
小陇山国家级自然保护区水楸种群结构和空间分布格局分析
2018-04-09 05:47赵欣鑫巨天珍卢秀娟谢顺涛张宋智
江苏农业科学 2018年5期
赵欣鑫, 巨天珍, 王 爽, 卢秀娟, 王 菁, 谢顺涛, 张宋智
(1.西北师范大学,甘肃兰州 730070; 2.天水小陇山林业科学研究所,甘肃天水 741022)
种群是物种存在和生物群落组成的基本单位[1-3]。种群结构可在一定程度上反映出种群个体在空间上的组配方式和发展趋势,是植物种群的重要属性之一[2];种群空间分布格局是指种群个体在水平空间的分布状态或配置状况,其既可对种群的水平结构进行定量描述,也可阐明种群与环境的互作过程[4]。研究种群结构和空间分布格局,有助于了解种群的生物学特征,利于预测群落的演替规律及发展方向,对种群合理保护、恢复及利用具有重要意义[5-7]。
水楸(FraxinusplatypodaOliv.)为木犀科白蜡属乔木,主要分布于我国东北长白山、小兴安岭、华北阔叶林地区。其喜生于湿润肥沃的土壤,树皮灰白色,鳞状裂,种子繁殖或萌芽更新繁殖,树体高大,材质优良,纹理细密,韧性大,广泛使用于室内装修、机械制造、家具、造船等方面[8]。该树种在甘肃小陇山林区的绝大多数林场均有分布,生长于海拔 1 600 m 以上的沟谷,是林区的主要成林树种之一。目前,国内学者对于林木种群结构与空间分布格局已有较多研究,如李荣等研究了天山北坡天山云杉的种群结构特征和空间分布格局,认为天山北坡云杉种群处于生命力旺盛的中龄期,格局呈聚集分布[5];潘霞等对百山祖北坡常绿阔叶林多脉青冈种群结构和分布格局进行了研究,认为多脉青冈种群结构属于稳定型,分布格局主要呈聚集分布[9];董秋莲等研究了甘肃省张掖市龙首山红砂种群结构和分布格局,认为红砂种群自然更新状况良好,格局为聚集分布[10]。但是,我国关于水楸种群的研究目前主要集中在水楸的繁育和造林技术等方面[11-13],有关小陇山水楸种群结构及分布格局方面的研究还未见报道。鉴于此,在甘肃省小陇山水楸林野外调查的基础上,从径级结构、静态生命表、存活曲线、生存函数曲线和空间分布格局等方面探讨水楸种群结构特征和分布格局的特点,并分析水楸种群动态和分布格局的相互关系,旨在为小陇山林区水楸种群的稳定性维持机制及可持续经营等提供理论依据。
1 研究区概况
甘肃小陇山林区为国家级自然保护区,位于105°30′~106°30′E、34°0′~34°40′N的秦岭西段,东西长212.5 km,南北宽146.5 km,海拔700~2 500 m,相对高差500~1 100 m,总面积31 938 hm2[14]。地处我国暖温带南缘与北亚热带的过渡地带,兼有我国南北方气候特点,气候温暖湿润,年均气温7~12 ℃,年降水量460~800 mm,年蒸发量989~1 658 mm,年日照时数1 520~2 313 h,相对湿度68%~78%,无霜期120~218 d,干燥度0.89~1.29,属湿润和半湿润类型区[15]。研究区内土壤大多为褐土和棕壤,土层厚度30~60 cm,较湿润,有机质及矿物质含量高,pH值6.5~7.5[16]。小陇山保护区现有林地面积28 723.2 m2,森林覆盖率高达80%,植被带谱完整,植物资源丰富,包括5个植被型组11个植被型53个群系,有草本植物1 900多种,木本植物800多种(不包括引种栽培),其中乔木层312种,灌木层512种,木质藤本层85种,以温带科属为主,热带、亚热带属种相对较少[14-15]。
2 研究方法
2.1 样地调查
基于实地勘察,针对林木不同生境、群落类型,在小陇山调查区选取5个20 m×20 m样方作为典型调查样地,采用典型样方法进行野外调查。在每个样地内采用每木调查法,记录所有个体的种名、树高、胸径、冠幅、树枝下高和各树种的密度、盖度等;利用GPS、海拔表、罗盘分别测定样方的地理坐标、海拔高度、坡向、坡度、坡位等生境因子(图1、表1)。
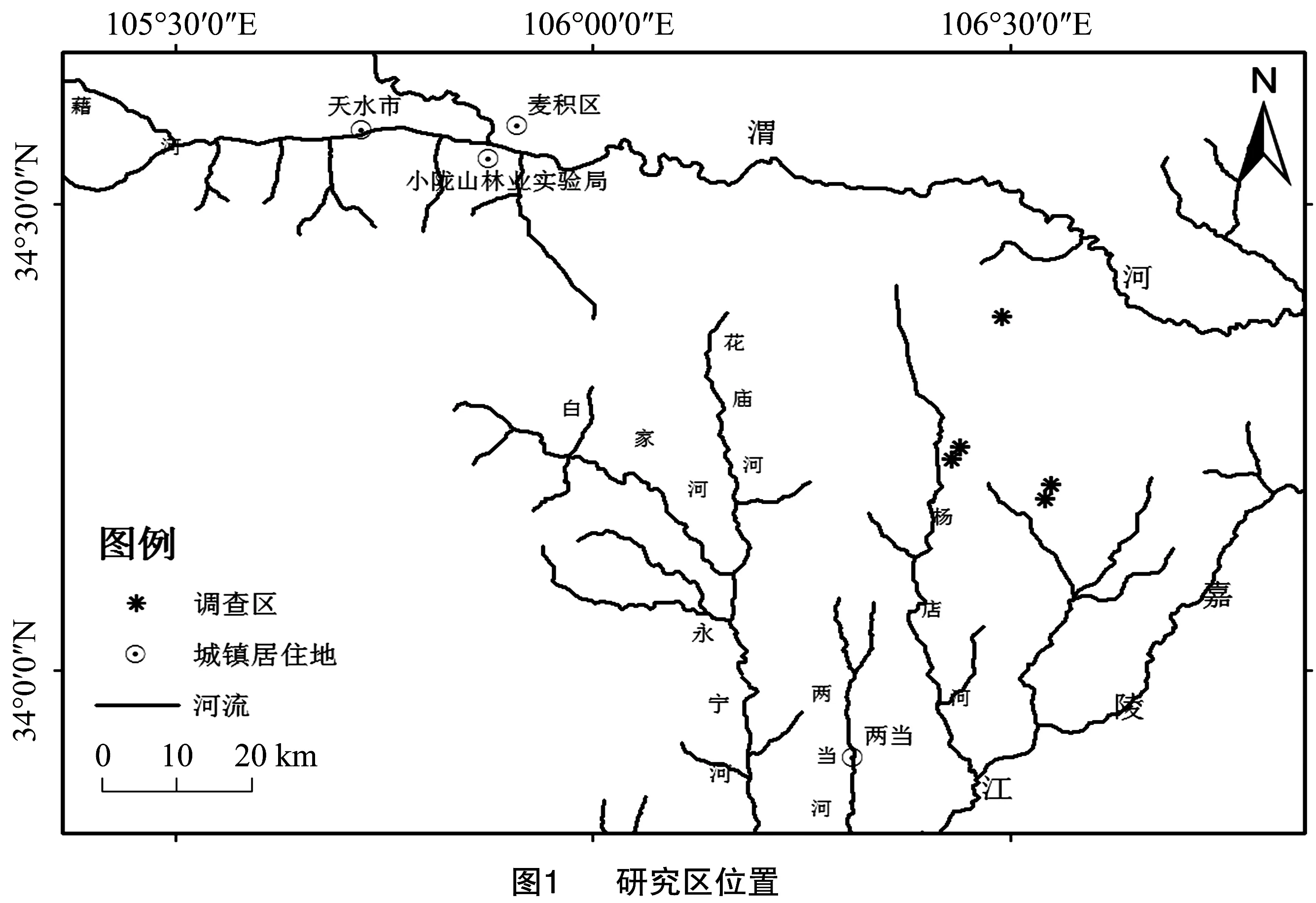
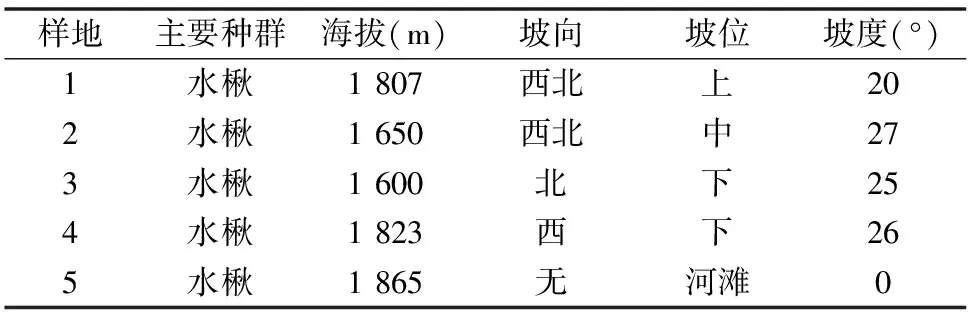
表1 调查区的群落环境
2.2 径级划分
采用径级结构代替年龄结构的方法分析种群动态特征[17-18]。依据调查数据,结合水楸种群生长规律和生活史特点,按胸径(diameter at breast height,DBH)每增加4 cm为1个径级,将所有水楸个体分为11个径级:D<4 cm为Ⅰ级,4 cm≤D<8 cm为Ⅱ级,8 cm≤D<12 cm为Ⅲ级,12 cm≤D<16 cm为Ⅳ级,16 cm≤D<20 cm为Ⅴ级,20 cm≤D<24 cm 为Ⅵ级,24 cm≤D<28 cm为Ⅶ级,28 cm≤D<32 cm为Ⅷ级,32 cm≤D<36 cm为Ⅸ级,36 cm≤D<40 cm为Ⅹ级,D≧40 cm为Ⅺ级(上限排除法)[17]。参照水楸的径级将其种群划分为幼苗幼树(Ⅰ级)、小树(Ⅱ级)、中树(Ⅲ-Ⅵ级)、大树(Ⅶ级-Ⅺ级)4个生长阶段[18]。
2.3 数据处理
2.3.1生命表编制根据基本数据,编制种群静态生命表,并绘制存活曲线,进而分析其种群动态变化[19]。由于在静态生命表的编制中,有时会出现死亡率为负的情况,故通常采取分段匀滑技术进行处理[20]。水楸种群静态生命表各指标计算公式[21]如下:
lx=ax/a0×1 000;
(1)
dx=lx-lx+1;
(2)
qx=dx/lx×100%;
(3)
Lx=(lx+lx+1)/2;
(4)
Tx=∑Lx;
(5)
ex=Tx/lx;
(6)
Kx=lnlx-lnlx+1。
(7)
式中:ax为x龄级内的现有存活数;lx是在x龄级开始时的标准化存活数;dx是从x~x+1龄级间隔期内标准化死亡数;qx是从x~x+1龄级间隔期间死亡率;Lx是从x~x+1龄级间隔期间还存活的区间寿命或个体数;Tx是从x龄级到超过x龄级的个体总数或总寿命;ex是进入x龄级个体的生命期望或平均寿命期望;Kx是为消失率(损失度)。
2.3.2生存分析方法为了更好地分析小陇山水楸种群结构,引入生存率函数(Sx)、积累死亡率函数(Fx)、死亡密度函数(fx)和危险率函数(λx)等4个生存分析函数于研究中,具体计算公式[21]如下:
Sx=P1P2…Px;
(8)
Fx=1-Sx;
(9)
fx=(Sx-1-Sx)/hx=Sx-1×qx/hx;
(10)
(11)
式中:x为龄级;Px为存活率,Px=1-qx;qx为死亡频率;hx为区间长度。
2.3.3空间分布格局主要选用以下5个基础指标,从不同的角度度量小陇山水楸种群的聚集特性,综合分析水楸种群的空间分布格局[14,22]:(1)扩散系数(C)。
(12)

(2)负二项分布参数(k)。此值与种群密度无关,用于判别种群中植株的聚集程度。
(13)
式中:当k<0时,为均匀分布;k>0时,为聚集分布;k值越接近于0,则聚集度越大;k值趋于无穷大(一般为8以上),为随机分布。
(3)聚集度指数(I)。
(14)
式中:当I<0时,为均匀分布;I=0时,为随机分布;I>0时,为聚集分布。
(4)Cassie指数。
(15)
式中:当Ca<0时,为均匀分布;Ca=0时,为随机分布;Ca>0时,为聚集分布。
(5)聚块性指标(PAI)。该值不受空样地的影响。
(16)
(17)
式中:当PAI<1时,为均匀分布;PAI=1时,为随机分布;PAI>1时,为聚集分布。
3 结果与分析
3.1 径级结构分析
由小陇山水楸种群径级结构图(图2)可以看出,水楸的径级结构接近倒“J”形,属于增长型结构[14]。从水楸径级分布来看,水楸种群在Ⅰ级分布最多,个体数为76株,Ⅱ级次之,为44株,在Ⅸ级分布最少,仅有5株;从水楸生长阶段来看,幼苗幼树(Ⅰ级)比例最高,占水楸总株数的33.78%,中树(Ⅲ~Ⅵ级)次之,占总株数的24%,小树(Ⅱ级)和大树(Ⅶ~Ⅺ级)比例相对较低。由此可见,水楸种群幼苗幼树充足,利于水楸进行种群的更新,但大树相对较缺乏,这可能与自然竞争、灾害、生理衰退、人为砍伐等因素有关。总体上来看,小陇山水楸种群径级结构完整,目前群落处于稳定发展中。

3.2 生命表分析
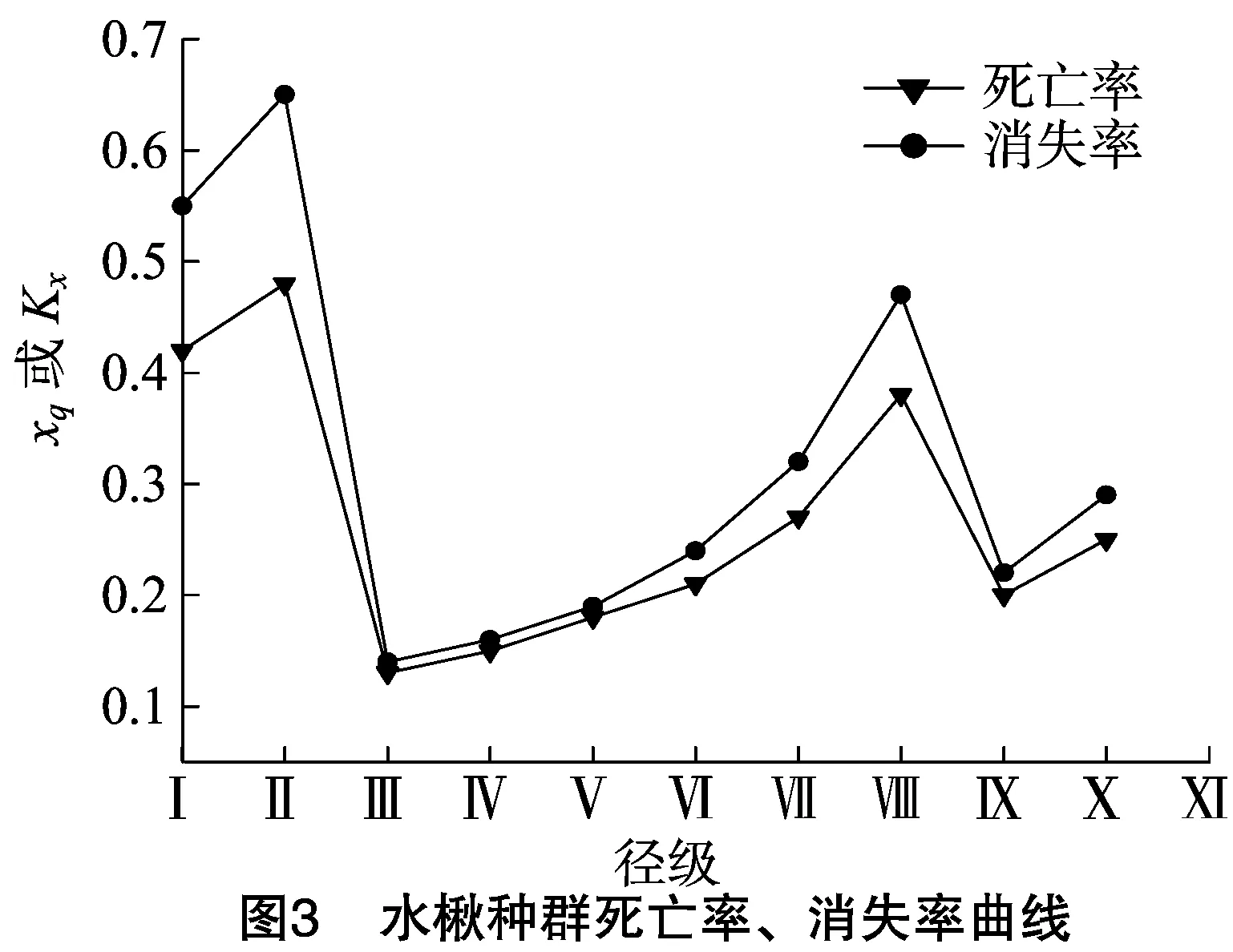
采用匀滑技术对调查数据进行处理,经修正后得到ax*,据此编制出小陇山水楸种群的静态生命表(表2),并绘制出水楸种群的死亡率、消失率曲线(图3)和存活曲线(图4)。由图3可知,水楸种群的死亡率、消失率变化趋势基本一致,并具有明显波动变化。其中死亡率在Ⅱ级最高,为48%;Ⅰ级次之,为42%;Ⅲ级最低,为13%;之后随着个体生长对资源需求的逐渐加大,致使其死亡率因种内竞争的不断加剧而随之增高,并于Ⅷ级再次达到峰值,为38%。随后水楸死亡率因自疏过程而在Ⅸ级有所降低,最后因生物学特征,种群个体达到其生理年龄,其死亡率再次升高。水楸种群存活曲线采用2种数学模型进行检验,即指数函数(Nx=N0e-bx)用以描述Deevey-Ⅱ型存活曲线,幂函数(Nx=N0x-b)用以描述Deevey-Ⅲ型存活曲线[23]。结果显示,Nx=7.184e-0.058x(r2=0.977 5,P<0.01),Nx=7.486x-0.246(r2=0.876 2,P<0.01)。2种数学模型中指数函数拟合方程相关系数更高, 故水楸种群的存活曲线更趋于Deevey-Ⅱ型(图4)。由图4可以看出,水楸种群各径级的死亡率基本相同,局部稍有波动,在幼龄期死亡率较高,但仍可保证一定数量以确保种群稳定地更新和发展,整体表明小陇山水楸种群目前处于稳定发展状态。

表2 小陇山水楸种群的静态生命
注:ax为存活数;ax*为匀滑后存活数;lx为标准化存活数;dx为死亡数;qx为死亡率;Lx为区间寿命;Tx为总寿命;ex为期望寿命;Kx为消失率。

3.3 生存分析
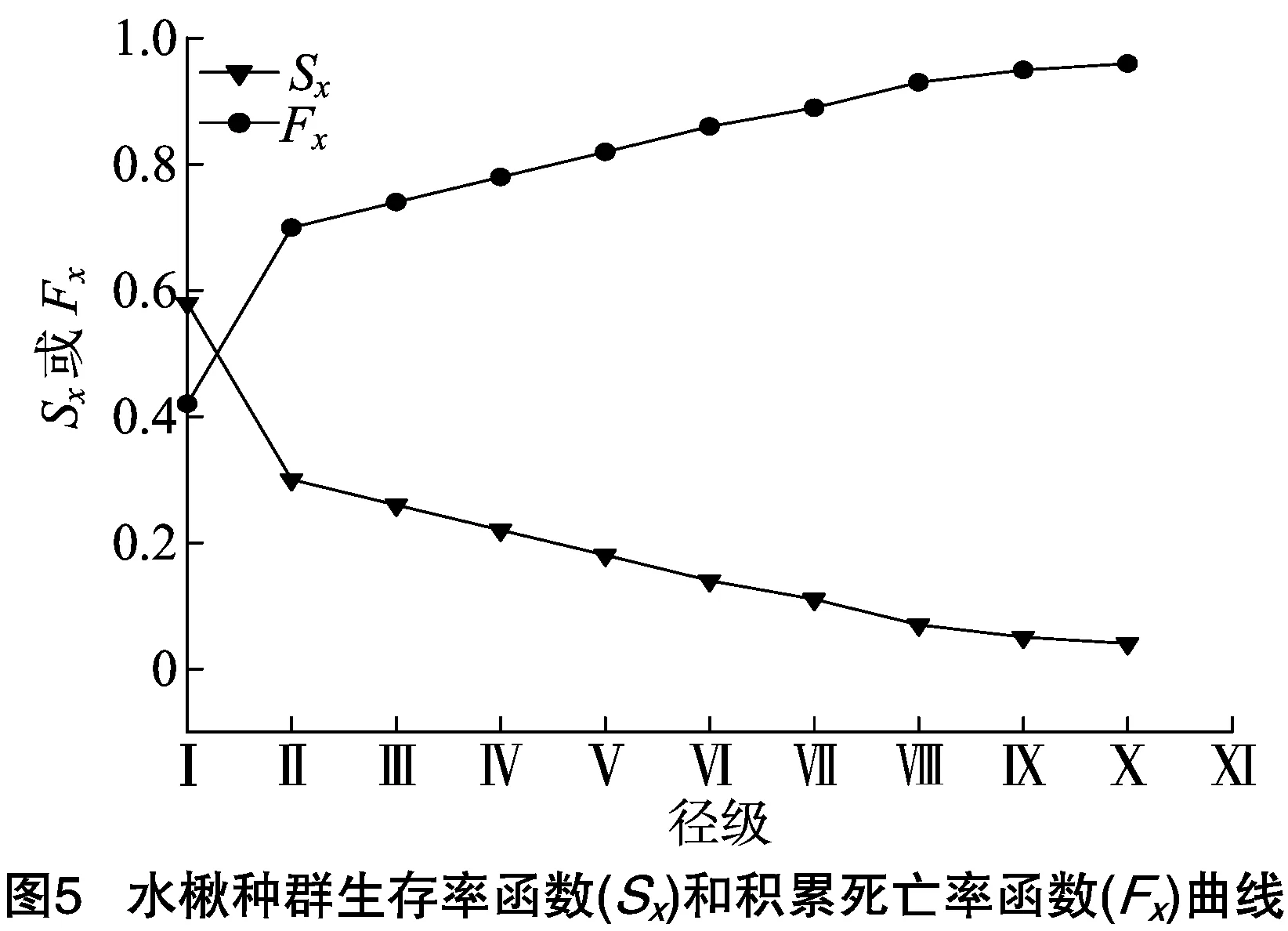
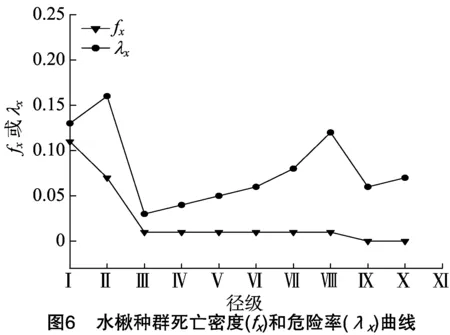
基于水楸种群4个生存函数估计值,分别绘制水楸种群生存率(Sx)、积累死亡率函数(Fx)曲线图(图5)和死亡密度(fx)、危险率(λx)曲线图(图6)。由图5可知,水楸种群生存率基本呈现单调递减趋势,积累死亡率基本趋于单调递增,两者互补,在Ⅰ~Ⅲ级变化幅度较大,后期变化相对平缓,并于Ⅹ级达到生理年龄,生存率低至5%,说明水楸种群在幼龄期死亡率较高,老龄树稀少。由图6可以看出,水楸种群的死亡密度曲线和危险率曲线变化趋势差别较明显,水楸林中幼苗幼树(Ⅰ级)、小树(Ⅱ级)死亡密度较高,大树(Ⅶ~Ⅸ级)相对较低且基本持平,均在1%以下,而水楸种群危险率在小树(Ⅱ级)最高,为16%,幼苗幼树(Ⅰ级)次之,为13%。整体来看4个生存函数与种群存活曲线、死亡率曲线和消失率曲线分析结果一致,表明小陇山水楸种群具有前期增长,中期稳定,后期衰退的特点。
3.4 空间分布格局分析
典型样地的种群格局动态趋势可反映出整个水楸种群动态格局的大致趋势,文中采用5种不同聚集度指标对各样地分布格局的判定结果基本一致(表3)。由表3可知,各样地水楸种群均呈聚集分布,但聚集强度有所不同。根据负二项参数(k)和聚块性指标(PAI),可以看出,样地4的k值最小,为0.28,PAI值最大,为4.63,说明该样地的水楸种群聚集强度最大;其次为样地1,k值为0.32,PAI值为4.14;样地5的k值最大,为4.11,PAI值最小,为1.24,即该样地聚集度最小。各样地具体聚集度由大到小依次为:样地4>样地1>样地3>样地2>样地5。结果表明,小陇山水楸种群空间分布格局呈聚集分布。

表3 水楸种群分布格局
4 结论与讨论
小陇山水楸种群的径级结构、生命表、存活曲线、生存分析曲线等均是水楸种群生物学特性与环境因素共同作用的结果。从小陇山水楸种群径级结构可以看出,水楸种群在幼苗幼树(Ⅰ级)生长阶段比例最高,大树(Ⅶ~Ⅺ级)比例相对较低,其径级结构完整,总体分布接近倒“J”形,种群潜在的自然更新能力较强,群落处于稳定发展中。静态生命表显示,其存活曲线基本接近于Deevey-Ⅱ型,各径级的死亡率基本相同,局部稍有波动。死亡率和消失率曲线以及4个生存函数均呈现出水楸幼苗死亡率较高的特征,在幼苗幼树(Ⅰ级)死亡率为42%,表明林下水楸幼苗幼树很丰富,但较难长成小树,这可能由于单位面积承载力有限,随着水楸分株的生长发育,其对环境资源需求加剧,特别是水分的争夺加剧,从而造成自疏和它疏作用,致使个体大量死亡。此外,水楸种群死亡率在Ⅸ级后又再次升高,这是种群个体因生物学特征达到生理年龄所致。从小陇山水楸种群的动态演化趋势看,水楸种群幼苗丰富,大树较少,种群结构属于稳定型,这与很多研究结果类似。柳新红等研究了清凉峰自然保护区木荷的种群结构,得出木荷的种群结构特征与小陇山水楸相似,即幼树丰富,大树较少,总体上有足够的幼树,能够实现自我更新,属稳定型种群[24];潘霞等研究了百山祖北坡常绿阔叶林多脉青冈种群,得出其种群结构幼树期丰富,衰退期个体数少[9],在幼苗期死亡率较高等分析结果均与小陇山水楸种群基本一致。基于已有研究成果,小陇山国家级自然保护区水楸种群动态分析表明,水楸种群具有相对的稳定性,但林下更新层仍存在不足,难以维持其长期稳定,须对其进行人工促进天然更新,以实现小陇山林业的稳定性维持机制和可持续经营。
小陇山水楸种群空间分布格局的分析结果表明,水楸种群在总体上分布格局呈聚集分布,这是种群生存的策略或适应机制,更有利于其获取足够的生存资源,是物种与环境长期相互作用的结果[23]。王晓雨等研究了长白山北坡林线岳桦种群空间分布格局,认为岳桦种群总体空间分布格局呈聚集分布[25];刘贵峰等对天山云杉种群分布格局进行了研究,认为天山云杉种群格局整体上呈集群分布[26]。这些研究均与小陇山水楸种群研究结果一致,小陇山水楸种群的聚集分布格局,与其自身的环境因素和生物因素有关。水楸依靠种子或萌芽更新繁殖,这决定了水楸幼苗的生长多在母树周围。另外,格局呈聚集分布的种群,在环境中会比单独个体具有更大的竞争优势,可大大提高物种个体的存活机会,能更好地发挥出群体效应来维持自身的稳定增长状态[27]。因此,植物种群与环境的长期作用下形成的特定聚集分布格局[28],更有利于水楸的生长,也是小陇山水楸群落处于稳定增长期特有的种群分布形式。
研究表明,小陇山水楸种群径级结构完整,属于稳定增长型,其空间分布格局总体上呈聚集分布,群落处于稳定发展阶段。自然水楸林种群结构动态分析和空间分布格局研究,为小陇山林区水楸种群的维持机制、可持续经营以及生态恢复植被的有效保护和利用提供了依据。
参考文献:
[1]闫淑君,洪伟,吴承祯. 闽江口琅岐岛朴树种群结构与空间分布格局[J]. 生态学杂志,2011,30(2):221-226.
[2]李效雄,刘贤德,赵维俊. 祁连山大野口流域青海云杉种群结构和空间分布格局[J]. 干旱区地理,2012,35(6):960-967.
[3]杨翔,卢杰. 大花黄牡丹群落主要种群的生态位研究[J]. 江苏农业科学,2010(1):314-318.
[4]Dray S,Legendre P,Peres-Neto P R. Spatial modeling:a comprehensive framework for principal coordinate analysis of neighbor matrices(PCNM)[J]. Ecological Modelling,2006,196(3/4):483-493.
[5]李荣,罗惠文,宋于洋,等. 天山北坡天山云杉的种群结构特征和空间分布格局[J]. 干旱区资源与环境,2013,27(1):91-96.
[6]Masaki T,Ota T,Sugita H,et al. Structure and dynamics of tree populations within unsuccessful conifer plantations near the Shirakami Mountains,a snowy region of Japan[J]. Forest Ecology and Management,2004,194(1/2/3):389-401.
[7]张希彪,上官周平,王金成,等. 子午岭人工油松林群落更新特征及影响因子[J]. 山地学报,2014,32(5):561-567.
[8]安定国. 甘肃省小陇山高等植物志[M]. 兰州:甘肃民族出版社,2002:324-350.
[9]潘霞,周荣飞,顾莎莎,等. 百山祖北坡常绿阔叶林多脉青冈种群结构和分布格局[J]. 亚热带植物科学,2013,3(3):227-232.
[10]董秋莲,李毅,单立山,等. 张掖市龙首山红砂种群结构和分布格局研究[J]. 水土保持通报,2013,33(4):284-288.
[11]安长生. 小陇山林区水楸造林技术[J]. 林业实用技术,2010(12):17-18.
[12]王喜强,张江涛. 水楸实生苗培育技术[J]. 甘肃科技,2010,26(23):160-161.
[13]徐映霞. 小陇山林区珍贵用材树种水楸的繁育推广[J]. 农业科技与信息,2008(14):15-16.
[14]巨天珍,宋晓伟,师贺雄,等. 小陇山国家级自然保护区油松林优势种群空间分布格局分析[J]. 干旱区资源与环境,2012,26(9):62-66.
[15]刘文桢,赵中华,胡艳波,等. 小陇山栎类混交林经营[M]. 北京:中国林业出版社,2015:15-20.
[16]索安宁,巨天珍,张俊华,等. 甘肃小陇山锐齿栎群落生物多样性特征分析[J]. 西北植物学报,2004,24(10):1877-1881.
[17]张毓涛,李吉玫,常顺利,等. 天山中部天山云杉种群空间分布格局及其与地形因子的关系[J]. 应用生态学报,2011,22(11):2799-2806.
[18]何东进,肖石红,李晓景,等. 中国红树林北缘区秋茄树种群空间分布格局和结构特征分析[J]. 植物资源与环境学报,2014,23(2):41-47.
[19]吴俊侠,张希明,邓潮州,等. 塔里木河下游胡杨种群特征与动态分析[J]. 干旱区资源与环境,2010,24(8):164-169.
[20]江洪. 云杉种群生态学[M]. 北京:中国林业出版社,1992:7-26.
[21]巨天珍,米彩燕,师贺雄,等. 小陇山国家级自然保护区油松种群更新及生存[J]. 山地学报,2012,30(1):36-40.
[22]张雷,邱乾栋,臧德奎. 山东百部种群的空间分布格局[J]. 武汉植物学研究,2009,27(6):617-621.
[23]赵志刚,郭俊杰,曾杰,等. 广西大明山格木种群的空间分布格局与数量动态特征[J]. 林业科学,2014(10):1-7.
[24]柳新红,李因刚,沈国春,等. 清凉峰自然保护区木荷种群结构研究[J]. 林业科学研究,2011,24(1):28-32.
[25]王晓雨,于大炮,周莉,等. 长白山北坡林线岳桦种群空间分布格局[J]. 生态学报,2015,35(1):116-124.
[26]刘贵峰,丁易,臧润国,等. 天山云杉种群分布格局[J]. 应用生态学报,2011,22(1):9-13.
[27]苏日古嘎,张金屯,程佳佳,等. 东灵山糠椴林优势种种群结构与分布格局[J]. 生态学杂志,2009,28(7):1253-1258.
[28]吴文娟,王庆成,许丽娟. 帽儿山地区天然次生林中水曲柳种群格局分析[J]. 东北林业大学学报,2013(10):54-59.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
现代园艺(2018年2期)2018-03-15
甘肃林业(2016年4期)2016-11-07
甘肃林业(2016年4期)2016-11-07
甘肃林业(2016年6期)2016-02-27
西北植物学报(2015年9期)2015-07-05
