
苜蓿栽培草地微生物C、N及呼吸特性对不同播种量的响应
2018-03-26 09:16:43王笛逄焕成李达陈金强辛晓平徐丽君唐雪娟郭明英朱树声
草业学报 2018年3期
王笛,逄焕成,李达,陈金强,辛晓平,徐丽君*,唐雪娟,郭明英,朱树声
(1.中国农业科学院农业资源与农业区划研究所,呼伦贝尔国家野外站,北京100081;2.呼伦贝尔市草原工作站,内蒙古 海拉尔021200;3.牙克石市草原工作站,内蒙古 牙克石022150)
土壤呼吸是土壤释放二氧化碳(CO2)、与大气交换的过程,是生态系统物质循环和能量流动的重要组成部分[1-2]。其中包括微生物呼吸、根系呼吸、土壤动物呼吸3个生物学过程。土壤呼吸是全球碳循环中最大的碳通量之一,约占生态系统呼吸的60%~90%[3]。其中影响土壤呼吸的因素除了温度、湿度、pH等非生物因素外,也受到植被、微生物等生物因素的影响,并且随着人类活动的增强,人为因素的作用也日益严重[4]。植物不同播种量的种植会影响植物地上、地下生物量的生长,从而影响土壤呼吸,成为影响土壤呼吸的主要因素之一。另一方面土壤微生物是促进有机质、养分循环和转化的驱动力,是反映土壤质量的重要组成部分[5-6]。土壤微生物分解有机质获得其生长所需能量的同时,产生CO2并排放到大气中[6-8],形成土壤呼吸中的微生物呼吸,所以微生物呼吸也是判断土壤肥力等指标的重要因素[9-11]。土壤微生物呼吸及其熵值(qMB,qCO2)作为土壤质量的敏感性指标,可以用于预测土壤有机碳的长期变化趋势[12]。研究表明,土壤微生物量碳(microbial biomass C)、氮(microbial biomass N)占土壤总碳、总氮的比例虽然较小,却是微生物量活性中最活跃的部分[11-14],我国土壤微生物量碳占土壤总碳量的2%~4%,被认为是影响土壤健康和质量最关键的因素[15],土壤微生物量氮占土壤总氮量的1%~5%[14],氮元素是植株生长发育所必需的生长因素,在草地生态系统中有着至关重要的作用[16],土壤微生物量碳、氮也可以综合反映土壤肥力和生物活性[17],综上,草地土壤呼吸、土壤微生物以及土壤微生物呼吸是反映植被生长发育、土壤健康的重要指标,而栽培草地不同播种量的种植方式,会导致植被群落地上、地下生物量的密度及质量,从而影响各项指标,所以寻求适宜的播种量以达到植物产量、土壤质量协同提高的试验势在必行。
呼伦贝尔草原是我国温带草甸草原分布最具代表性的地区,是具有典型原生态特征的保存最完整的草原之一[18-19]。近年来,天然草原不同程度的退化,严重制约着草地畜牧业的健康发展,栽培草地作用日益凸显[20],苜蓿(Medicagosativa)是蛋白含量高、适口性良好、营养价值较高的豆科牧草,被称为“牧草之王”[20],由于商品经济的发展,苜蓿的市场需求量加大,使苜蓿产业的规模快速发展,其中在呼伦贝尔地区种植苜蓿的面积逐年增加,合理的种植结构是影响栽培草地可持续利用的首要问题[21]。因此,本研究在多年种植苜蓿的试验基础上,选择5个不同播种量的方式,研究苜蓿栽培草地微生物C、N及呼吸特性对不同播种量的响应,探讨该地区最适宜的苜蓿播种量,以期为该地区苜蓿栽培和利用提供理论依据。
1 材料与方法
1.1 试验区概况及试验设计
1.1.1试验区概况 试验地选择呼伦贝尔草原生态系统国家野外试验站(内蒙古海拉尔市谢尔塔拉镇)栽培草地试验田,地处N 49°06′-49°32′,E 119°32′-120°35′,研究区域内水热条件较好,属于温带大陆季风性气候,海拉尔河与伊敏河交汇于此,无霜期为110 d左右,年降水量250~400 mm,自东南向西北递减,年平均气温-2 ℃,自东南向东北递增,土壤以黑钙土为主,区域内耕地开垦面积较大。代表性植被类型为温带草甸草原,代表群系有苜蓿、羊草(Leymuschinensis)、贝加尔针茅(Stipabaicalensis)、草地早熟禾(Poapratensis)、山野豌豆(Viciaamoena)等。
1.1.2供试材料 选择种植年限为2年的龙牧801(MedicagosativaLongmu 801)为供试材料。由于苜蓿初花期营养物质含量最高,苜蓿利用以收干草为主,分别在2015年7月26日和9月25日进行第1次和第2次刈割。
1.1.3试验设计 在2014年6月5日选择地势平坦、土壤肥沃的土地播种,种植方式为条播,行距为15 cm,不同梯度的播种量分别为6,9,12,15和18 kg·hm-2,每个处理3次重复,随机区组设计,每个处理的试验面积为3 m×5 m,共15组试验地。
1.2 土样采集
于2015年7月26日、9月25日在15个处理组分别取0~10 cm、10~20 cm土壤,采集方法为“ S ”型多点采集,每小区选取5个点混合作为一个样品,共30份土壤样品,标记好后装入塑料自封袋迅速带回实验室,过2 mm筛,一部分暂存于4 ℃冰箱,5日内用于土壤微生物量碳、氮测定,其余自然风干后过筛保存备用。
1.3 样品测定方法
1.3.1分枝数 选择植株生长状况基本相同的样地,设置每小区0.5 m×0.5 m样方,于2015年7月26日和2015年9月25日统计单位面积内苜蓿分枝数,共计45组样方。
1.3.2生物量 地上生物量:15组样地中每小区设置3个0.5 m×0.5 m的样方,在7月26日和9月25日进行土壤样品收集之前,将样方中的植株留茬10 cm,其余均匀剪齐并收集至信封袋中,称其鲜重,之后将其放入65 ℃烘箱烘24 h至恒量,称重;地下生物量:15组样地中每小区设置3个0.5 m×0.5 m×0.2 m的样方在土层深度0~20 cm采集地下生物量,将植株与根系上粘附的土壤洗净,将其放入80 ℃烘箱烘24 h至恒量,称重。
根冠比:地上生物量与地下生物量干重的比值。
1.3.3微生物量碳、氮测定 使用氯仿熏蒸浸提法[22-23]进行土壤微生物量碳、氮的研究,其中微生物量碳测定采用重铬酸钾硫酸外加热法[24],微生物量氮的测定采用半微量凯氏定氮法[25],其含量计算用熏蒸和未熏蒸土样之差除以系数Kc=0.38和Kn=0.54,公式如下:
土壤微生物量碳(mg/kg)=(Ec-Ec0)/0.38
式中:Ec为熏蒸土壤浸提液中有机碳量;Ec0为不熏蒸土壤浸提液中有机碳量;0.38为校正系数。
土壤微生物量氮(mg/kg)=(En-En0)/0.54
式中:En为熏蒸土壤浸提液中氮量;En0为不熏蒸土壤浸提液中氮量;0.54为校正系数。
1.3.4微生物呼吸与呼吸熵 本研究采用直接呼吸法,使用CO2容量盐酸滴定测定[22]。称取5组预培养的土壤20 g于烧杯中,每组重复3次,共15只烧杯,均匀加入蒸馏水调节土壤含水量为最大含水量60%,将盛土烧杯移入2.5 L广口瓶中,在广口瓶内同时放入20 mL 0.1 mol·L-1氢氧化钠标准溶液,用于吸收土壤微生物所释放的CO2,随后将广口瓶置入25 ℃恒温人工培养箱继续培养,试验开始后的1,2,4,7,11和15 d对取出的氢氧化钠溶液进行盐酸滴定,同时注意每次滴定后更新氢氧化钠标准溶液,以备下次滴定。呼吸熵是微生物呼吸与微生物量碳间比率,即每单位微生物生物量碳的具体呼吸速率[7]。
1.4 数据处理
经Excel整理数据后,采用SPSS 20.0软件进行单因素方差(One-way ANOVA)分析,不同处理之间多重比较采用LSD(Least-significant difference)方法,然后经过T检验(P<0.05),相关分析采用双变量(Spearman相关系数)分析,然后经过双侧检验(P<0.05)。
2 结果与分析
2.1 生产性能比较
随着播种量的增加,7月苜蓿的分枝数、地上和地下生物量基本呈现增加趋势,其中分枝数与地上生物量在处理4时达到最大值,分枝数处理4显著高于处理1(P<0.05),其数值大小比较如下:处理1<处理2<处理3<处理5<处理4,地上生物量与地下生物量各处理之间差异不显著(P>0.05);9月经过一次刈割后苜蓿的分枝数在处理5时达到最大值,显著高于处理1(P<0.05),其数值大小比较如下:处理1<处理2<处理4<处理3<处理5,地上生物量与地下生物量在处理3时达到最大值,处理5其次,地上生物量处理3数值显著高于处理2(P<0.05),地下生物量各处理数值差异不显著(P>0.05)。
9月苜蓿的分枝数相较7月分枝数平均增加63.00%,增加最多的是处理1达到85.73%;地上生物量平均减少51.38%,减少最多的是处理4达到63.48%;地下生物量平均增加18.89%,增加最多的是处理3达到29.75%(表1)。
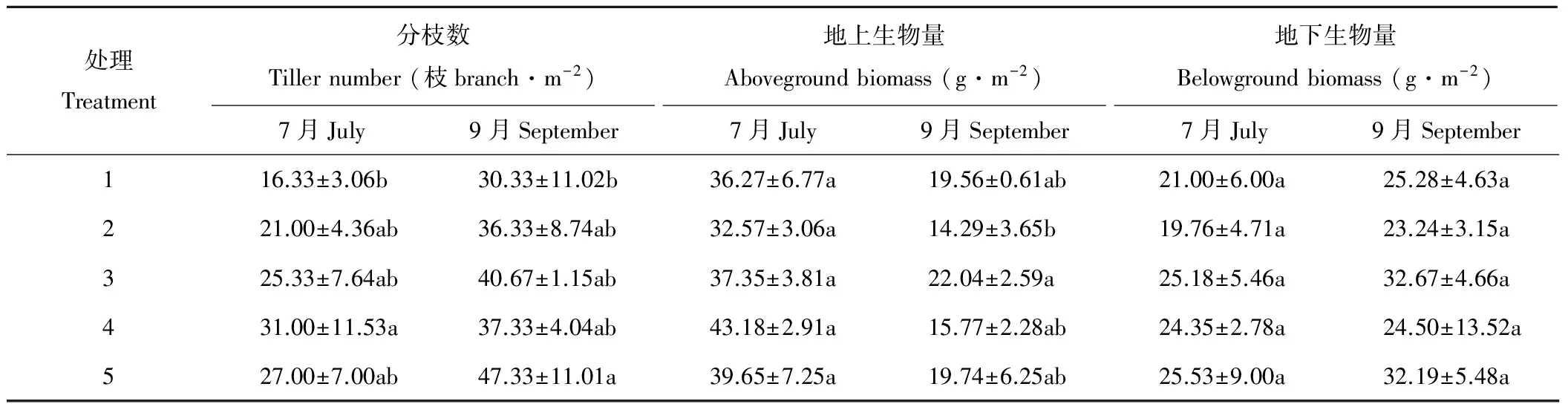
表1 不同播种量条件下不同月份苜蓿分枝数、地上生物量、地下生物量的比较Table 1 Comparison of tiller number, aboveground biomass and belowground biomass of alfalfa in different months under different sowing quantity
注:1代表播种量6 kg·hm-2,2代表9 kg·hm-2,3代表12 kg·hm-2,4代表15 kg·hm-2,5代表18 kg·hm-2,下同;同列数字后不同字母表示处理间差异显著(P<0.05)。
Note: In the Table number one mean sowing amount 6 kg·ha-1, two mean sowing amount 9 kg·ha-1, three mean sowing amount 12 kg·ha-1, four mean sowing amount 15 kg·ha-1, five mean sowing amount 18 kg·ha-1, the same below; Values followed by different letters in the same column mean significant differences at 5% level.
2.2 植物根冠比比较
随着播种量的增加,第一次刈割(7月)苜蓿的地上生物量总体呈增加趋势,第二次刈割(9月)苜蓿的地上生物量与7月刈割的地上生物量差异明显,但与7月刈割地下生物量呈正比例关系,表明7月刈割苜蓿的根系发育决定了之后苜蓿的再生性(图1和图2);由于刈割致使地上生物量降低,导致9月苜蓿的根冠比远高于7月刈割的苜蓿根冠比,结合表1可以发现7月分枝数最多的处理4其根冠比反而最低,其原因可能是因为生长密度的增加有利于水分的固持,使得地上生物量生长迅速,地下生物量生长较为缓慢,到9月刈割时根系越发达的处理其地上部分产量越高。
2.3 土壤微生物生物量碳、氮对不同播种量的响应
7月土层0~10 cm微生物生物量碳处理1最高,但各处理之间差异不显著(P>0.05),说明土层0~10 cm微生物生物量碳随着播种量增加变化平稳,微生物量碳最高的处理1与微生物量碳最低的处理3之间相差34.39%;土层10~20 cm微生物生物量碳处理5最高,显著高于其他处理(P<0.05),说明土层10~20 cm微生物生物量碳随着播种量增加变化剧烈,微生物量碳最高的处理5与微生物量碳最低的处理4之间相差66.47%(图3)。
9月土层0~10 cm微生物生物量碳处理5最高,显著高于处理1、2和3(P<0.05),微生物量碳最高的处理5与微生物量碳最低的处理1之间相差60.68%;土层10~20 cm微生物生物量碳也是处理5最高,但处理之间差异不显著(P>0.05),说明土层10~20 cm微生物生物量碳随着播种量增加变化平稳,微生物量碳最高的处理5与微生物量碳最低的处理3之间相差29.52%(图4)。
7月土层0~10 cm微生物生物量氮处理5最高,并且显著高于其余各组(P<0.05),微生物量氮最高的处理5与微生物量氮最低的处理4之间相差52.14%;土层10~20 cm微生物生物量氮处理3最高,显著高于处理5(P<0.05),微生物量氮最高的处理3与微生物量氮最低的处理5之间相差43.10%(图5)。
9月土层0~10 cm微生物生物量氮处理2最高,显著高于其余各处理(P<0.05),微生物量氮最高的处理2与微生物量氮最低的处理4之间相差63.46%;土层10~20 cm微生物生物量氮处理3最高,显著高于处理5(P<0.05),微生物量氮最高的处理3与微生物量氮最低的处理5之间相差48.30%(图6)。
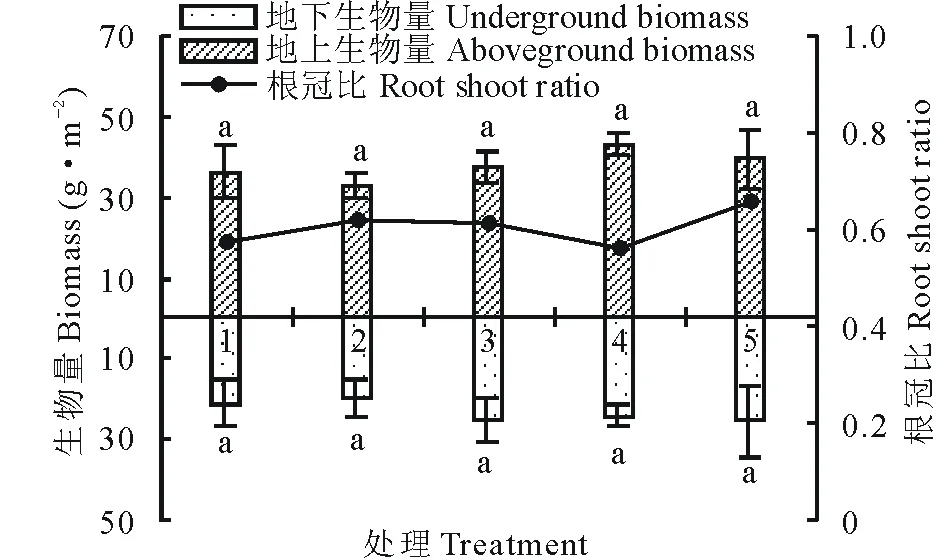
图1 7月不同播种量下根冠比的比较Fig.1 Comparison of root shoot ratio at different sowing quantity in July
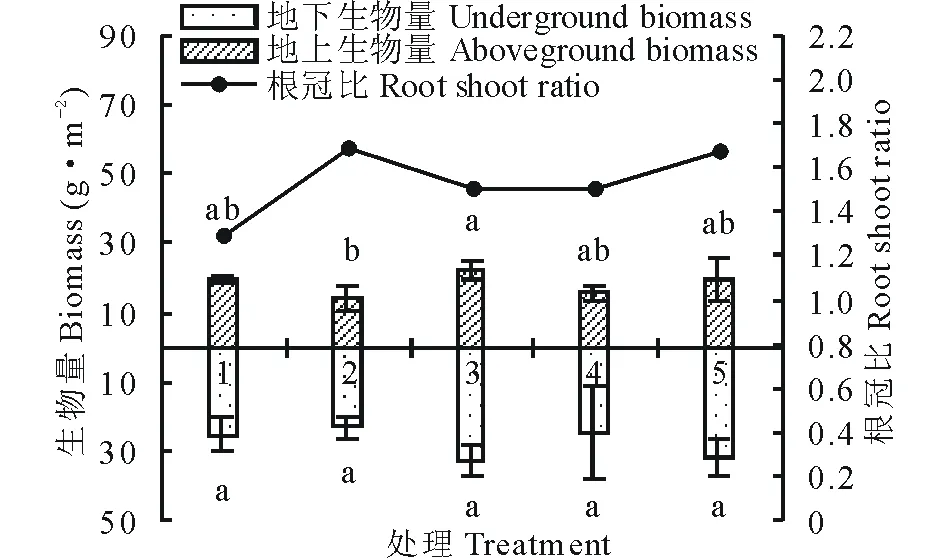
图2 9月不同播种量下根冠比的比较Fig.2 Comparison of root shoot ratio at different sowing quantity in September
不同字母表示处理间差异显著(P<0.05)。下同。The different letters indicate significant difference between the treatments (P<0.05).The same below.
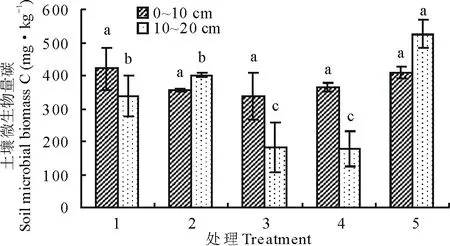
图3 7月不同播种量下土壤微生物量碳变化Fig.3 Changes of soil microbial biomass carbon under different sowing quantity in July
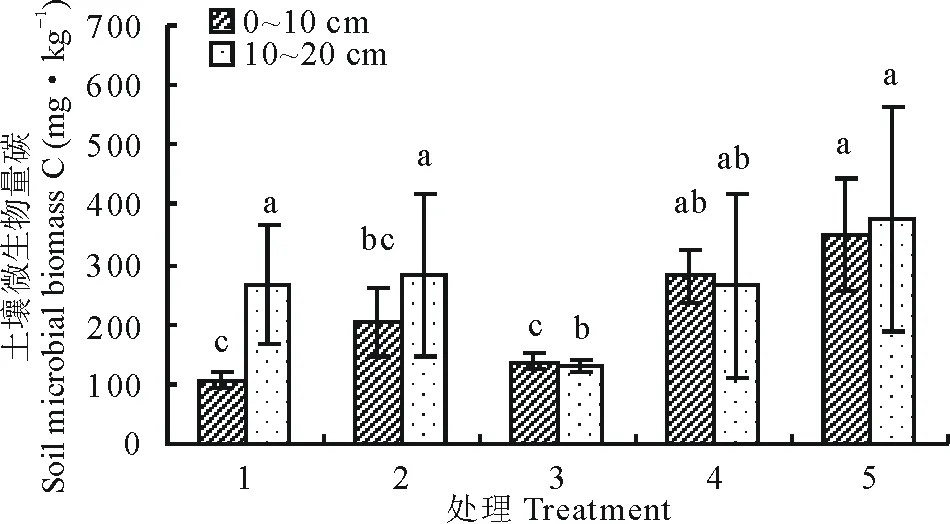
图4 9月不同播种量下土壤微生物量碳变化Fig.4 Changes of soil microbial biomass carbon under different sowing quantity in September
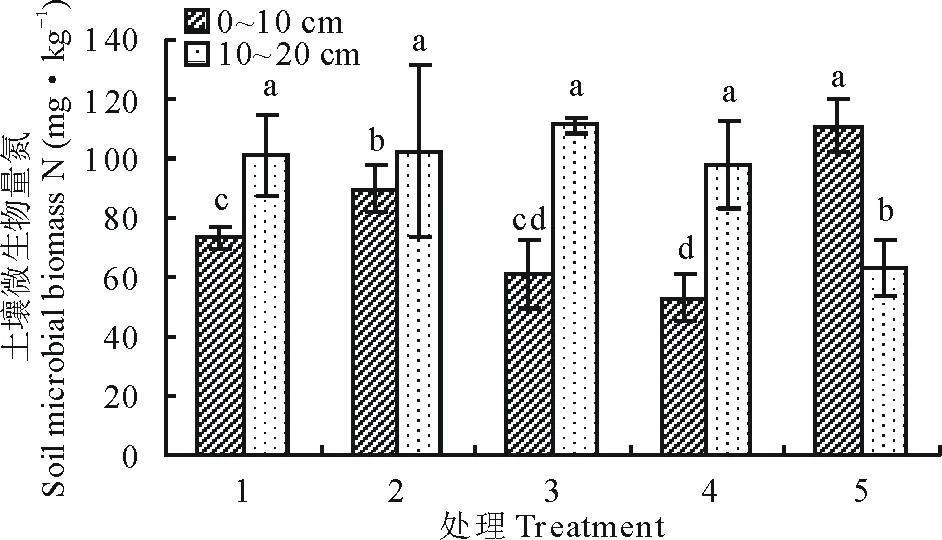
图5 7月不同播种量下土壤微生物量氮变化Fig.5 Changes of soil microbial biomass nitrogen under different sowing quantity in July
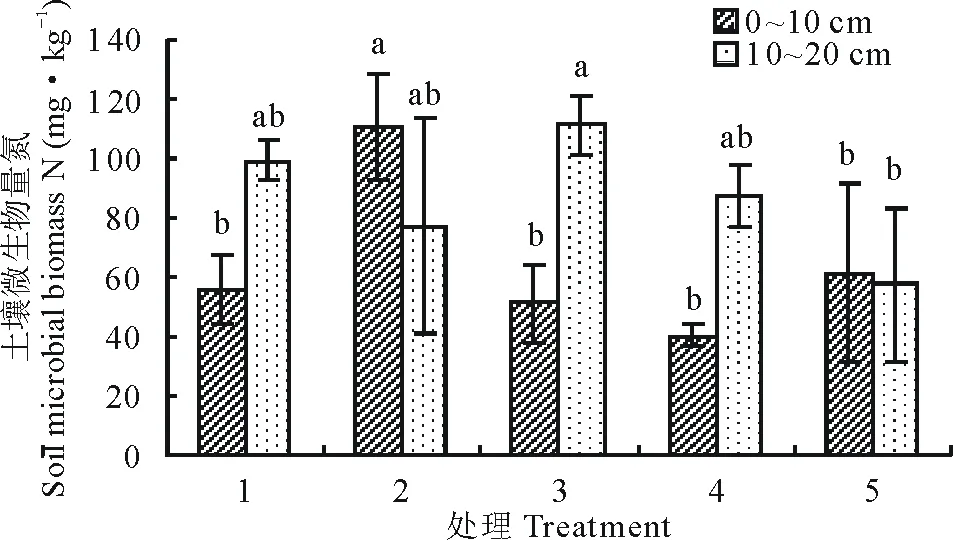
图6 9月不同播种量下土壤微生物量氮变化Fig.6 Changes of soil microbial biomass nitrogen under different sowing quantity in September
2.4 土壤微生物呼吸及呼吸熵对不同播种量的响应
7月土壤深浅两层微生物呼吸及呼吸熵随着播种量的增加(图7和图8)均呈现出较为一致的先降低后升高趋势,土层0~10 cm处理1微生物呼吸、呼吸熵数值最高,趋势变化为由处理1降低至处理3,后上升至处理5,但各处理之间差异不显著(P>0.05)。
9月土壤深浅两层微生物呼吸及呼吸熵随着播种量的增加(图9和图10)表现出相反的趋势,土层0~10 cm微生物呼吸处理4和5显著高于其余各处理(P<0.05);呼吸熵处理4显著高于其余各处理(P<0.05),处理5其次;土层10~20 cm微生物呼吸处理2最高,显著高于处理3和4(P<0.05),处理1其次;呼吸熵处理1最高,处理2其次,但各处理之间差异不显著(P>0.05)。
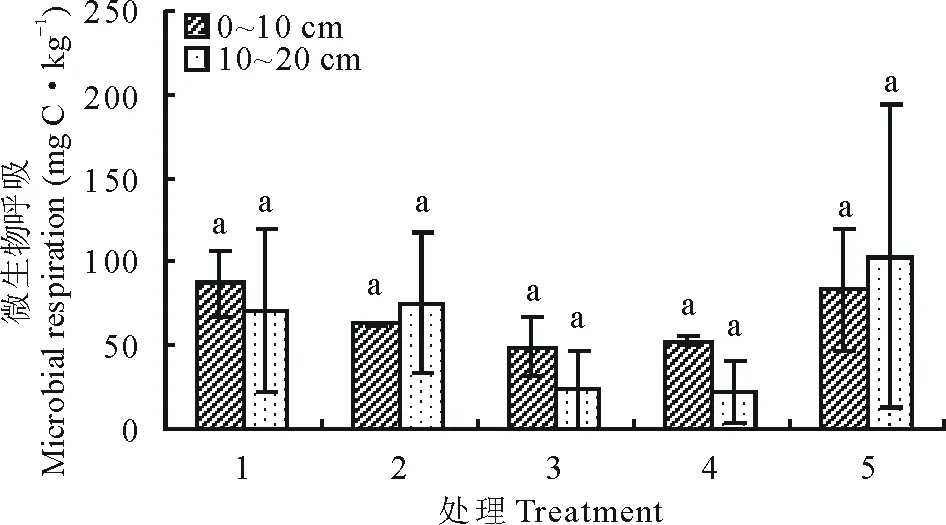
图7 7月不同播种量下土壤微生物呼吸变化Fig.7 Changes of soil microbial respiration under different sowing quantity in July
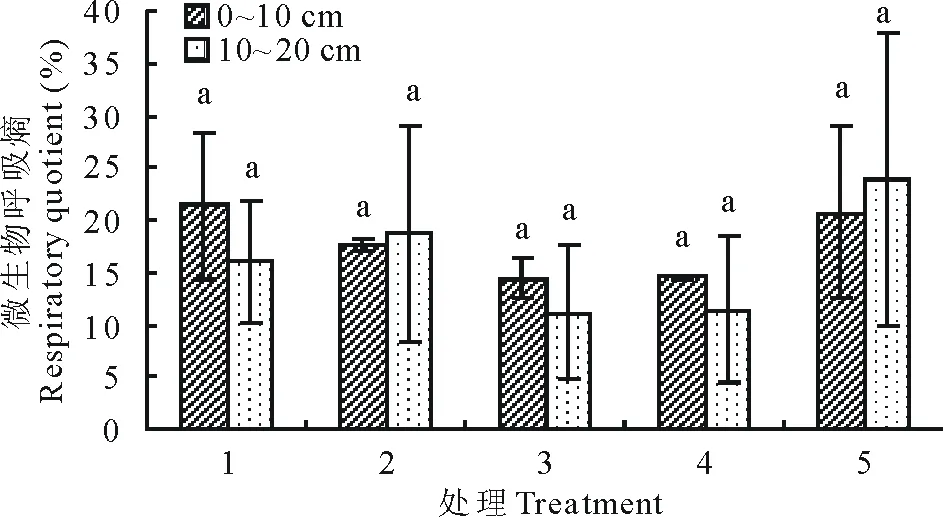
图8 7月不同播种量下土壤微生物呼吸熵变化Fig.8 Changes of soil microbial respiratory quotient under different sowing quantity in July
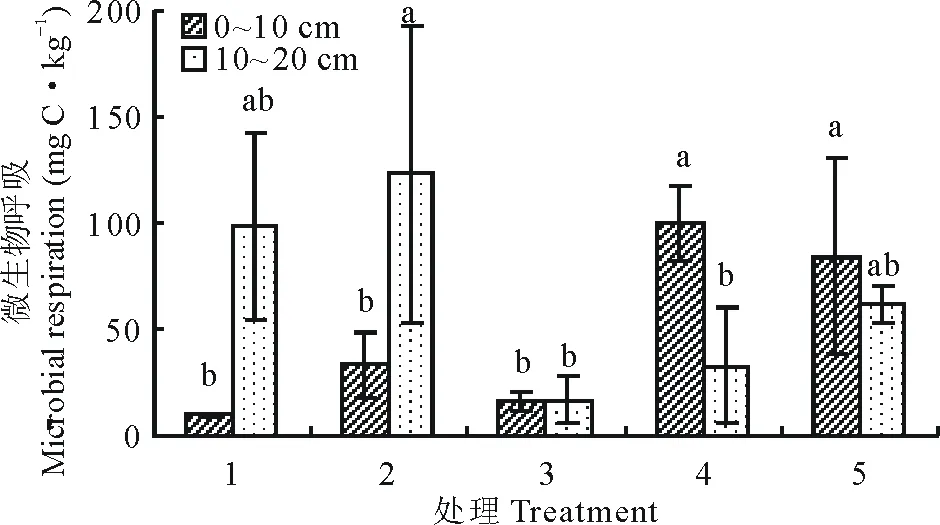
图9 9月不同播种量下土壤微生物呼吸变化Fig.9 Changes of soil microbial respiration under different sowing quantity in September
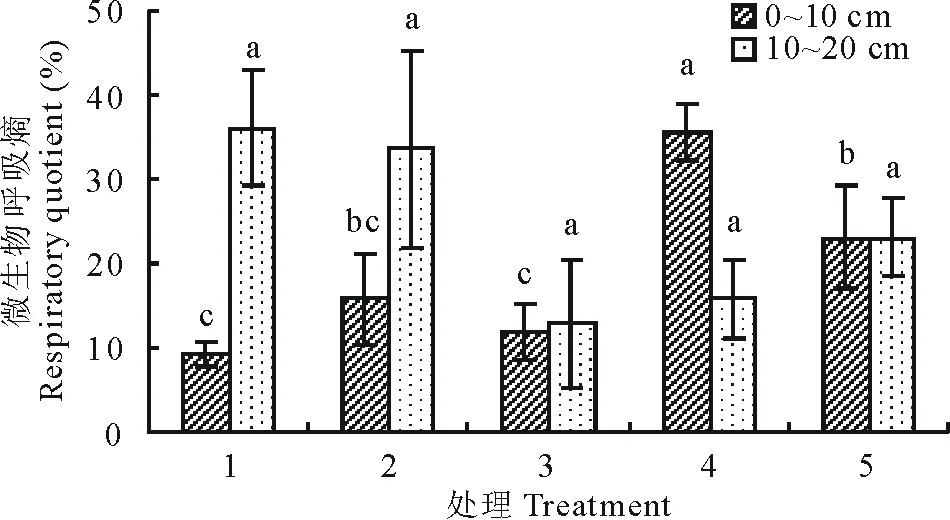
图10 9月不同播种量下土壤微生物呼吸熵变化Fig.10 Changes of soil microbial respiratory quotient under different sowing quantity in September
2.5 呼吸熵与微生物特性相关分析
通过对不同时期不同土层深度下呼吸熵与微生物特性相关关系进行分析(表2),结果表明,呼吸熵与微生物呼吸、微生物量碳呈正相关,其中除7月0~10 cm土层微生物量碳外均达到显著水平(P<0.05);呼吸熵与微生物量氮除7月0~10 cm土层外其余呈负相关,但未达到显著水平(P>0.05)。
3 讨论
苜蓿栽培草地不同播种量的种植方式,会直接反映苜蓿地上、地下生物量的不同;从而导致凋落物质量、数量发生变化,凋落物回归土壤导致土壤元素发生变化,元素变化则导致微生物发生变化[26],所以不同播种量对微生物呼吸以及微生物C、N有重要影响。土壤微生物调控土壤中有机化合物的转化和利用以及养分的吸持和释放,且土壤中的一系列过程均以碳、氮循环为中心[27-28]。
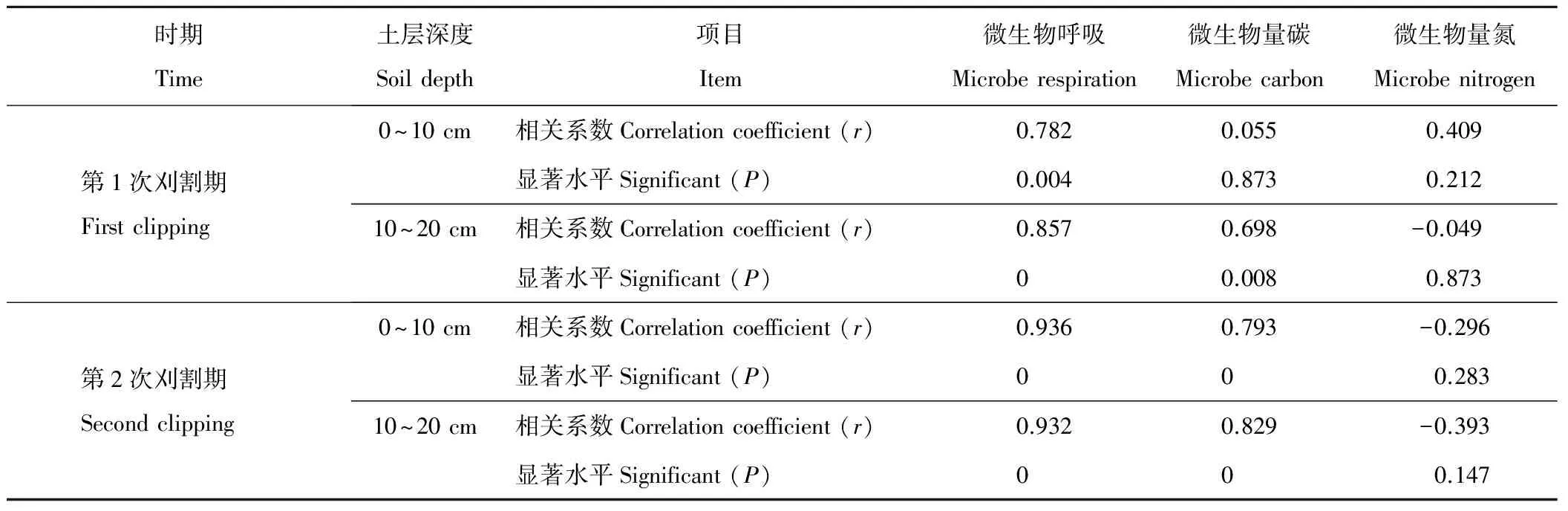
表2 呼吸熵与微生物特性相关分析Table 2 Correlation analysis between respiratory quantity and microbial characteristics
土壤微生物碳不仅能反映微生物对环境作用的能力,还能通过微生物量碳的情况揭示土壤有机质的状态和积累情况[23-24],本研究结果表明,微生物生物量碳7月10~20 cm土层与9月0~10 cm和10~20 cm土层均在播种量为18 kg·hm-2时达到最大值,说明土壤微生物量碳随播种量变化,在本实验中18 kg·hm-2为最佳播种量,但伴随其变化趋势应当在之后实验中设置更高梯度,从而排除样本容量的影响来验证18 kg·hm-2是否为促进土壤微生物量碳的最佳播种量;通过两个月份的对比可以看出数值上微生物量碳7月整体高于9月,说明随着时间变化土壤微生物量碳呈降低趋势,表层土壤可能由于微生物活动剧烈并且容易受到外界环境因素影响导致其变化明显[27]。
微生物量氮是指活的微生物体内所含有的氮,是主要的可矿化氮源,对土壤氮素供应和循环具有重要的意义[27-30]。数值上微生物量氮7月整体高于9月,则说明随着时间变化土壤微生物量氮也呈降低趋势,这种相似的规律说明7月是植株生长旺期,其植株的生长以及根系的原因都有利于土壤碳、氮的积累,而9月微生物量氮数值较低则说明刈割后苜蓿再生对土壤养分的吸收导致土壤微生物量碳氮的整体含量逐渐降低。本研究中微生物氮与碳的结果不同,9月土层0~10 cm、10~20 cm均因播种量的增加大致呈先升高后降低的趋势,微生物氮与碳在7月0~10 cm变化趋势基本相同,同样说明在土层0~10 cm微生物量变化较为平稳,而其他3种情况变化较为剧烈,其原因可能是由于植被建植第二年,表层土壤植株根系、凋落物趋于稳定,但在深层土壤10~20 cm植株根系仍在生长发育,特别是建植第二年,苜蓿总生物量的增加导致了植物根系的变化,这与黄懿梅等[27]研究结果基本相同。
微生物呼吸强度是判断土壤微生物总活性的指标[31],本研究结果表明,7与9月微生物呼吸基本呈先降低后上升的趋势,而9月土层0~10 cm呼吸强度则随着播种量的增加呈直线上升趋势,其原因可能是由于7月第一次刈割后植株要经历第二次生长发育,播种密度越高则根系活动作用越突出,所以导致呼吸变化呈上升趋势。同时结果表明,呼吸熵与微生物呼吸变化趋势一致,但呼吸熵代表的是土壤受胁迫程度,有研究表明呼吸熵值越低,则说明土壤受胁迫程度越小,其土壤环境越成熟[27]。结合本试验,播种量为12 kg·hm-2时呼吸熵值最低,说明该植株播种量土壤环境最成熟稳定;在播种量为18 kg·hm-2时呼吸强度与呼吸熵的数值与播种量为12 kg·hm-2时差异不显著,说明18 kg·hm-2时土壤环境同样趋于稳定;另一方面随着播种量的增加呼吸熵与微生物量碳呈正相关,除7月0~10 cm土层外均达到显著水平,分析其原因可能是随播种量的增加凋落物太多或质量发生变化导致土壤中C发生变化;呼吸熵与微生物量氮除7月0~10 cm土层外其余呈负相关,但未达到显著水平,分析其原因可能是因为土壤氮长期累积的结果,短期内是难以检测到比较明显的变化[32]。
生物量是人工栽培草地的重要指标,实验中发现7月生物量高于9月生物量,同时微生物碳、氮与之相同,7月高于9月,其原因可能是生物量的增加,导致凋落物的增加,凋落物回归土壤则影响微生物量碳、氮数值变化,但其具体反映的机理过程还需进一步研究[26]。通过根冠比的高低可以看出7月播种量为18 kg·hm-2的处理最大,其微生物碳高于其余各组,0~10 cm土层的微生物氮显著高于其余各组;9月播种量为9和18 kg·hm-2的处理的根冠比较高,对应的微生物量碳18 kg·hm-2的处理最高,微生物量氮在0~10 cm土层9 kg·hm-2的处理最高,18 kg·hm-2的处理其次,通过微生物碳、氮与生物量的变化可以发现生物量对微生物量碳影响较为明显,而对微生物量氮只能影响到表层土壤,因此,不同播种量苜蓿种植方式与微生物氮的响应关系应有待进一步研究。
4 结论
1)结果中显示随播种量的增加微生物呼吸熵与微生物呼吸、微生物量碳呈正相关;
2)土壤微生物碳、氮及土壤微生物呼吸对苜蓿不同播种量有一定的响应作用;
3)不同播种量结合植株生物量、土壤微生物呼吸、土壤微生物量碳、氮等指标结果显示在本实验中播种量为18 kg·hm-2时效果最好。
References:
[1] Cui X Y, Chen Z Z, Chen S Q. Progress in research on soil respiration of grasslands. Acta Ecologica Sinica, 2001, 21(2): 315-325.
崔骁勇, 陈佐忠, 陈四清. 草地土壤呼吸研究进展. 生态学报, 2001, 21(2): 315-325.
[2] Liu J, Zhao Y, Zhang Q M,etal. Effects of land use on soil microbial biomass and community structure in the loess hill region of west Henan. Acta Prataculturae Sinica, 2016, 25(8): 36-47.
刘晶, 赵燕, 张巧明, 等. 不同利用方式对豫西黄土丘陵区土壤微生物生物量及群落结构特征的影响. 草业学报, 2016, 25(8): 36-47.
[3] Schimel D S, Louse J I, Libbard K A,etal. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature, 2001, 414(6860): 169-172.
[4] Liu S H, Fang J Y. Effect factors of soil respiration and the temperatures effects on soil respiration in the global scale. Acta Ecologica Sinica, 1997, 17(5): 469-476.
刘绍辉, 方精云. 土壤呼吸的影响因素及全球尺度下温度的影响. 生态学报, 1997, 17(5): 469-476.
[5] Ma X X, Wang L L, Li Q H,etal. Effects of long-term fertilization on soil microbial biomass carbon and nitrogen and enzyme activities during maize growing season. Acta Ecologica Sinica, 2012, 32(17): 5502-5511.
马晓霞, 王莲莲, 黎青慧, 等. 长期施肥对玉米生育期土壤微生物量碳氮及酶活性的影响. 生态学报, 2012, 32(17): 5502-5511.
[6] Wang W J, Dalal R C, Moody P W,etal. Relationships of soil respiration to microbial biomass, substrate availability and clay content. Soil Biology & Biochemistry, 2003, 35(2): 273-284.
[7] Zhu R F, Tang F L, Liu J L,etal. Response of soil microbial biomass carbon and nitrogen on a short-term fertilizing N inLeymuschinensismeadow. Acta Agrestia Sinica, 2016, 24(3): 553-558.
朱瑞芬, 唐凤兰, 刘杰淋, 等. 羊草草甸草原土壤微生物生物量碳氮对短期施氮的响应. 草地学报, 2016, 24(3): 553-558.
[8] Zhang P J, Li L Q, Pan G X,etal. Influence of long-term fertilizer management on topsoil microbial biomass and genetic diversity of a paddy soil from the Tai Lake region. Acta Ecologica Sinica, 2004, 24(12): 2818-2824.
张平究, 李恋卿, 潘根兴, 等. 长期不同施肥下太湖地区黄泥土表土微生物碳氮量及基因多样性变化. 生态学报, 2004, 24(12): 2818-2824.
[9] Wang Y, Li Z J, Han B,etal. Effects of conservation tillage on soil microbial characters and soil enzyme activities. Acta Ecologica Sinica, 2007, 27(8): 3384-3390.
王芸, 李增嘉, 韩宾, 等. 保护性耕作对土壤微生物量及活性的影响. 生态学报, 2007, 27(8): 3384-3390.
[10] Hu Y L, Wang S L, Huang Y,etal. Effects of litter chemistry on soil biological property and enzymatic activity. Acta Ecologica Sinica, 2005, 25(10): 2662-2668.
胡亚林, 汪思龙, 黄宇, 等. 凋落物化学组成对土壤微生物学性状及土壤酶活性的影响. 生态学报, 2005, 25(10): 2662-2668.
[11] Jagadish C T, Subhash C M, Shyam K. Influence of straw size on activity and biomass of soil microorganisms during decomposition. European Journal of Soil Biology, 2001, 37: 157-160.
[12] Liu S S, Wang F, Zhang X H,etal. Effect of grazing disturbance on soil microbial biomass and respiratory quotient in mountain forest of arid valley ecotone in upper reaches of Minjiang river. Bulletin of Soil and Water Conservation, 2014, 34(2): 63-68.
刘珊珊, 王芬, 张兴华, 等. 放牧干扰对岷江上游山地森林-干旱河谷交错带土壤微生物量及呼吸熵的影响. 水土保持通报, 2014, 34(2): 63-68.
[13] Mers I W, Schinn Er F. An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride. Biology and Fertility of Soils, 1991, 11: 216-220.
[14] Smith J L, Paul E A. The significance of soil microbial biomass estimations//Bollag J M, Stotzky G. Soil biochemistry. New York: Marcel Dekker, Inc., 1990.
[15] Qu W H, Li Z G, Li J. Effects of fertilizing organic materials on contents of carbon and nitrogen, enzyme activity of desertified soils and alfalfa biomass. Pratacultural Science, 2017, 34(3): 456-464.
屈皖华, 李志刚, 李健. 施用有机物料对沙化土壤碳氮含量、酶活性及紫花苜蓿生物量的影响. 草业科学, 2017, 34(3): 456-464.
[16] Gao C, Zhang Y X, Chen J S,etal. Analysis of soil nitrogen dynamics ofMedicagosativaandLeymuschinensispasture in the Songnen Plain. Pratacultural Science, 2015, 32(4): 501-507.
高超, 张月学, 陈积山, 等. 松嫩平原苜蓿和羊草栽培草地土壤氮素动态分析. 草业科学, 2015, 32(4): 501-507.
[17] Zhang H B, Yang G X, Huang Q,etal. Analysis of dynamic spatial-temporal changes of landscape patterns in Hulunbur meadow steppes-taking Hailaer and surrounding areas as an example. Acta Prataculturae Sinica, 2009, 18(1): 134-143.
张宏斌, 杨桂霞, 黄青, 等. 呼伦贝尔草甸草原景观格局时空演变分析——以海拉尔及周边地区为例. 草业学报, 2009, 18(1): 134-143.
[18] Liu Y, Han S J, Hu Y L,etal. Effects of soil temperature and humidity on soil respiration rate in pinus olgensis forest. Chinese Journal of Applied Ecology, 2005, 16(9): 1581-1585.
刘颖, 韩士杰, 胡艳玲, 等. 土壤温度和湿度对长白松林土壤呼吸速率的影响. 应用生态学报, 2005, 16(9): 1581-1585.
[19] Shanghai Society for Plant Physiology. Laboratory manual of plant physiology. Shanghai: Shanghai Science and Technology Press, 2009: 3.
上海植物生理学会. 植物生理学实验手册. 上海: 上海科学技术出版社, 2009: 3.
[20] Sun Q Z, Yu Z, Xu C C. Urgency of further developing alfalfa industry in China. Pratacultural Science, 2012, 29(2): 314-319.
孙启忠, 玉柱, 徐春城. 我国苜蓿产业亟待振兴. 草业科学, 2012, 29(2): 314-319.
[21] Jiang Y M, Yao T, Li J H,etal. Effects of different management measures on soil microbial biomass in an alpine meadow. Acta Prataculturae Sinica, 2016, 25(12): 35-43.
蒋永梅, 姚拓, 李建宏, 等. 不同管理措施对高寒草甸土壤微生物量的影响研究. 草业学报, 2016, 25(12): 35-43.
[22] Brookes P C, Landman A, Pruden G,etal. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology & Biochemistry, 1985, 17(6): 837-842.
[23] Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology & Biochemistry, 1987, 19(6): 703-707.
[24] Lin Q M, Wu Y G, Liu H L. Modification of fumigation extraction method for measuring soil microbial biomass carbon. Chinese Journal of Ecology, 1999, (2): 63-66.
林启美, 吴玉光, 刘焕龙. 熏蒸法测定土壤微生物量碳的改进. 生态学杂志, 1999, (2): 63-66.
[25] Liu Y, Zhou L R, Miao S J. Effects of long-term fertilization on mollisoil enzyme activities and microbial respiration. Soil and Fertilizer in China, 2010, (1): 7-10.
刘妍, 周连仁, 苗淑杰. 长期施肥对黑土酶活性和微生物呼吸的影响. 中国土壤与肥料, 2010, (1): 7-10.
[26] Xie T T. Effects of litter changes after simulating forest canopy damage on soil nitrogen mineralization rate. Beijing: Chinese Academy of Forestry, 2013.
谢亭亭. 模拟林冠受损后凋落物变化对土壤氮矿化速率的影响. 北京: 中国林业科学研究院, 2013.
[27] Huang Y M, An S S, Xue H. Responses of soil microbial biomass C and N and respiratory quotient (qCO2) to revegetation on the Loess Hilly-Gully region. Acta Ecologica Sinica, 2009, 29(6): 2811-2818.
黄懿梅, 安韶山, 薛虹. 黄土丘陵区草地土壤微生物C、N及呼吸熵对植被恢复的响应. 生态学报, 2009, 29(6): 2811-2818.
[28] Zhao J, Geng Z C, Shang J,etal. Effects of biochar and biochar-based ammonium nitrate fertilizers on soil microbial biomass carbon and nitrogen and enzyme activities. Acta Ecologica Sinica, 2016, 36(8): 2355-2362.
赵军, 耿增超, 尚杰, 等. 生物炭及炭基硝酸铵对土壤微生物量碳、氮及酶活性的影响. 生态学报, 2016, 36(8): 2355-2362.
[29] Bastida F, Barberá G G, García C,etal. Influence of orientation, vegetation and season on soil microbial and biochemical characteristics under semiarid conditions. Applied Soil Ecology, 2008, 38(1): 62-70.
[30] Breuer L, Huisman J A, Keller T,etal. Impact of a conversion from cropland to grassland on C and N storage and related soil properties: Analysis of a 60-year chronosequence. Geoderma, 2006, 133(1/2): 6-18.
[31] Insam H. Are the soil microbial biomass and basal respiration governed by the climatic regime. Soil Biology & Biochemistry, 1990, 22(4): 525-532.
[32] Grove S J. Extent and composition of dead wood in Australian lowland tropical rainforest with different management histories. Forest Ecology and Management, 2001, 154(22): 35-33.
猜你喜欢
现代畜牧科技(2021年9期)2021-10-13 06:38:52
现代畜牧科技(2021年4期)2021-07-21 06:13:04
河南农业·综合版(2021年2期)2021-03-24 11:27:38
河南农业(2021年2期)2021-03-14 10:06:34
湖北农业科学(2020年22期)2020-12-18 07:32:08
今日农业(2020年19期)2020-12-14 14:16:52
河南农业(2018年12期)2018-12-05 08:06:22
中国三峡(2017年4期)2017-06-06 10:44:22
为了孩子(3~7岁)(2016年6期)2016-05-14 03:09:17
中国科技信息(2015年2期)2015-11-16 08:18:32
