
ABA及氟啶酮调控下冬小麦分蘖节抗寒相关蛋白的鉴定与分析
2018-03-28 10:33:51包雨卓赵浡彤彭瞰看徐庆华王军虹
麦类作物学报 2018年2期
杨 宁,包雨卓,赵浡彤,吕 岩,彭瞰看,田 宇,徐庆华,于 晶,王军虹,苍 晶
(1.东北农业大学生命科学学院,黑龙江哈尔滨 150030; 2.广东白云学院,广东广州 510450)
低温胁迫是限制小麦产量的主要因素。ABA是重要的植物激素之一。研究证实,外源施加ABA能够促进植物体内ABA的合成和运输,增加渗透调节物质的含量,提高抗氧化酶活性,诱导抗寒相关基因表达以及促进相关酶的重新合成,从而提高植物抗寒性[1-2]。氟啶酮是ABA合成的抑制剂,通过抑制ABA前体物质类胡萝卜素的合成,从而抑制植物体内ABA的合成[3-4]。用氟啶酮作对照,可进一步明确哪些蛋白可能受到ABA的诱导。
双向电泳技术是蛋白质组学研究的关键技术,在植物抗逆相关研究中应用日趋广泛,对植物抗寒等抗性研究已取得了一些进展。Meza-Basso等[5]首次通过双向电泳技术在油菜中发现受低温诱导的蛋白,并在多种植物中得到应用。Hashimoto等[6]采用双向电泳在对低温胁迫下水稻幼苗的研究中发现,水稻叶鞘、叶片和根系在低温处理后出现39个蛋白差异点。Cui等[7]采用双向电泳对不同温度处理下水稻幼苗叶片蛋白进行分离,鉴定到41个低温诱导蛋白。Gao等[8]采用双向电泳在对盐芥莲座叶响应低温胁迫的蛋白研究中发现,所有参与RNA代谢、防御和蛋白代谢的蛋白质都显著上调。
东农冬麦1号是首例可在北方高寒地区安全越冬的强抗寒冬小麦品种,返青率大于85%。本课题组前期研究发现,东农冬麦1号分蘖节是其物质准备能力和抗寒代谢适应最强的器官,在冬小麦安全越冬中起重要作用[9]。鉴于此,本试验以东农冬麦1号为材料,于三叶期分别外源喷施10 μmol·L-1ABA和50 μmol·L-1氟啶酮,待大田自然降温至4 ℃和-25 ℃时取分蘖节,采用聚丙烯酰胺凝胶电泳对各处理组全蛋白进行分离,然后进一步采用双向电泳分离蛋白,筛选出与ABA调控相关或与低温应答相关的差异蛋白后进行质谱分析,探讨外源ABA对低温下冬小麦蛋白质组的影响,进而揭示冬小麦抗寒的蛋白质基础及其与ABA调控的关系。
1 材料与方法
1.1 试验材料种植及取样
供试冬小麦(TriticumaestivumL.)材料为东农冬麦1号,由东北农业大学小麦育种研究室提供。
选取籽粒饱满一致的冬小麦种子于东北农业大学试验田播种。完全区组设计,3次重复,小区行长1.5 m,10行区,行距0.2 m,播种量为450粒·m-2,播深5 cm,常规管理。待冬小麦生长至三叶期时,分别采用10 μmol·L-1ABA、50 μmol·L-1氟啶酮(二者浓度为前期试验确定的最适浓度)进行叶面喷施,用量为0.25 L·m-2,以喷施等量蒸馏水为对照。分别于10 d最低温平均4 ℃和-25 ℃时取样。各处理分别取50株长势相似的麦苗,将分蘖节剪成0.5 cm的小段,混匀后用液氮迅速冷冻,于-80 ℃冰箱保存备用。
1.2 全蛋白的提取、裂解及定量
全蛋白的提取参照Damerval等[10]的方法并略作改进。取1.0 g样品在液氮中充分研磨,转入50 mL离心管中。加入10 mL预冷的10% 三氯乙酸(TCA)/丙酮溶液,于-20 ℃冰箱放置45 min。4 ℃、16 000 r·min-1离心15 min,弃上清,重悬沉淀。加入10 mL预冷的10% TCA/丙酮溶液,于-20 ℃冰箱放置45 min。4 ℃、16 000 r·min-1离心15 min,弃上清,沉淀中加入预冷的80%丙酮,振荡混匀后于-20 ℃冰箱放置45 min(重复两次该步骤)。弃上清,沉淀低温冷冻干燥成粉末状。
在上述所得到的粉末状蛋白质样品中加入500 μL裂解液(7 mol·L-1尿素、2 mol·L-1硫脲、4% CHAPS、65 mol·L-1DTT、2% IPG buffer)混匀,涡旋振荡20 min,4 ℃过夜。取上清,用蛋白质定量试剂盒(2-D Quant-Kit,GE-Healthcare)定量。
1.3 蛋白双向电泳
根据蛋白质含量吸取一定体积的蛋白溶液,加适量水化液(7 mol·L-1尿素、2 mol·L-1硫脲、2% CHAPS、20 mmol·L-1DTT、0.5% IPG buffer、0.1%溴酚蓝)补齐至450 μL,加入胶条槽中,用24 cm IPG strip覆盖,通过Ettan IPGphor Ⅲ(GE-Healthcare)进行等电聚焦,聚焦程序设置为:主动水化30 V 10 h,200 V 1 h,500 V 1 h,1 000 V 1 h,4 000 V 1 h,8 000 V 1 h,64 000 Vh聚焦,500 V维持。
第一相等电聚焦结束后,将胶条取出置于放入10 mL胶条平衡缓冲液Ⅰ[6 mol·L-1尿素、75 mmol·L-1Tris-HCl(pH 8.8)、29.3%甘油、2% SDS、0.002%溴酚蓝、1% DTT]的水化盘中,平衡15 min。取出后再放入10 mL胶条平衡缓冲液Ⅱ[6 mol·L-1尿素、75 mmol·L-1Tris-HCl(pH 8.8)、29.3%甘油、2% SDS、0.002%溴酚蓝、2.5%碘乙酰胺]中平衡15 min。平衡完毕后,将胶条在电极缓冲液中浸泡2 s后转移到12.5%的SDS-PAGE凝胶上,低熔点琼脂糖封胶后,应用Ettan Dalt Six(GE-Healthcare)进行第二相SDS-PAGE电泳,20 ℃恒温下,1 W 30 min,13 W直至溴酚蓝移至凝胶底部,电泳停止。
1.4 染色与脱色
电泳结束后,将胶块取出置于20%TCA中固定15 min后,用染色液(450 mL蒸馏水、450 mL无水乙醇、100 mL冰乙酸、2.5 g考马斯亮蓝R250)染色2 h后,转至脱色液中(450 mL蒸馏水、450 mL无水乙醇、100 mL冰乙酸)中,不断更换脱色液直到条带清晰可见。
1.5 凝胶扫描及质谱分析
待凝胶背景清晰后,用Ettan Image Scanner扫描仪拍照后,将凝胶保存于含有10%冰乙酸的溶液中。利用ImageMasterTM2D Platinum 7.0软件(GE Healthcare)对图像进行分析,筛选出差异表达的蛋白点,选择相对体积改变大于2倍的蛋白点,借助SPSS进行分析,选择其中差异倍数在2倍以上的24个蛋白点切胶后进行质谱分析。
用含50%乙腈的25 mmol·L-1碳酸氢铵洗脱后乙腈脱水。56 ℃下用10 mmol·L-1DTT打开二硫键,然后暗处55 m mol·L-1IAM处理,进行半胱氨酸的烷基化封闭,最后用胰蛋白酶溶液裂解。裂解液用MALDI-TOF/TOF进行肽指纹图谱及二级质谱测定。质谱仪检测时设置参数为:检测模式为反射模式,谱图质量范围为500~3 500 Da,分辨率为50 000。默认每张一级谱图选择5个母离子进行二级鉴定。蛋白质胶点鉴定主要是通过试验产生的质谱数据,与数据库模拟得到的理论质谱数据进行匹配,从而得到蛋白质鉴定结果。
2 结果与分析
2.1 各处理组冬小麦分蘖节的双向电泳图谱
4 ℃时,各处理组双向电泳结果如图1。对图1各处理组进行软件分析,得到可匹配的蛋白点587个。ABA处理组与对照组相比,133个蛋白点差异表达变化量达到2倍以上,其中,115个蛋白点上调,18个蛋白点下调(图1a、图1b);氟啶酮处理组与对照组相比,59个蛋白点差异表达变化量达到2倍以上,其中,31个点上调,28个点下调(图1a、图1c);ABA处理组与氟啶酮处理组相比,差异表达变化量达到2倍以上的点有64个,其中36个点上调,28个点下调(图1b、图1c)。
25 ℃时,各处理组双向电泳结果如图2。对图2各处理组进行软件分析得到可以匹配的蛋白点394个。ABA处理组与对照组相比,61个蛋白点差异表达变化量达到2倍以上,其中,41个蛋白点上调,20个蛋白点下调(图2a、图2b);氟啶酮处理组与对照组相比,84个蛋白点差异表达变化量达到2倍以上,其中,29个点上调,55个点下调(图2a、图2c);ABA处理组与氟啶酮处理组相比,差异表达变化量达到2倍以上的点有65个,其中34个点上调,31个点下调(图2b、图2c)。
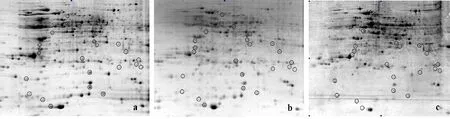
a:对照组;b:ABA处理组;c:氟啶酮处理组。图2同。
a:Control; b:ABA treatment; c:Fluridone treatment. The same in figure 2.
图14℃时各处理组的2-DE图谱
Fig.12-DEmapsofeachtreatmentat4℃
2.2 差异表达蛋白点的选择和质谱鉴定结果
对各处理之间差异倍数达到2倍以上的蛋白点进行筛选,选择在低温下变化倍数较大及在ABA处理组和氟啶酮处理组中变化相反的23个蛋白点,和低温下消失的蛋白点1个,共24个蛋白点(图3)。其中,21个(点1~14、点16~17、点19~23)在ABA处理组中上调,而在氟啶酮处理组下调;2个(点15、点18)在ABA处理组中下调,而在氟啶酮处理组上调;1个(点24)在4 ℃特异表达,-25 ℃消失的蛋白点。对以上筛选到的24个蛋白点进行质谱分析,均得到了完整的肽指纹图谱,Mascot数据库检索结果如表1示,其中15种为已知蛋白,9种未知蛋白。
由图4可知,鉴定出的蛋白与谱图匹配的肽段长度占整个蛋白长度40%~100%、35%~40%、30%~35%、25%~30%的分别有28%、20%、11%、11%,说明鉴定出的蛋白覆盖率较高,有较高的可信度。
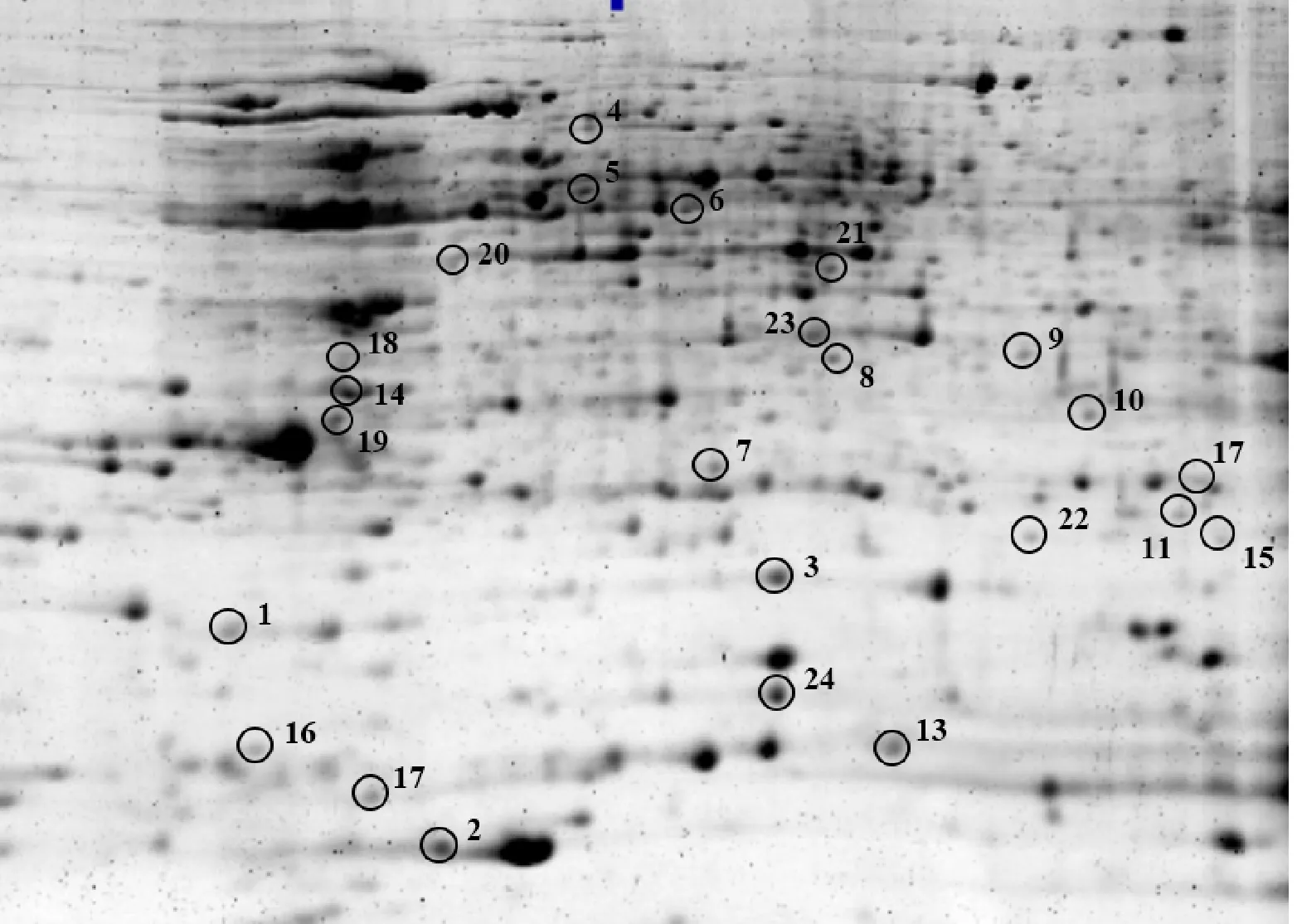
图3 4 ℃下进行质谱分析的蛋白点
2.3 差异蛋白的功能分析
已鉴定出的差异蛋白可分为逆境蛋白、代谢相关蛋白、转录翻译相关蛋白和未知蛋白。逆境蛋白包括低温诱导蛋白16(点1)、低温响应RNA结合蛋白(点2)、冷调节蛋白(点16、点17)。代谢相关蛋白包括磷酸甘油酸变位酶(点4)、磷酸甘油酸激酶(点23)、ATP合酶β亚基(点5)、谷胱甘肽S转移酶1(点7)、26S蛋白酶非ATP酶调节亚基(点10)、谷胱甘肽转移酶(点11)和谷胱甘肽转移酶F5(点15)。转录翻译相关蛋白包括新生多肽相关复合物β亚基(点3)、4A真核起始因子(点6)和真核翻译起始因子5A3(点24)。未知蛋白包括点8、9、12、13、14、18、19、20、21和22。
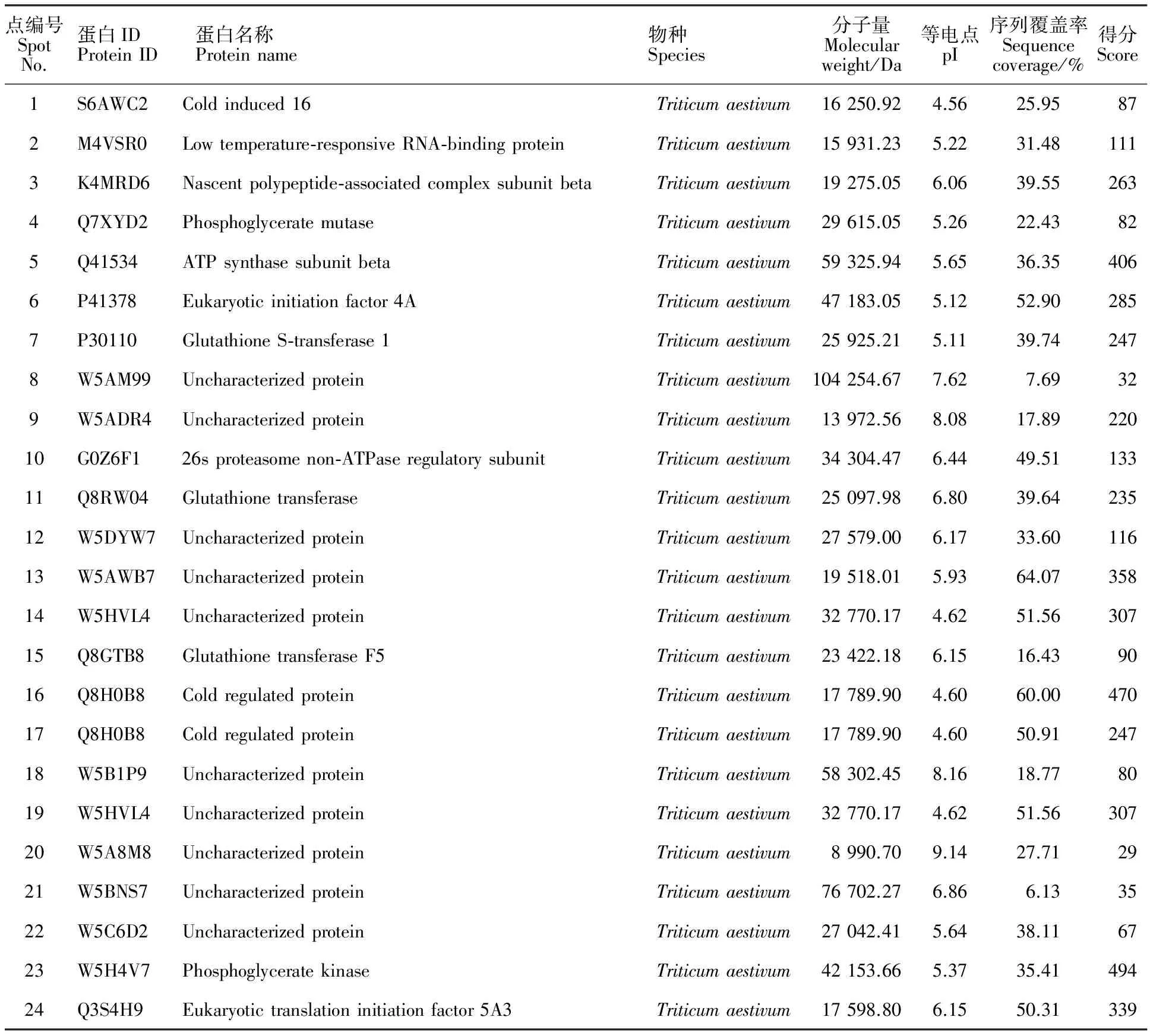
表1 质谱分析结果Table 1 Results of mass spectrometry
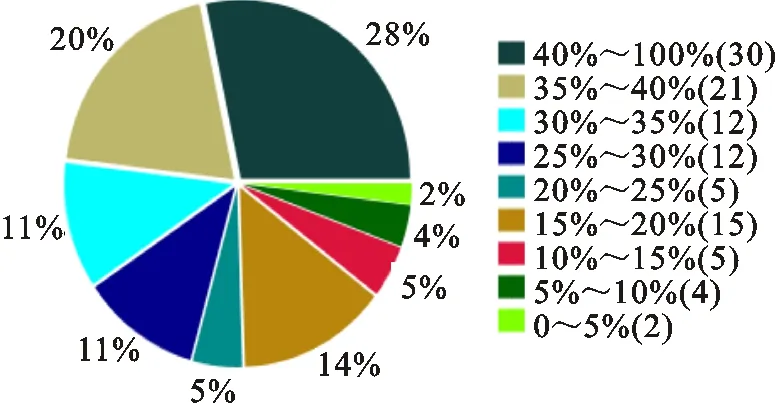
图例中,百分数表示蛋白鉴定覆盖率,括号内数字表示覆盖率范围内的蛋白种类数。
In the legend,the percentage indicates protein sequence coverage,and the data in the parenthesis indicates the number of protein species within that sequence coverage.
图4蛋白鉴定覆盖率分布
Fig.4Distributionofproteinsequencecoverage
3 讨 论
3.1 逆境蛋白
低温诱导蛋白16(cold induced 16, WCI16)(点1)是冬小麦在冷驯化期间诱导的一种蛋白,Sasaki K等[11]的研究从晚期胚胎发育蛋白(late embriogenesis abundant protein, LEA)通常具有的4个特征(超亲水性、沸腾溶解度、应力诱导性和应激保护性)证明了WCI16可能代表一类新的LEA蛋白。植物LEA蛋白被认为是一种逆境响应蛋白,低温、干旱及ABA等均可诱导其表达,其中ABA是LEA蛋白基因表达的重要调节因子,根据LEA蛋白基因表达是否依赖ABA可分为ABA依赖型和非ABA依赖型[12]。本试验中WCI16在ABA处理组与氟啶酮处理组表达量变化相反的特点,说明该蛋白可能是ABA依赖型LEA蛋白。
RNA结合蛋白(RNA-binding protein, Rbps)是基因表达转录后调控的关键蛋白[13],植物中编码该蛋白的基因在玉米、拟南芥、烟草和大麦等植物中均被克隆得到。研究表明,拟南芥中编码该蛋白的多个基因表达量在低温条件下显著增加[14],与本研究中低温响应RNA结合蛋白(low temperature-responsive RNA-binding protein, LTR-Rbps)在低温ABA处理下表达量上调的趋势一致,而在ABA处理组与氟啶酮处理组表达量相反的结果则表明,低温响应RNA结合蛋白受到外源ABA的诱导。
冷调节蛋白(cold regulated protein, CORPs)(点16、17)是大多数植物在低温驯化诱导条件下产生的一类特异性蛋白,研究发现,除低温外,外源ABA、盐胁迫、干旱、热激等逆境胁迫均能诱导CORPs产生[15]。本试验得到的COR蛋白WCOR18与之前报道过的大麦COR蛋白Tma-ap3具有高度同源性[16]。
3.2 代谢相关蛋白
磷酸甘油酸变位酶(phosphoglycerate mutase, PGAM)(点4)是糖酵解和糖异生途径中催化3-磷酸甘油酸和2-磷酸甘油酸相互转化的关键酶,一般分为辅因子依赖型PGAM(dPGAM)和辅因子非依赖型PGAM(iPGAM),其中,iPGAM广泛分布在植物中,并且在响应非生物胁迫中起重要作用。Amme等[17]在拟南芥的低温胁迫研究中发现,AtiPGAM1表达量显著升高,说明AtiPGAM1在植物遭受冷胁迫时起正调节作用。本试验中PAGM在低温和ABA处理下表达量上升,说明该酶能够响应植物的低温胁迫,并可能受到外源ABA的诱导。
磷酸甘油酸激酶(phosphoglycerate kinase, PGK)(点23)的主要功能是作为糖酵解途径的关键酶,催化1,3-二磷酸甘油酸转化成3-磷酸甘油酸的过程,并产生1分子ATP,逆反应生成1分子ADP[18]。PGK在非生物胁迫下表达量升高产生ATP以增强植物抗性,该结果在水稻[19]和豌豆[20]均已被证实。本研究中PGK在低温和ABA处理下表达量升高,说明PGK能够提高植物在低温胁迫下的抗性,一定量的ABA可能诱导PGK表达量升高从而增强植物抗逆性。
20S蛋白酶体能选择性降解氧化胁迫受损的蛋白,这一功能不依赖ATP和泛素[21]。本研究中得到的26S蛋白酶体非ATP酶调节亚基(26S proteasome non-ATPase regulatory subunit, 26SRP)(点10)在低温下表达量增加,可能是为了清除低温胁迫中被氧化的正常蛋白质,在ABA处理下表达量增加结果则表明,该蛋白可能受到外源ABA的诱导。
ATP合成酶广泛分布于线粒体膜和叶绿体类囊体膜上。作为在高等植物能量代谢中的关键酶之一,它跨膜质子动力促进合成ATP,参与氧化磷酸化和光合磷酸化反应。本试验中ATP合成酶β-亚基(ATP synthase subunit beta, ATPase-β)(点5)的表达量上调这一结果与Kong等[22]在 OsNAS1转基因甘蓝型油菜盐胁迫条件下得到的结论一致。基于以上推测,ATP合成酶β-亚基在低温下ABA处理后参与离子转运能力增强,从而降低细胞渗透势,提高植物的抗寒性。
谷胱甘肽转移酶在植物的初级代谢和次级代谢、耐受胁迫和细胞信号转导过程中行使许多不同的功能,从而影响植物的生长发育。植物GST家族中的许多成员都与植物抗逆性有关,如对除草剂、高盐、低温和紫外辐射等的抗性[23]。Harshavardhanan等[24]在关于甘蓝GST的全基因组表征研究中,发现BoGST基因具有潜在的抗寒功能。本研究中谷胱甘肽S转移酶1(glutathione S-transferase 1, GST1)(点7)、谷胱甘肽转移酶(glutathione transferase,GST)(点11)在低温下ABA处理后表达量上调,而谷胱甘肽转移酶F5(glutathione transferase F5, GSTF5)(点15)则下调,这一结果表明GST参与植物应对低温胁迫的调控过程,一定浓度ABA能提高植物抗寒性。点15表达量下调,则可能是由于这三种不同的谷胱甘肽转移酶调控的反应不同导致的,点7、点11和点15的调控机制仍有待进一步研究。
3.3 转录翻译相关蛋白
新生多肽复合物β亚基(点3)即为基本转录因子3(basic transcription factor 3, BTF3)。有研究从小麦中克隆到一个受冻害胁迫诱导表达上调的BTF3家族基因 TaBTF3,该基因在盐、干旱、冷、ABA、MeJA和SA胁迫处理下均被诱导表达上调[25],这与本试验得到的该蛋白在低温、ABA处理下表达量上调的结果一致。
本研究中真核翻译起始因子4A(eukaryotic initiation factor 4A, eIF4A)(点6)在低温ABA处理条件下表达量增加,能够增加其与真核基因启动子区域中顺式作用元件的特异性结合,通过与其他相关蛋白的相互作用,激活转录过程,增强目的基因的表达强度,增加目的蛋白的表达量,从而影响植物抗寒能力。与Stockinger等[26]在研究拟南芥中由 CBF1编码含有转录因子的AP2结构域在低温胁迫条件下的调节作用时也得到类似的研究结果。
真核细胞翻译起始因子5A(eukaryotic translation initiation factor 5A3, eIF5A)是真菌、动植物体内普遍存在的蛋白质翻译起始因子。拟南芥的研究中发现了三个表达特征不同的eIF5Ab编码基因[27],其中,eIF5A3在细胞分裂活跃的吸胀种子中高表达,在拟南芥中过表达eIF5A3,花瓣从正常的四瓣变为五瓣,叶子变为并生叶,种子变为原来的两倍大,说明eIF5A3在发育早期促进细胞分裂。本研究中真核翻译起始因子5A3(点24)在4 ℃下特异表达,该时期恰好是冬小麦分蘖的旺盛期,与其分蘖节的细胞增殖相适应,而在-25 ℃时该蛋白消失,与小麦在-25 ℃低温下细胞分裂停滞相一致,可能是低温下eIF5A3不表达的结果。
综上所述,植物应对低温是一个复杂的调控过程。本研究发现外源ABA处理下,与对照组相比出现显著差异的蛋白可分为逆境蛋白、代谢相关蛋白和转录翻译相关蛋白。其中,代谢相关蛋白最多,包括物质代谢和能量代谢。糖代谢相关蛋白在ABA处理下表达量上调,促进植物新陈代谢,为抵御外界胁迫提供一定动力,本课题组前期研究也发现,ABA对东农冬麦1号糖代谢有促进作用[1]。ATP合成酶相关蛋白在ABA处理后表达量上调,为植物体内抵抗胁迫的各种代谢活动提供能量。有两种低温诱导的转录因子在ABA处理组表达量上调,通过在转录水平调控相关目的蛋白的表达量,从而增强植物抗寒性。逆境蛋白都在ABA处理组上调,氟啶酮处理组下调,说明ABA对冬小麦抗逆性的提高起重要作用。此外,本研究还有10种未知蛋白,这主要由于小麦的蛋白信息相对比较匮乏,但近年来蛋白质组学发展迅速,相信随着蛋白质数据库的不断完善,这些问题会很快得到解决。
[1] 刘丽杰,苍 晶,李怀伟,等.外源ABA对冬小麦越冬期呼吸代谢关键酶与糖代谢的影响[J].麦类作物学报,2013,33(1):71.
LIU L J,CANG J,LI H W,etal.Effects of exogenous abscisic acid on key enzymes of respiratory metabolism and sugar metabolism of winter wheat in the wintering period [J].JournalofTriticeaeCrops,2013,33(1):71.
[2] ZENG Y,YU J,CANG J,etal.Detection of sugar accumulation and expression levels of correlative key enzymes in winter wheat(Triticumaestivum) at low temperatures [J].BioscienceBiotechnologyBiochemistry,2011,75(4):681.
[3] TOSHIHITO Y,TAKASHI E,SHIGERU S.Restoration of seed germination at supraoptimal temperatures by fluridone,an inhibitor of abscisic acid biosynthesis [J].PlantCellPhysiology,1998,39(3):308.
[4] RASMUSSEN R D,HOLE D,HESS J R,etal.Wheat kernel dormancy and(+)-abscisic acid level following exposure to fluridone [J].JournalofPlantPhysiology,1997,150(4):444.
[5] MEZA-BASSO L,ALBERDI M,RAYNL M,etal.Changes in protein synthesis in rapeseed seedlings during a low temperature treatment [J].PlantPhysiology,1986,82(3):733.
[6] HASHIMOTO M,KOMATSU S.Proteomic analysis of rice seedlings during cold stress [J].Proteomics,2007,7(8):1293.
[7] CUI S,HUANG F,WANG J,etal.A proteomic analysis of cold stress responses in rice seedlings [J].Proteomics,2005,5(12):3162.
[8] GAO F,ZHOU Y,ZHU W,etal.Proteomic analysis of cold stress-responsive proteins inThellungiellarosette leaves [J].Planta,2009,230(5):1043.
[9] 于 晶,张 林,崔 红,等.高寒地区冬小麦东农冬麦1号越冬前的生理生化特征[J].作物学报,2008,34(11):2020.
YU J,ZHANG L,CUI H,etal.Physiological and biochemical characteristics of Dongnongdongmai 1 before wintering in high-cold area [J].ActaAgronomicaSinica,2008,34(11):2020.
[10] DAMERVAL C,VIENNE D D,ZIVY M,etal.Technical improvements in two-dimensional electrophoresis increase the level of genetic variation detected in wheat-seedling proteins [J].Electrophoresis,1986,7(1):52.
[11] SASAKI K,CHRISTOV N K,TSUDA S,etal.Identification of a novel LEA protein involved in freezing tolerance in wheat [J].PlantandCellPhysiology,2014,55(1):136.
[12] 刘 洋,邢 鑫,李德全.LEA蛋白的分类与功能研究进展[J].生物技术通报,2011(8):38.
LIU Y,XING X,LI D Q.Studies on classification and function of LEA proteins [J].BiotechnologyBulletin,2011(8):38.
[13] 唐 蜻.植物RNA结合蛋白研究进展[J].安徽农业科学,2010,38(1):38.
TANG J.Research progress on the plant RNA-binding proteins [J].JournalofAnhuiAgriculturalScience,2010,38(1):38.
[14] FUSARO A F,BOCCA S N,RAMOS R L B,etal.AtGRP2,a cold-induced nucleo-cytoplasmic RNA-binding protein,has a role in flower and seed development [J].Planta,2007,225:1348.
[15] 郭新勇,程 晨,王爱英,等.天山雪莲冷调节蛋白基因siCOR转化烟草植株的抗旱性分析[J].植物学报,2012,47(2):115.
GUO X Y,CHENG C,WANG A Y,etal.Drought-tolerance analysis of tobacco plant transformed withSasussuredinvolucratasiCORgene [J].ChineseBulletinofBotany,2012,47(2):115.
[16] KOBAVASHIA F,TAKUMIA S,NAKATAA M,etal.Comparative study of the expression profiles of theCor/Leagene family in two wheat cultivars with contrasting levels of freezing tolerance [J].PhysiologlaPlantarum,2003,120:591.
[17] AMME S,MATROS A,SCHLESIER B,etal.Proteome analysis of cold stress response inArabidopsisthalianausing DIGE-technology [J].JournalofExperimentalBotany,2006,57(7):1537.
[18] 郭 楠,赵敬会,高天姝,等.甘蓝型油菜PGK基因的克隆与表达分析[J].西北植物学报,2014,34(11):2189.
GUO N,ZHAO J H,GAO T S,etal.Cloning and expression analysis of PGK gene inBrassicanapus[J].ActaBotanicaBoreali-OccidentaliaSinica,2014,34(11):2189.
[19] SINGH R,JWA N S.Understanding the responses of rice to environmental stress using proteomics [J].JournalofProteomeResearch,2013,12:4652.
[20] KHATOON A,REHMAN S,HIRAGA S,etal.Organ-specific proteomics analysis for response mechanism in soybean seedlings under flooding stress [J].JournalofProteomics,2012,75:5706.
[21] SHRINGARPURE R,GRUNESCAPE T,MEHLHASE J,etal.Ubiquitin conjugation is not required for the degradation of oxidized proteins by proteasome [J].JournalofBiologicalChemistry,2003,278(1):311.
[22] KONG F,MAO S,DU K,etal.Comparative proteomics analysis of OsNAS1 transgenicBrassicanapusunder salt stress [J].ChineseScienceBulletin,2011,56(22):2343.
[23] 胡廷章,周大祥,罗 凯.植物谷胱甘肽转移酶的结构与功能及其基因表达[J].植物生理学通讯,2007,43(1):198.
HU T Z,ZHOU D X,LUO K.Structure and biological function of glutathione transferases and their genes in plant [J].PlantPhysiologyCommunications,2007,43(1):198.
[24] HARSHAVARDHANAN V,SENTHIL K T,ASHOKRAJ S,etal.Glutathione transferases superfamily:cold-inducible expression of distinctGSTgenes inBrassicaoleracea[J].InternationalJournalofMolecularSciences,2016,17:1211.
[25] KANG G Z,MA H Z,LIU G Q,etal.Silencing of TaBTF3 gene impairs tolerance to freezing and drought stresses in wheat [J].MolecularGeneticsandGenomics,2013,288:591.
[26] STOCKINGER E J,GILMOUR S J,THOMASHOW M F,etal.ArabidopsisthalianaCBF1 encodes an AP2 domain containing transcriptional activator that binds to the C-repeat/DRE,acis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit [J].PlantBiology,1997,94:1039.
[27] THOMPSON J E,HOPKINS M T,TAYLOR C,etal.Regulation of senescence by eukaryotic translation initiation factor 5A:implications for plant growth and development [J].TrendsinPlantScience,2004,9(4):177.
猜你喜欢
植物保护(2017年1期)2017-02-13 06:44:34
天然产物研究与开发(2016年6期)2016-06-05 10:29:26
分析测试学报(2015年6期)2016-01-13 06:19:00
中学生(2015年4期)2015-08-31 02:53:50
畜牧兽医学报(2015年3期)2015-07-05 08:22:50
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:36
新疆农垦科技(2014年5期)2014-02-28 19:20:00
新疆农垦科技(2014年2期)2014-02-28 19:19:18
食品科学(2013年24期)2013-03-11 18:30:33
食品科学(2013年13期)2013-03-11 18:24:23
