
岩溶石灰土微生物丰度的影响因素及其指示意义
2018-04-16 05:58:43朱美娜梁月明刘畅靳振江李强
生态环境学报 2018年3期
朱美娜,梁月明*,刘畅,靳振江,李强*
1. 中国地质科学院岩溶地质研究所,国土资源部//广西岩溶动力学重点实验室,广西 桂林 541004;2. 联合国教科文组织国际岩溶研究中心,广西 桂林 541004;3. 桂林理工大学环境科学与工程学院,广西 桂林 541006
土壤微生物作为土壤生态系统重要的组成部分,不仅参与土壤养分循环(刘丽等,2013),还对外界环境变化反应非常敏感(贺纪正等,2013;李瑞等,2017),因此土壤微生物具有调节生态系统的功能及指示作用。微生物的指示作用已在土壤重金属污染、石漠化恢复以及土壤质量评估方面得到应用(李韵诗等,2015;朱新玉等,2014;Xie et al.,2015;周卫红等,2017),还有学者(贺凤鹏等,2016)利用土层之间的微生物学特征来评价草地生态系统的健康状况,从而为利用土壤微生物评价土壤质量提供新思路和依据。
岩溶土壤主要以发育于碳酸盐岩母质上的非地带性土壤——石灰土为主,其隶属于初育土纲,石质初育土亚纲,石灰(岩)土类,下有黑色石灰土、棕色石灰土、黄色石灰土和红色石灰土4个亚类(张美良等,1994)。不同亚类的土壤其物理、化学、生物学性质都有所差异。尽管关于岩溶生态系统中土壤的理化性质已有较深入的研究和报道(邓艳等,2009;杨柳等,2011;蓝家程等,2011),但有关不同亚类石灰土的生物学特征及土壤质量的研究相对薄弱。冯书珍等(2015)探究了岩溶区原生林不同坡位土层(0~10、30~50和70~100 cm)对土壤微生物丰度的影响,认为微生物丰度对土层的响应强于坡位,并且微生物丰度受土壤养分中碱解氮、速效磷、速效钾影响显著。也有学者(葛云辉等,2012)以广西环江县岩溶区的棕色石灰土、黑色石灰土为研究对象,探究添加外源14C-稻草和Ca14CO3对土壤微生物群落丰度的影响。然而,很少有学者细化土壤梯度分析,更极少有学者将微生物与土壤类型关系及石灰土成土机理相联系。此外,关于利用土层之间土壤微生物学特征的差异评价岩溶石灰土土壤质量还鲜有报道。
本研究以广西人为干扰较弱的两种典型石灰土(黄色石灰土和红色石灰土)为研究对象,应用实时荧光定量 PCR技术探讨细菌与真菌丰度对土壤类型及层位(0~10 cm 表层土,10~20、20~40、40~60、60~80和80~100 cm)的响应,旨在探究岩溶石灰土微生物丰度的影响因素,为利用土壤微生物丰度在层位之间的特征差异评价岩溶区土壤质量研究提供理论依据。
1 材料和方法
1.1 研究样区
黄色石灰土取自百色市(24°42'N,106°30'E),红色石灰土取自来宾市(23°25'N,109°30'E)。百色市气候属亚热带季风气候,年平均气温为19.0~22.1 ℃,年平均降雨量约为 1114.9 mm;来宾市地处中亚热带向南亚热带过渡的季风气候区,年平均气温为 20.3 ℃,年平均降雨量约为 1360 mm。在人为干扰少的地方布设样方,约有10~15 a的休耕史,每个样方保持坡位(中坡位)、坡向(北坡)一致。黄色石灰土研究区主要分布着小果蔷薇(Rosa cymosa Tratt)、缫丝花(Rosa roxburghii Tratt)和火棘(Pyracantha fortuneana)。红色石灰土研究区主要分布着桃金娘(Rhodomyrtus tomentosa)、檵木(Loropetalum chinense)和紫牡丹(Paeonia delavayi Franch)。
1.2 土壤样品采集
土壤样品采集于2015年11月,每个样方10 m×10 m,共10个样方(包括4个黄色石灰土样方和6个红色石灰土样方)。分别采集富含腐殖质的表层土,10~20、20~40、40~60、60~80 和 80~100 cm层土样,共计 60个土壤样品。土壤样品带回实验室经处理后,一部分保存于-80 ℃冰箱中用于土壤微生物分析,一部分经风干后用于土壤理化性质分析。
1.3 土壤基本理化性质的测定
土壤有机碳用重铬酸钾-硫酸氧化法测定(Bremner,1965);土壤速效磷经 0.5 mol∙L-1NaHCO3浸提后用钼锑抗比色法测定(Colwell,1963);溶解有机碳采用 TOC仪测定(盛浩等,2015);全氮采用半微量凯式法测定(鲍士旦,2000)。
1.4 土壤DNA提取和浓度测定
按照MOBIO公司土壤DNA快速提取试剂盒说明书提取土壤微生物总DNA,提取的DNA样品经1%琼脂糖凝胶电泳检测大小约为20 kb;利用微量紫外分光光度计(Quawell 5000,美国)测定DNA浓度(c)和纯度(p)。DNA 样品置于-20 ℃冰箱中保存,以供下游分子生物学实验使用。
1.5 实时荧光定量PCR
采用荧光定量PCR测定细菌16S rRNA、真菌18S rRNA基因拷贝数。细菌引物为F338:5′-CT ACG GGA GGC AGC AG-3′;R518:5′-ATT ACC GCG GCT GCT GG-3′(Manerkar et al.,2008)。真菌引物为 FUNGI:5′-GTA GTC ATA TGC TTG TCT C-3′;NSI:5′-ATT CCC CGT TAC CCG TTG-3′(Li et al.,2010)。细菌反应体系为 25 μL,模板 DNA(genomic DNA,5 ng∙μL-1)1 μL,Green-2-Go qPCR Mastermix(Sangon Biotech,上海)12.5 μL,去离子无菌水9.5 μL,10 μm∙μL-1的正向引物和反向引物各 1 μL。真菌反应体系为 25 μL,模板 DNA(5 ng∙μL-1)1 μL,Green-2-Go qPCR Mastermix(Sangon Biotech,上海)12.5 μL,去离子无菌水 10.7 μL,10 μm·μL-1的正向引物和反向引物各0.4 μL。
16S rRNA的扩增程序为95 ℃预变性3 min,95 ℃变性45 s,56 ℃退火30 s,72 ℃延伸30 s,30个循环,最后72 ℃延伸1 min。18S rRNA的扩增程序为95 ℃预变性15 min,95 ℃变性1 min,57 ℃退火1 min,72 ℃延伸1 min,39个循环,最后72 ℃延伸5 min。
微生物丰度 A(即核酸拷贝数,copies∙mL-1)计算公式(梁月明等,2017)如下:

式中,l为DNA长度。
1.6 统计分析
运用SPSS 19.0进行数据统计分析。差异显著性采用One-way ANOVA(单因素方差分析)进行分析,多重比较采用LSD法,相关性采用Pearson相关系数法(双尾检验)进行分析。此外,还运用Canoco 5进行冗余分析(Redundancy Analysis,RDA)。
2 结果与分析
2.1 不同土壤类型各土层的理化性质
由表1可知,黄色石灰土表层土土壤有机碳、全氮及速效磷质量分数均显著高于红色石灰土表层土,而两种石灰土在其他层位间无显著差异。由此说明,两种石灰土的养分差异性主要体现在表层上。
此外,黄色石灰土土壤有机碳质量分数与全氮质量分数均表现出随土层加深而依次递减,且表层土的速效磷质量分数极显著高于其他土层(P=0.004)。同时,红色石灰土土壤有机碳质量分数也呈现出随土层加深而依次递减,并且10~20 cm土层速效磷质量分数显著高于其下层位的土壤速效磷质量分数(P=0.012)。
2.2 土壤类型与土层对微生物丰度的影响
由图1可知,黄色石灰土与红色石灰土细菌丰
度差异性亦主要体现在表层上。两种石灰土细菌丰度均表现为随土层加深而逐渐降低,其在层位间的差异性表现为,黄色石灰土表层土与20 cm以下土层细菌丰度差异显著(P=0.027)。红色石灰土表层土与 40 cm以下土层细菌丰度差异显著(P=0.023)。两种石灰土真菌丰度都以表层土丰度最高,但随土层加深并未呈规律递减。
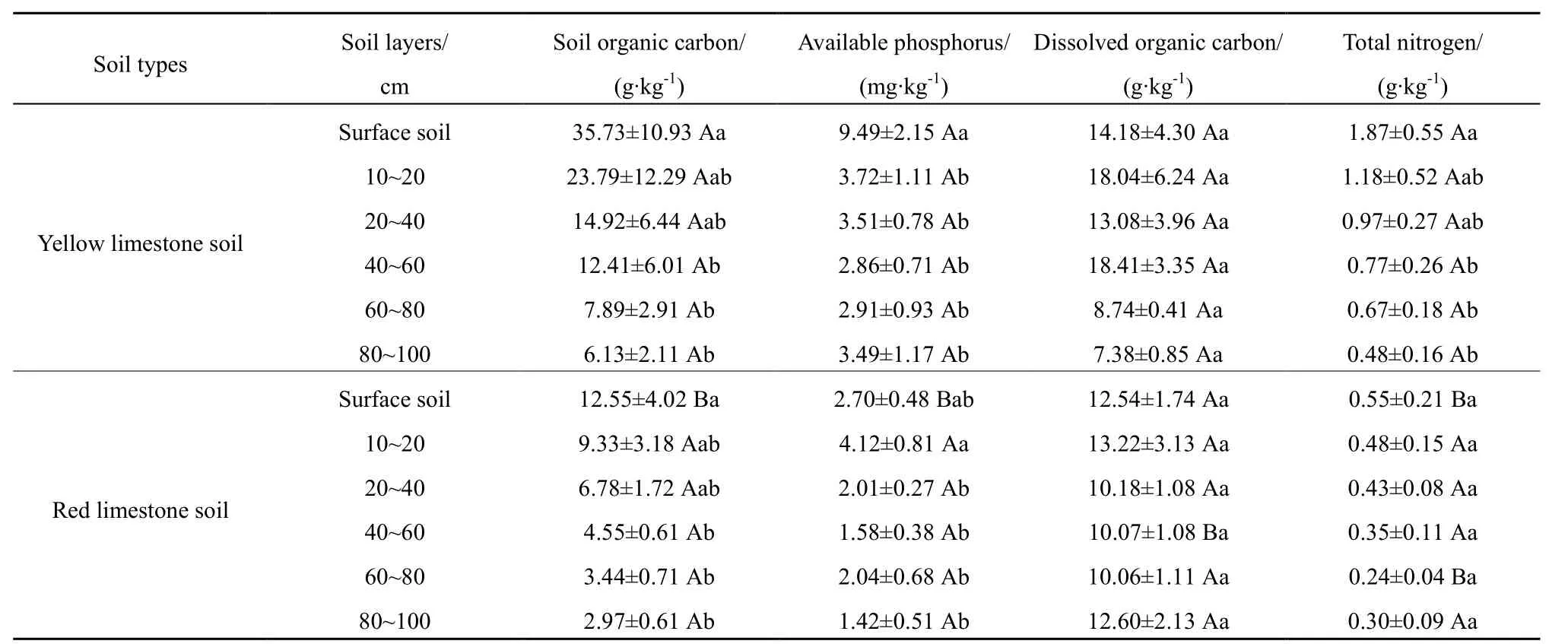
表1 两种石灰土土壤理化性质在不同土层间的比较Table 1 Comparison on soil physicochemical properties among two kinds of limestone soil types and soil layers

图1 微生物丰度在土壤类型和土层间的比较Fig. 1 Comparison on soil microbial abundance among different soil types and soil layers
2.3 岩溶石灰土微生物丰度的影响因素
由表2可知,土壤类型对细菌丰度具有显著影响(F=4.413,P=0.041),对真菌丰度影响不显著(F=0.016,P=0.901)。土层对细菌丰度具有极显著影响(F=7.225,P=0.000),对真菌丰度具有显著影响(F=2.732,P=0.030),从 F值来看,土层对细菌丰度的影响高于真菌丰度。土壤类型与土层对细菌与真菌丰度的交互作用不明显。

表2 土壤类型及土层对细菌与真菌丰度的双因素方差分析结果Table 2 Effect of soil type, soil layers and their interaction on bacterial and fungal abundance by two-way ANOVA
由表3可知,,细菌丰度与土壤有机碳、速效磷、全氮均存在极显著正相关关系(P=0.000),与溶解有机碳呈显著正相关关系(P=0.015)。真菌丰度与土壤有机碳相关性也达到了极显著水平(P=0.005),与速效磷(P=0.011)和全氮(P=0.012)则呈显著正相关关系。不仅微生物与土壤因子存在显著或极显著相关关系,微生物之间即细菌丰度与真菌丰度也呈极显著正相关关系(P=0.000)。

表3 土壤理化性质与微生物丰度Pearson相关性分析Table 3 Pearson correlation among soil physicochemical properties and microbial abundance
RDA分析结果(表4)表明,土壤有机碳、全氮、速效磷和溶解有机碳对微生物丰度影响的解释率分别为31.8%、27.2%、21.1%和1.7%,前3种土壤因子累计解释率为80.1%,说明土壤有机碳、全氮、速效磷是影响微生物丰度的主要土壤因子。

表4 土壤微生物丰度主要影响因子Table 4 The main influencing factors of soil microbial abundance
3 讨论
3.1 不同石灰土土壤理化性质的影响因素
不同亚类石灰土成土年龄、所经历的溶蚀、地形地貌、地上植被、风化程度、淋溶程度以及富钙、镁的岩溶地球化学背景不同,会导致土壤间理化性质差异(张美良等,1994;曹建华等,2003;李阳兵等,2004;Yuan et al.,2016)。已有研究表明(邸欣月等,2015),石灰土淋溶程度增加,其土壤有机碳质量分数有降低趋势。本研究结果显示,两种典型石灰土土壤理化指标存在差异(图1),且差异性主要体现在表层上,红色石灰土淋溶程度强于黄色石灰土,可能是造成其土壤有机碳质量分数低于后者的原因。靳振江等(2014)研究典型岩溶土壤微生物丰度对碳循环的意义,指出微生物较高的丰度和功能活性可加快碳循环并促进土壤有机碳积累。本研究发现,黄色石灰土的微生物丰度高于红色石灰土,土壤有机碳质量分数同样表现为黄色石灰土高于红色石灰土,说明黄色石灰土较高的微生物丰度可促进其土壤有机碳积累,且由于微生物丰度差异主要体现在表层,故不同土层土壤有机碳存在差异。
本研究区土层对土壤养分的影响与大部分地区一致,均表现为土壤有机碳质量分数、速效磷质量分数以及全氮质量分数等随着土层的加深而下降,这是由于受上层土壤植被覆盖、植物残枝落叶以及根系影响,表层土壤有机质质量分数较高且碳、氮、磷等营养元素丰富(王静娅等,2014)。随着土层的加深,土壤容重加大,孔隙度减小,养分含量也持续下降,如不经人为扰动,表层营养元素较难向下迁移(宋贤冲等,2015)。本研究样地受人为干扰较弱,因此,土壤养分也表现出类似的变化趋势。
3.2 影响岩溶石灰土土壤微生物丰度特征的因素分析
土壤微生物大多为异养型,因此其在土壤中的主要限制因素仍为土壤养分。相关性分析(表3)与RDA分析(表4)结果均显示,土壤有机碳、速效磷和全氮与细菌、真菌丰度呈极显著正相关。双因素方差分析结果表明,相比土壤类型,土层深度对细菌与真菌丰度的影响更大(表2)。然而,土层深度与土壤类型对微生物丰度的影响也是由于土壤养分的限制。本研究发现,土层养分分布趋势与细菌丰度基本一致,两种石灰土养分差异显著性时,其细菌丰度也差异显著。说明土壤类型与土层对微生物丰度的影响主要由各土壤因子造成。正因为土层间土壤因子的差异强于土壤类型,故微生物丰度对土层的响应强于土壤类型。此外,细菌丰度与土壤有机碳、速效磷及全氮的相关性强于真菌丰度,说明细菌对土壤因子的反应更为敏感。Koide et al.(2005)指出竞争是构建真菌群落的重要因素。细菌丰度随土层加深而减少时,红色石灰土真菌丰度甚至出现增加的趋势,并且真菌可利用的有机物种类高于细菌(李宜浓等,2016),二者可能共同造成真菌对土壤因子的敏感度弱于细菌。
相关性分析结果显示,不仅土壤因子与微生物丰度存在相关性,微生物之间即细菌丰度与真菌丰度在0.01水平呈极显著正相关。这是因为土壤真菌对植物残渣的分解能力极强,可以将纤维素、半纤维素、蛋白质类化合物、木质素等以及其他类似化合物分解成较简单的物质,而土壤细菌参与较简单物质的分解(王少昆等,2009),因此真菌分解枯枝落叶产生较简单物质时可促进细菌的生长,使二者呈极显著正相关。真菌对细菌的这种促进作用也可能是造成细菌与土层和土壤类型呈显著正相关的重要原因。也有学者指出,土壤有机碳、速效磷和全氮与细菌、真菌丰度存在不同程度的显著正相关,说明土壤微生物不但参与土壤中 C、N、P循环(林巧燕,2011),而且还通过细菌与真菌间的协同作用参与土壤养分循环。
3.3 岩溶石灰土土壤微生物丰度的指示意义
黄色石灰土土壤有机碳、速效磷及全氮的质量分数均普遍高于红色石灰土,说明黄色石灰土养分含量高于红色石灰土。表层土即腐殖质堆积层是对环境和人类活动变化做出响应的第一个矿化层(Bagherzadeh et al.,2008),而微生物又是以上指标中对环境及人类活动反应较为敏感的指标(焦海华等,2016)。贺凤鹏等(2016)用表层(0~10 cm)与10~20 cm土层土壤微生物学特征的差异性评价草地退化程度,其差异性越小说明退化程度越严重。本研究发现,黄色石灰土表层的细菌丰度与20 cm以下土层差异性显著,红色石灰土表层土的细菌丰度与40 cm以下土层差异显著,说明红色石灰土较黄色石灰土贫瘠,此结果与养分指标结果一致。冯书珍等(2015)对岩溶原生林不同坡位、土层细菌丰度进行差异性分析,结果显示下坡位表层(0~10 cm)土与其他土层细菌丰度差异最显著,且其土壤养分含量最高。靳振江等(2014)对典型岩溶洼地、坡地和垭口不同土层细菌丰度进行差异性分析,结果显示,坡地表层(0~10 cm)土与其他土层细菌丰度差异最小,其土壤养分含量亦为最低。由于细菌丰度对土壤因子的敏感性强于真菌丰度,故细菌更适合用于表征土壤质量,表层土细菌丰度与其下土层差异越小,说明土地退化或营养贫瘠更为严重。
土壤细菌不仅具有指示土壤质量的作用,而且还因其几乎参与了土壤全部的生物化学过程而在石灰土演化过程中起到推进作用。有学者认为,土壤铁还菌希瓦氏菌对绿脱石进行还原之后,其分泌的电子中介体可促使其中的蒙脱石向伊利石转化(Liu et al.,2011;Liu et al.,2012),因而碳酸盐岩红土中的主要粘土矿物即伊利石的形成可能与微生物有关(李强等,2016)。此外,铁、锰在自然界中可发生细菌介导的氧化、还原(秦松岩等,2008)反应,而土壤中锰氧化物被还原的过程往往伴随着不同类型次生矿物的形成,包括蓝铁矿(Fe3(PO4)2)、菱铁矿(FeCO3)等 Fe(Ⅱ)铁矿物、针铁矿等 Fe(Ⅲ)矿物和磁铁矿(Fe3O4)等Fe(Ⅱ)-Fe(Ⅲ)混合的铁矿物(Borch et al.,2010),进而成为碳酸盐岩红色风化壳中的氧化铁矿物(朱立军等,2001)。尽管石灰土来源 于碳酸盐岩母质,但是在气候、生物、地形、时间综合作用下,特别在细菌的作用下土壤呈现不同的颜色,因此细菌在岩溶区黄色石灰土发育成红色石灰土过程中可能起着关键作用,这将是今后研究的重点。此外,植被凋落物的分解是植被归还土壤养分的主要方式,是生态系统内部物质循环与能量流动的重要环节。细菌与真菌作为土壤中物质循环与能量流动的推动者可通过分解凋落物起到养分保蓄作用,尤其在0~40 cm土层。本研究发现,两种石灰土细菌丰度与真菌丰度基本表现为表层土与40 cm以下土层差异显著,土壤有机碳质量分数也表现为表层土与40 cm以下土层差异显著,二者呈极显著正相关,由此说明较高的细菌与真菌丰度有助于促进土壤有机碳积累。
4 结论
(1)两种石灰土的微生物丰度受土壤有机碳、全氮及速效磷等土壤因子影响显著,土层间土壤因子差异性强于土壤类型,故微生物丰度表现出对土层的响应强于土壤类型。相关性分析结果表明,细菌丰度对土层养分变化的敏感度高于真菌,说明细菌丰度在土层间的差异度更适合表征土壤质量。
(2)细菌在真菌的协同作用下与土层和土壤类型的相关性更强。细菌在养分循环及保蓄方面起了更重要的作用。
致谢:感谢中国地质科学院岩溶地质研究所张美良研究员提供石灰土地质背景资料及鉴定。
参考文献:
BAGHERZADEH A, BRUMME R, BEESE F. 2008. Temperature dependence of nitrogen mineralization and microbial status in OH horizon of a temperate forest ecosystem [J]. Journal of Forestry Research, 19(1): 37-43.
BORCH T, KRETZSCHMAR R, KAPPLER A, et al. 2010. Biogeochemical redox processes and their impact on contaminant dynamics [J].Environmental Science & Technology, 44(1): 15-23.
BREMNER J M. 1965. Total nitrogen [M]//Black C A. Methods of Soil Analysis. Madison: American Society of Agronomy: 1149-1178.
COLWELL J D. 1963. The estimation of the phosphorus fertilizer requirements of wheat in southern New South Wales by soil analysis[J]. Australian Journal of Experimental Agriculture and Animal Husbandry, 3(10): 190-197.
KOIDE R T, XU B, SHARDA J, et al. 2005. Evidence of species interactions within an ectomycorrhizal fungal community [J]. New Phytologist, 165(1): 305-316.
LI C G, LI X M, KONG W D, et al. 2010. Effect of monoculture soybean on soil microbial community in the Northeast China [J]. Plant and Soil,330(1-2): 423-433.
LIU D, DONG H, BISHOP M E, et al. 2011. Reduction of structural Fe(III)in nontronite by methanogen Methanosarcina barkeri [J]. Geochim Cosmochim Acta, 75(4): 1057-1071.
LIU D, DONG H, BISHOP M E, et al. 2012. Microbial reduction of structural iron in interstratified illite-smectite minerals by a sulfate-reducing bacterium [J]. Geobiology, 10(2): 150-162.
MANERKAR M A, SEENA S, BÄRLOCHER F. 2008. Q-RT-PCR for assessing archaea, bacteria, and fungi during leaf decomposition in a stream [J]. Microbial Ecology, 56(3): 467-473.
XIE L W, DENG Y L, WANG B, et al. 2015. Changes in soil microbial biomasses and their quotients over the succession of karst rocky desertification [C]//International Conference on Advances in Energy and Environmental Science.
YUAN H, XIE H, LUO L, et al. 2016. Genetic characteristics and classification of limestone soil in south China [J]. Chinese Agricultural Science Bulletin, 32(21): 124-128.
鲍士旦. 2000. 土壤农化分析[M]. 北京: 中国农业出版社.
曹建华, 袁道先, 潘根兴. 2003. 岩溶生态系统中的土壤[J]. 地球科学进展, 18(1): 37-44.
邓艳, 蒋忠诚, 覃星铭, 等. 2009. 岩溶生态系统中不同植被枯落物对土壤理化性质的影响及岩溶效应[J]. 生态学报, 29(6): 3307-3315.
邸欣月, 安显金, 董慧, 等. 2015. 贵州喀斯特地区域土壤有机质的分布与演化特征[J]. 地球与环境, 43(6): 697-708.
冯书珍, 苏以荣, 张伟, 等. 2015. 坡位与土层对喀斯特原生林土壤微生物生物量与丰度的影响[J]. 环境科学, 36(10): 3832-3838.
葛云辉, 苏以荣, 邹冬生, 等. 2012. 桂西北石灰土土壤有机碳矿化对外源有机物质和碳酸钙的响应[J]. 生态学杂志, 31(11):2748-2754.
贺凤鹏, 曾文静, 王曌迪, 等. 2016. 温带草原退化对土壤剖面微生物学特征的影响[J]. 微生物学通报, 43(3): 702-711.
贺纪正, 李晶, 郑袁明. 2013. 土壤生态系统微生物多样性-稳定性关系的思考[J]. 生物多样性, 21(4): 412-421.
焦海华, 张淑珍, 景旭东, 等. 2016. 油田区多环芳烃污染盐碱土壤活性微生物群落结构解析[J]. 生态学报, 36(21): 6994-7005.
靳振江, 汤华峰, 李敏, 等. 2014. 典型岩溶土壤微生物丰度与多样性及其对碳循环的指示意义[J]. 环境科学, 35(11): 4284-4290.
蓝家程, 傅瓦利, 甄晓君, 等. 2011. 岩溶山区土壤性质及其对土下溶蚀速率的影响[J]. 水土保持学报, 25(2): 58-62.
李强, 靳振江. 2016. 岩溶生物地球化学研究的进展与问题[J]. 中国岩溶, 35(4): 349-356.
李瑞, 刘旻霞, 张灿, 等. 2017. 甘南亚高寒草甸不同坡向土壤微生物群落分布特征[J]. 生态环境学报, 26(11): 1884-1891.
李阳兵, 王世杰, 李瑞玲. 2004. 岩溶生态系统的土壤[J]. 生态环境,13(3):434-438.
李宜浓, 周晓梅, 张乃莉, 等. 2016. 陆地生态系统混合凋落物分解研究进展[J]. 生态学报, 36(16): 4977-4987.
李韵诗, 冯冲凌, 吴晓芙, 等. 2015. 重金属污染土壤植物修复中的微生物功能研究进展[J]. 生态学报, 35(20): 6881-6890.
梁月明, 苏以荣, 何寻阳, 等. 2017. 岩性对喀斯特灌丛土壤固氮菌与丛枝菌根真菌群落结构及丰度的影响[J]. 环境科学, 38(3): 1253-1261.
林巧燕. 2011. 增温和放牧对青藏高原高寒草甸土壤微生物结构和功能影响的研究[D]. 中国科学院研究生院.
刘丽, 徐明恺, 汪思龙, 等. 2013. 杉木人工林土壤质量演变过程中土壤微生物群落结构变化[J]. 生态学报, 33(15): 4692-4706.
秦松岩, 马放, 黄鹏. 2008. 自然水体中铁-锰氧化细菌的研究[J]. 环境科学, 29(6): 1649-1654.
盛浩, 宋迪思, 王翠红, 等. 2015. 土壤溶解性有机碳四种测定方法的对比和转换[J]. 土壤, 47(6): 1049-1053.
宋贤冲, 曹继钊, 唐健, 等. 2015. 猫儿山常绿阔叶林不同土层土壤微生物群落功能多样性[J]. 生态科学, 34(6): 93-99.
王静娅, 张凤华. 2014. 干旱区典型盐生植被群落土壤养分特征[J]. 水土保持学报, 28(5): 235-241.
王少昆, 赵学勇, 左小安, 等. 2009. 科尔沁沙质草甸土壤微生物数量的垂直分布及季节动态[J]. 干旱区地理, 32(4): 610-615.
杨柳, 何腾兵, 舒英格, 等. 2011. 贵州喀斯特区草地生态条件下石灰(岩)土的发生特性及系统分类研究[J]. 中国岩溶, 30(1): 93-99.
张美良, 邓自强. 1994. 我国南方喀斯特地区的土壤及其形成[J]. 贵州工学院学报, 23(1): 67-75.
周卫红, 张静静, 邹萌萌, 等. 2017. 土壤重金属有效态含量检测与监测现状、问题及展望[J]. 中国生态农业学报, 25(4): 605-615.
朱立军, 李景阳. 2001. 碳酸盐岩红色风化壳中的氧化铁矿物[J]. 地质科学, (4): 395-401, 515.
朱新玉, 胡云川, 芦杰. 2014. 豫东黄河故道湿地土壤生物学性状及土壤质量评价[J]. 水土保持学报, 21(2): 27-32.
猜你喜欢
宝藏(2022年11期)2022-03-07 08:57:48
海洋石油(2021年3期)2021-11-05 07:43:10
河北环境工程学院学报(2021年1期)2021-03-19 08:43:00
四川蚕业(2021年4期)2021-03-08 02:59:54
浙江农业学报(2016年7期)2016-06-15 20:30:08
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:16
现代冶金(2016年6期)2016-02-28 20:53:12
现代农业(2016年4期)2016-02-28 18:42:07
人间(2015年11期)2016-01-09 13:12:58
环境科技(2015年2期)2015-11-08 12:11:30
