转录,负调控AsA的合成。且AMR1的表达受环境因子和植物自身条件的影响,在胁迫条件下或一些衰老组织中表达水平升高,这与AsA含量呈负相关。以上研究表明,拟南芥AMR1可能作为SCF复合物的一部分,通过泛素化调控AsA的合成,但其具体的作用机制目前还不清楚。
甜樱桃被誉为“早春第一果”,经济价值极高,在我国很多温带地区都有种植。课题组前期的研究发现,AsA在甜樱桃果实生长过程中的含量变化明显,而不同生长阶段叶片中的AsA含量差异也很显著,这些特性为我们研究AsA调控机制提供了很好的材料。为了研究甜樱桃AsA生物合成调控机制,本研究拟通过从甜樱桃中克隆出PacAMR1基因,并检测该基因在果实生长期和不同发育时期叶片中的表达模式,分析PacAMR1表达与抗坏血酸含量变化的相关性。研究结果将为更深入地认识植物AsA合成的调控机制和通过生物技术手段改良植物AsA水平奠定基础。
1 材料与方法
1.1 植物材料
以甜樱桃品种佐藤锦(Prunusaviumcv. Satonishiki)为实验材料,供试样品均采自四川雅安市汉源县九襄甜樱桃示范园。于2016年3—6月,在谢花后0、10、20、30、40和50 d分别取正常发育的果实。在果实成熟时,选取树冠外围当年生枝,分别选取枝条顶端、中部和基部叶片作为幼叶、成熟叶和老叶。所有样品迅速用液氮速冻,放超低温保存待用。
1.2 果实总RNA提取
采用改良的CTAB法提取RNA[18],并测定核酸浓度及质量,存于-80 ℃冰箱备用。
1.3 甜樱桃AMR1基因全长的获得
根据GenBank中其他植物(详见图3)AMR1基因氨基酸序列和项目组前期对甜樱桃转录组测序的结果,利用Primer 5.0软件设计甜樱桃AMR1全长引物(表1)。
按照TaKaRa公司反转录试剂盒的说明书进行反转录,获得cDNA第一链后进行聚合酶链式反应,PCR反应体系为25 μL,反应程序:94 ℃ 预变性5 min;94 ℃变性45 s,50 ℃退火45 s,72 ℃延伸90 s,变性到延伸35个循环;接着72 ℃ 10 min,最后保存于4 ℃。参照AxyPrep DNA凝胶回收试剂盒说明书进行操作,回收和纯化目的DNA片段,连接到pMD 19-T vector上,重组质粒导入大肠埃希菌感受态细胞中,进行克隆、测序。利用DNAMAN 软件将所得的片段拼接,得到PacAMR1全长cDNA序列。
1.4 甜樱桃AMR1的相关生物信息学分析
根据所测得的核苷酸序列,利用DNAStar软件分析其序列,并在ORF Finder(https://www.ncbi.nlm.nih.gov/orffinder/)上找出其开放阅读框;利用EMBL(https://www.embl.org)分析氨基酸的保守功能域。从GenBank(https://www.ncbi.nlm.nih.gov/genbank/)中挑选来源于不同植物的AMR1氨基酸序列,用Blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行同源性比对,用MEGA5.0软件进行进化树的构建。
1.5 甜樱桃AMR1的表达分析
根据克隆得到的目的基因AMR1基因序列,用Primer Express 3.0 软件设计定量表达分析所需的引物序列(表1)。分别提取不同样品的总RNA,使用DNase Ⅰ处理后,利用PrimeScript RT Reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa)进行反转录,按照SYBR Premix Ex TaqTM(Tli RNaseH Plus)试剂盒(TaKaRa)在荧光定量PCR仪上进行Real-time PCR扩增。Real-time PCR反应体系:SYBR Green qPCR SuperMix 10 μL,上游引物 0.8 μL,下游引物0.8 μL,cDNA模板1.5 μL,ddH2O 6.9 μL。反应程序:95 ℃ 10 min;95 ℃ 10 s;60 ℃ 31 s;72 ℃ 20 s;熔解曲线反应。共40个循环。PCR扩增采用BIO-RAD公司iQ5TM型实时定量 PCR仪进行,利用 iO5TMReal-Time PCR Detection System 进行基因表达数据分析。数据分析采用2-ΔΔct法,重复3次。
表1本研究所用引物
Table1Primers used in this study

引物名称Primer引物核苷酸序列Nucleotideofsequences引物用途UsageofprimersPacAMR1-S5'-CCACTCTGCCAAGTCAGTCA-3'基因克隆GenecloningPacAMR1-A5'-GGGATTGGATTTTTCCACCT-3'基因克隆GenecloningPacAMR-REAL-S25'-CCACTCTGCCAAGTCAGTCA-3'基因表达GeneexpressionPacAMR-REAL-A25'-TCGTCAGCCATAGCTCCTTT-3'基因表达Geneexpression
1.6 甜樱桃果实不同时期AsA含量的测定
冷冻组织2.0 g以提取AsA[5]。HPLC法测定AsA,使用带有VWD检测器和SB-Zobax C18柱(150 mm×4.6 mm,5 μm)的Agilent 1260型高效液相色谱仪进行测量。流动相为15%甲醇和pH为2.5的85%偏磷酸水溶液组成。流速1 mL·min-1,进样体积10 μL,柱温度35 ℃,检测波长243 nm。
2 结果与分析
2.1 PacAMR1基因全长cDNA的获得及序列分析
以反转录cDNA为模板,利用特异引物PacAMR1-S和PacAMR1-A进行基因完整编码区的扩增,产物经1%的琼脂糖凝胶电泳,在1 300 bp左右有单一明亮的条带,与预测片段大小相符(图1)。对该片段进行回收、连接、转化、重组质粒导入大肠埃希菌感受态细胞以及阳性克隆的筛选,送成都擎科生物公司测序。测序正确的序列命名为PacAMR1。
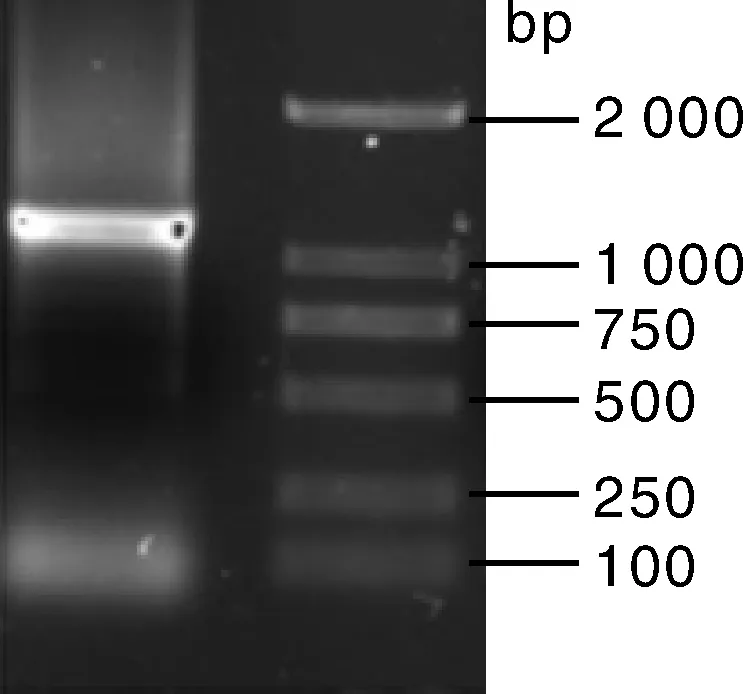
图1 甜樱桃PacAMR1 cDNA全长RT-PCR扩增Fig.1 PCR amplification of full-length cDNA of PacAMR1 in sweet cherry
利用EditSeq软件分析测序结果表明,克隆获得PacAMR1基因cDNA全长为1 327 bp,该基因完整开放阅读框为1 206 bp(图2),位于23~1 228 bp,编码401个氨基酸,分子量为25.78 ku,等电点为7.144,带正电荷残基总数45,带负电荷残基总数46,属于稳定类蛋白。在氨基酸N端和C端分别有F-box和DUF295功能域(图2),与拟南芥同源基因AMR1有相似的结构域。
NCBI Blast(https://blast.ncbi.nlm.nih.gov/Blast.cgi)和DNAMAN的分析结果表明,PacAMR1编码的氨基酸与甜樱桃(Prunusavium,XP_021819517.1)、梅(Prunusmume,XP_016651609.1)、桃(Prunuspersica,XP_020424646.1)、白梨(Pyrusbretschneideri,XP_009337088.1)和苹果(Malusdomestica,XP_008337956.1)的AMR1蛋白序列同源性分别为99%、97%、96%、77%和77%,表明PacAMR1与其他植物的AMR1氨基酸序列具有较大的相似性(图3)。

ATG,起始密码子; *,终止密码子。被框住的区域分别为F-box和DUF295功能域。ATG, Initiation codon; *, Termination codon. Boxed area was F-box and DUF295 domain.图2 PacAMR1基因序列及推测的氨基酸序列Fig.2 Full-length cDNA and its protein sequence of PacAMR1

Ⅰ~Ⅵ依次为甜樱桃Prunus avium AMR1,桃Prunus persica XP_020424646.1,梅Prunus mume XP_016651609.1,甜樱桃Prunus avium XP_021819517.1,白梨Pyrus bretschneideri XP_009337088.1,苹果Malus domestica XP_008337956.1。Ⅰ-Ⅵ represented sweet cherry Prunus avium AMR1, peach Prunus persica XP_020424646.1, plum blossom Prunus mume XP_016651609.1, sweet cherry Prunus avium XP_021819517.1, pear Pyrus bretschneideri XP_009337088.1, apple Malus domestica XP_008337956.1, respectively.图3 PacAMR1基因编码的氨基酸序列与其他相关序列的多重对比Fig.3 Alignment of multiple sequences of PacAMR1 protein amino acid sequence with that of other plants
选择GenBank 中其他7种植物的AMR1氨基酸序列与PacAMR1对比分析后,使用MEGA5.0软件中的NJ法(Neighbor-Joining)制作进化树(图4)。从进化树中可以看出,甜樱桃与桃树、梅聚为一类。
2.2 PacAMR1基因在甜樱桃果实发育过程中的表达及AsA含量的测定
由图5-A可以看出,在果实开始生长发育到花后第10天时,PacAMR1基因表达量下降,大约下降1倍左右;第10~30天,PacAMR1基因表达量缓慢上升;第30天的表达量约为10 d的3倍;第30~40天,PacAMR1基因表达量迅速上升并达到峰值,为第10天的7倍左右;从果实生长第40天以后,PacAMR1基因表达量下降。从整体趋势看,随着果实发育成熟,PacAMR1基因的表达量是逐渐上升的。在第10天时,PacAMR1基因的表达量最低,之后甜樱桃果实在发育第40天时其表达量达到最大。
使用高效液相色谱法测定甜樱桃果实成熟过程中AsA含量变化,由图5-B可以看出,在果实开始生长发育到第10天时,AsA含量上升到峰值;第10~20天,AsA含量迅速下降到最低值;第20~50天,AsA含量缓慢上升;但上升到第50天时,其AsA含量也要比第0天低。从整体趋势看,随着果实发育成熟AsA含量是下降的。在果实第10天时,AsA含量最高。

图4 不同植物PacAMR1基因编码的氨基酸序列的系统进化Fig.4 Phylogenetic evolution of amino acid sequences encoded by different plant PacAMR1

图5 甜樱桃果实发育过程PacAMR1基因表达量(A)及ASA含量(B)变化Fig.5 PacAMR1 gene expression (A) and ascorbic acid content (B) during fruit development of sweet cherry
2.3 PacAMR1基因在不同发育时期叶片的表达及AsA含量的测定
由图6-A发现,在幼叶期,PacAMR1基因的表达量最低,从幼叶到成熟叶,PacAMR1基因的表达量缓慢上升,且表达量上升地较少,从成熟叶到老叶,PacAMR1基因的表达量迅速上升,且达到峰值。从整体上看,随着叶片的生长PacAMR1基因的表达量呈上升趋势。
使用高效液相色谱法测定不同叶龄中AsA含量变化,由图6-B可以看出,幼叶的AsA含量最高,从幼叶到成熟叶,AsA含量缓慢下降,且下降的数量较少,从成熟叶到老叶,AsA含量快速下降,在老叶中AsA含量最低。整体上看,随着叶片的生长发育,AsA含量呈下降趋势变化。

图6 甜樱桃不同叶龄PacAMR1基因表达量(A)及AsA含量(B)变化Fig.6 PacAMR1 gene expression (A) and ascorbic acid content (B) in different leaf development stages of sweet cherry
3 讨论
目前为止,抗坏血酸生物合成的代谢通路已经有明确结论[8-10],但对这些合成途径的调控机制尚不明确,所以研究抗坏血酸合成调控机制具有重要意义。本研究从甜樱桃果实中克隆出一条AMR1基因,通过对其核苷酸和推测的蛋白质序列的分析发现,PacAMR1基因cDNA全长为1 327 bp,在位于23~1 228 bp有完整开放阅读框,长度为1 206 bp,编码401个氨基酸,且PacAMR1与其他植物的AMR1基因具有很高的同源性(图3、图4),表明我们从佐藤锦甜樱桃中获得了AMR1基因。
氨基酸序列分析的结果表明,在PacAMR1编码的蛋白质序列的N端和C端分别具有F-box和DUF295功能域(图2)。以前的研究表明,具有F-box功能域是F-box类蛋白家族的典型特征[19-21],这说明本研究获得的PacAMR1属于F-box类蛋白。F-box类蛋白是SCF类E3泛素连接酶复合物的一部分,通常通过泛素化对下游目标蛋白进行调控,而通过泛素化降解目标蛋白是包括开花、生物节律、根系发育和光信号感受在内的多个生物学进程的重要调控机制[19-21]。这些结果也暗示着PacAMR1在甜樱桃的各种生理进程中可能发挥着重要的调控功能。
Zhang等[17]的研究发现,拟南芥中AMR1的表达与GME、GMP、GGP和GPP基因的表达和AsA的含量呈相反的趋势。在本研究中,我们也发现随着果实发育成熟,PacAMR1基因的表达量是逐渐上升的,而AsA含量是下降的(图5)。PacAMR1的表达在幼叶中最低,在老叶中最高,但幼叶中的AsA含量最高,老叶中ASA含量最低。这些结果表明,PacAMR1的表达与AsA含量呈负相关,PacAMR1可能通过某种机制反向调控甜樱桃中AsA的生物合成。本课题组以前对甜樱桃果实生长过程L-半乳糖途径相关基因表达模式有过研究[22],对比这些基因的表达模式与本文中PacAMR1表达模式,我们也发现在果实生长过程中PacAMR1的表达与L-半乳糖途径中的GME、GMP、GGP和GPP等的表达呈相反趋势。综合这些发现,我们更进一步地认为PacAMR1可能作为泛素化体的一部分,通过泛素化影响L-半乳糖途径中相关基因的表达,从而调控植物体中AsA的生物合成,但这些还需要进一步的互作和转基因等实验来验证。
参考文献(References):
[1]FOYER C H, NOCTOR G.Ascorbate and glutathione: The heart of the redox hub[J].PlantPhysiology, 2011, 155(1):12-18.
[2]LAING WA, BULLEY S, WRIGHT M, et al. A highly specific L-galactose-1-phosphate phosphatase on the path to ascorbate biosynthesis[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2004, 101(48):16976-16981.
[3]BULLEY S M, RASSAM M, HOSER D, et al. Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis[J].JournalofExperimentalBotany, 2009, 60(3):765-778.
[4]LI M, MA F, LIANG D, et al. Ascorbate biosynthesis during early fruit development is the main reason for its accumulation in kiwi[J].PLoSOne, 2010, 5(12): e14281.
[5]HUANG M, XU Q, DENG X X. L-Ascorbic acid metabolism during fruit development in an ascorbate-rich fruit crop chestnut rose (RosaroxburghiiTratt)[J].JournalofPlantPhysiology, 2014, 171(14):1205-1216.
[6]LI M, CHEN X, WANG P, et al. Ascorbic acid accumulation and expression of genes involved in its biosynthesis and recycling in developing apple fruit[J].JournaloftheAmericanSocietyforHorticulturalScience, 2011, 136(4):231-238.
[7]IOANNIDI E, KALAMAKI M S, ENGINEER C, et al. Expression profiling of ascorbic acid-related genes during tomato fruit development and ripening and in response to stress conditions[J].JournalofExperimentalBotany, 2009, 60(2):663-678.
[8]CRUZ-RUS E, BOTELLA M A, VALPUESTA V, et al. Analysis of genes involved in L-ascorbic acid biosynthesis during growth and ripening of grape berries[J].JournalofPlantPhysiology, 2010, 167(9):739-748.
[10]LINSTER C L, CLARKE S G. L-ascorbate biosynthesis in higher plants: the role of VTC2[J].TrendsinPlantScience, 2008, 13(11):567-573.
[12]YE J, HU T X, YANG C M, et al. Transcriptome profiling of tomato fruit development reveals transcription factors associated with ascorbic acid, carotenoid and flavonoid biosynthesis[J].PLoSOne, 2015, 10(7): e0130885.
[13]LIU F H, WANG L, GU L, et al. Higher transcription levels in ascorbic acid biosynthetic and recycling genes were associated with higher ascorbic acid accumulation in blueberry[J].FoodChemistry, 2015, 188: 399-405.
[14]ZHANG C M, HUANG J, LI X G. Transcriptomic analysis reveals the metabolic mechanism of l-ascorbic acid inZiziphusjujubaMill.[J].FrontiersinPlantScience, 2016, 7: 122.
[15]MOON J, PARRY G, ESTELLE M. The ubiquitin-proteasome pathway and plant development[J].PlantCell, 2004, 16(12): 3181-3195.
[16]NI W, XIE D, HOBBIE L, et al. Regulation of flower development in Arabidopsis by SCF complexes[J].PlantPhysiology, 2004, 134(4): 1574-1585.
[17]ZHANG W, LORENCE A, GRUSZEWSKI H A, et al. AMR1, an Arabidopsis gene that coordinately and negatively regulates the mannose/L-galactose ascorbic acid biosynthetic pathway[J].PlantPhysiology, 2009, 150(2):942-950.
[18]GASIC K, HERNANDEZ A, KORBAN S S. RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction[J].PlantMolecularBiologyReporter, 2004, 22(4):437-438.
[19]BAI C, SEN P, HOFMANN K, et al. SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box[J].Cell, 1996, 86(2): 263-274.
[20]XIAO W Y, JANG J C. F-box proteins in Arabidopsis[J].TrendsinPlantScience, 2000, 5(11):454-457.
[21]WOO H R, CHUNG K M, PARK J H, et al. ORE9, an F-box protein that regulates leaf senescence in Arabidopsis[J].PlantCell, 2001, 13(8): 1779-1790.
[22]LIANG D, ZHU T T, NI Z Y, et al. Ascorbic acid metabolism during sweet cherry (Prunusavium) fruit development[J].PLoSOne, 2017, 12(2): e0172818.
猜你喜欢
环球时报(2022-09-20)2022-09-20 15:18:57
金桥(2022年2期)2022-03-02 05:43:04
今日农业(2020年24期)2020-12-15 16:16:00
中国洗涤用品工业(2019年4期)2019-05-11 09:27:18
人大建设(2018年5期)2018-08-16 07:09:08
中成药(2018年1期)2018-02-02 07:20:05
喜剧世界(2017年13期)2017-07-24 15:43:42
兽医导刊(2016年12期)2016-05-17 03:51:50
动物医学进展(2015年10期)2015-12-07 05:46:19
现代检验医学杂志(2015年4期)2015-02-06 02:02:06
