
帽儿山不同年龄森林土壤呼吸速率的影响因子
2018-03-22 11:33:17王家骏王传宽
生态学报 2018年4期
王家骏, 王传宽, 韩 轶
东北林业大学生态研究中心, 哈尔滨 150040
陆地生态系统的最大碳库是土壤有机碳库(约1500Pg C),是大气碳库的两倍[1],因此在全球碳循环中发挥着重要作用[2]。土壤呼吸(RS)是陆地生态系统碳循环的重要环节[3],也是土壤碳排放的主要途径[4],每年全球RS总量大约为68—98Pg C[5- 7],相当于化石燃料所释放碳的10倍[8]。因此,即使RS发生微小的变化,也可能对大气中的CO2浓度、全球碳平衡产生显著的影响。
RS受气候变化、植被类型、土壤性质等众多因子的影响[9- 11],其中林龄是一个重要的影响因素[12]。随着林龄的增加,森林的物种组成、群落结构、生物量积累以及碳分配等均会发生变化[13- 15],从而能显著地影响森林与大气的碳交换,但有关林龄对RS影响的研究结果尚存在较大的不确定性。例如,Ewel等[16]的热带森林研究报道,RS随林龄增大而增加;而Wang等[17]对加拿大北方森林火烧演替序列的研究发现,RS随林龄增大呈现先增后减的趋势,Klopatek等[18]对美国中南部花旗松采伐年龄序列的研究和也得出类同的结果;Tang等[12]对美国五大湖地区山杨年龄序列的研究则报道,随林龄增大RS先增大后减小。这些研究结果的不确定性可能与干扰方式或研究地区不同有关,但还需要更多的验证研究。
东北林区是我国重点林区,在我国的生态环境建设和区域生物地球化学循环中起着至关重要的作用。该地区历经半个多世纪采伐干扰,林地面积显著减小,原始林基本殆尽,森林年龄结构趋于单一化和低龄化。近年来随着退耕还林、天然林保护等林业生态工程的实施,出现了大面积的皆伐后自然恢复的林分[19- 20]。然而,不同年龄林分的RS特征及其影响因子尚不清楚。本研究选择测定帽儿山地区皆伐后天然更新恢复的4个年龄(1a、10a、25a和56a)的林分的RS,目的为了探究(1)不同年龄林分RS季节变化趋势以及土壤温度和含水率对其影响;(2)不同年龄林分全年、生长季、非生长季RS通量随林龄的变化趋势及其影响因素。
1 材料和方法
1.1 研究地概况和样地设置
研究地位于黑龙江帽儿山森林生态系统国家野外科学观测研究站(45°24′N,127°40′E)。该地区具有大陆性温带季风气候,冬季寒冷干燥,夏季短促湿热。平均年降水量629mm,大约50%的降水集中于6—8月,平均年蒸发量864mm。年平均气温3.1℃,年平均总日照时数1857h,无霜期约为120—140d,平均海拔400m。土壤为地带性暗棕色森林土壤。植被属于长白植被区系,现有植被是原地带性植被-阔叶红松林屡遭人为采伐后经过次生演替而形成的次生林,代表着东北东部山区典型的森林类型。
选取皆伐后天然更新恢复过程中的4个年龄(1a、10a、25a和56a)的林分,每个林分设置3个重复样地,每个样地大小为20m×30m。1年样地为皆伐迹地,其余3个年龄林分均为次生林(表1)。
1.2 土壤呼吸及相关环境因子的测定
2013年5月初在每个样地内布置8个内径为10.2cm、高为5cm的PVC土壤环。将PVC环的一端削尖,压入土中,减少布置土壤环对土壤的镇压作用,并保持土壤环在整个测定期间位置不变。RS测定采用自制的气室连接到LI- 840 CO2/ H2O分析仪(LI-COR Co.,Lincoln,USA)上进行。2014年4月—2015年3月,生长季内每月测定2次,非生长季每月测定1次。与此同时,在土壤环的附近用T型热电偶温度探针测定5cm深度的土壤温度(T5),用TDR100土壤水分速测仪(Spectrum Technologies,USA)测定0—5cm深度的土壤含水率(体积含水率,W5)。2014年11月—2015年3月由于土壤冻结,W5未能测定。
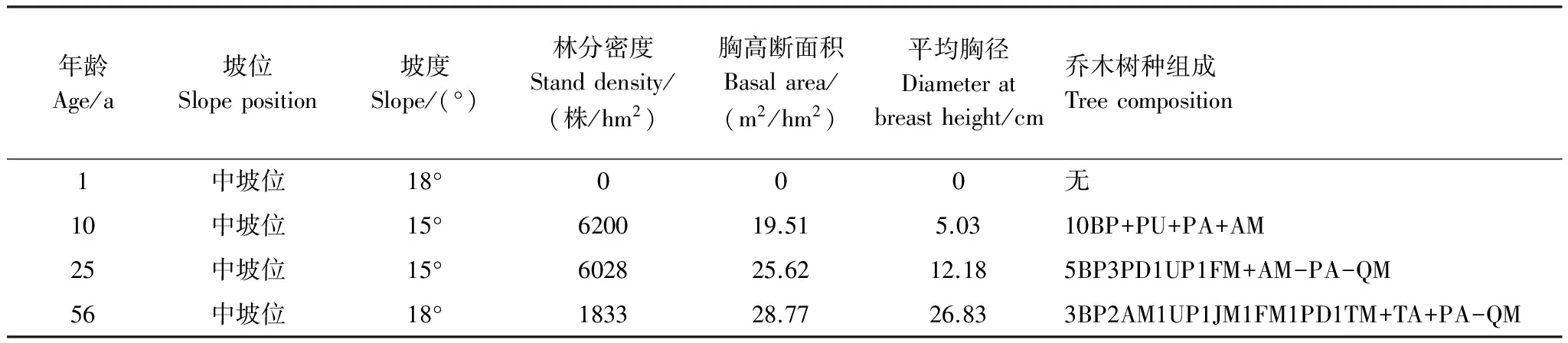
表1 4种年龄林分的立地状况和乔木层组成
PD:山杨,Populusdavidiana;AM:五角槭,Acermono;BP:白桦,Betulaplatyphylla;QM:蒙古栎,Quercusmongolica;JM:胡桃楸,Juglansmandshurica;FM:水曲柳,Fraxinusmandshurica;TA:紫椴,Tiliaamurensis;PA:黄菠萝,Phellodendronamurense;UP:春榆,Ulmuspropinqua; TM:糠锻,Tiliamandshurica;PU:大青杨,Populusussuriensis. 树种组成中的数字表示该树种胸高断面积所占的成数,“+”、“-”分别表示组成比例<5%和<2%
细根生物量的测定采用根钻法。2015年9月底在各年龄林分的每个土壤环附近1m处,用内径为10cm的根钻取土,深至50cm,取土每隔10cm一层,共5层。将取回的土样分成两份,一份风干后测定土壤有机碳(SOC)浓度;另一份先用作测定根系生物量。先将土壤放在网筛上洗净,然后根据外形、颜色、弹性等特征将根分为死根和活根;再进一步将活根按其直径分为两级:L1级(0—0.5mm)和L2级(0.5—2mm);最后将分级后的样品在65℃下烘干至恒重并称重。
1.3 数据分析
土壤呼吸(RS,μmolCO2m-2s-1)与土壤温度(T5,℃)之间关系采用指数函数模型[21]模拟:
RS=R0eβT
(1)
式中,R0为0℃时的土壤呼吸(μmolCO2m-2s-1),β为温度系数。
土壤呼吸与土壤含水率(W5,%)之间关系采用二次函数模型[22]模拟:
RS=aW52+bW5+c
(2)
根据以往研究[10,23-26],土壤呼吸与土壤温度和土壤含水率的关系采用下列模型[17]模拟:
ln(RS)=d+fT5+gW5+hT5W5
(3)
式中:a、b、c、d、f、g、h为模型参数。
采取多元线性回归法针对每个年龄林分分别拟合模型(1)和(2),获得表2;然后运用表2中的回归模型计算得到每天RS值。对比该模拟值与实测值(图1)发现:虽然全年4个年龄林分的RS与T5均呈极显著的指数函数关系(P<0.01;表2),但是通过指数函数预测的全年RS通量在6—8月与实测值离差较大;而6—8月采用模型(2)不但模型本身拟合效果较好(P<0.05),而且模拟值与实测值相吻合(图2)。因此,本研究中各林龄年龄的RS采用了分段模拟估算,即6—8月期间采用模型(2)的二次函数关系估算,其余月份则采用模型(1)的指数函数关系估算。
在比较不同年龄样地间RS的差异时,由于受到测定时间的限制,不可能在同一时间内对每个土壤环进行同步测定,而且各年龄样地的土壤温度和土壤含水率本身就存在差异,因此不能用每次实测RS直接进行比较。为了比较各年龄样地的RS,先建立每个样地实测的T5和W5与附近土壤温度和含水率自动数据采集器在同一时刻所记录的T5和W5之间的回归模型,然后通过数据采集器的连续测定值来获取每个样地的连续的T5和W5,然后代入各年龄样地RS的回归模型,以天为时间步长累加得到各年龄林分的生长季、非生长季和全年RS通量。

表2 土壤呼吸(RS)与土壤温度(T5)和土壤含水率(W5)的拟合模型
表中二次模型为RS=aW52+ bW5+ c,而指数模型为:RS=R0eβT;前者为6─8月的关系,而后者为全年其余月份的关系

图1 利用0─5cm土层的土壤温度(T5)或土壤含水率(W5)模拟不同年龄林分土壤呼吸值和实测值比较Fig.1 Comparisons between measured and modeled soil respiration rates with soil temperature (T5) or soil water content (W5) at the 5cm depth for different aged stands图中A1和A2为T5控制阶段,B为W5控制阶段
用重复测量方差分析(repeated measures ANOVA)来分析林龄和测定时间对RS的影响,并用邓肯检验(Duncan test)比较不同年龄林分T5、W5以及RS的差异。采用多元线性回归分析各年龄林分全年RS通量与T5、W5、SOC密度以及细根生物量的关系。所有数据分析由SPSS 19.0(SPSS Inc.,Chicago,Illinois,USA)完成,图表绘制由Sigmaplot 10.0(Systat Software Inc.,San Jose,CA,USA)完成。
2 结果与分析
2.1 不同年龄林分土壤呼吸的季节动态及其影响因子
所有林分的T5的季节变化趋势基本一致,最高值均出现在6—8月的生长季,最低值出现在1—2月的冬季(图2)。然而,不同年龄林分W5的季节变化趋势除了在6—8月期间基本一致以外,其他月份分异较大。不同年龄样地之间T5在生长季期间表现出的变异性大于非生长季;相反,W5在生长季期间不同年龄样地之间的差异小于非生长季(图2)。
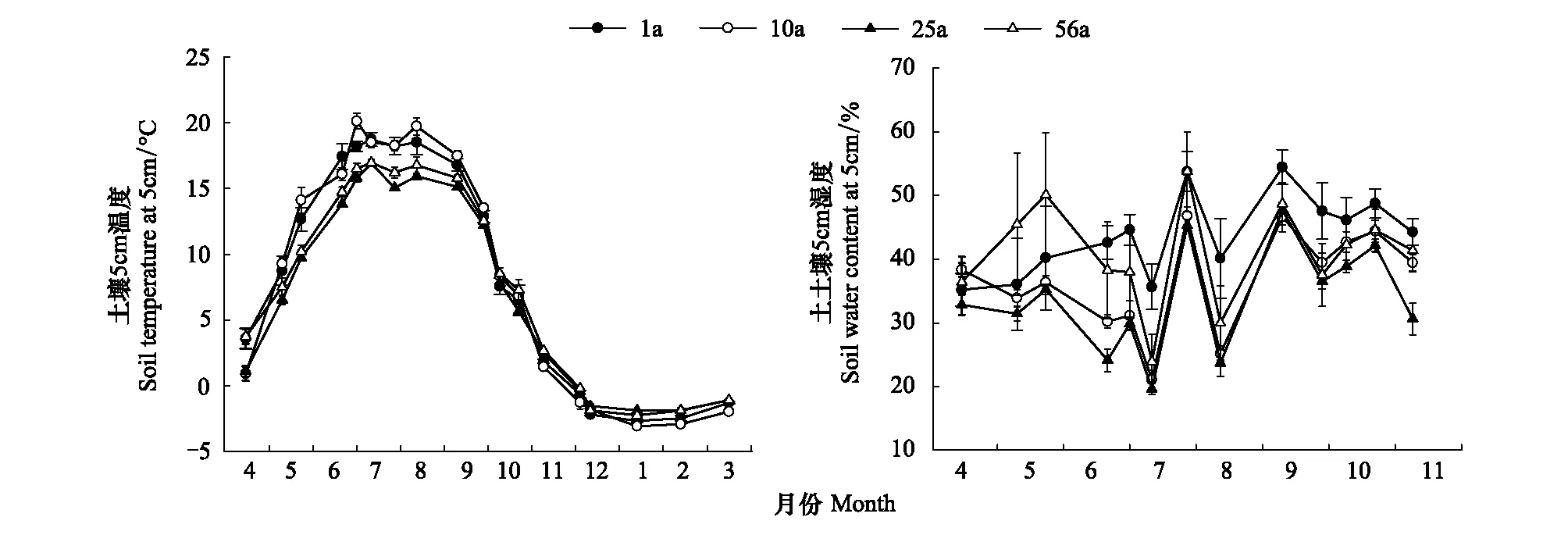
图2 不同年龄林分5cm深土壤温度和土壤含水率的季节变化Fig.2 Seasonal variations in soil temperature and soil water content at the 5cm depth in different aged stands
4个年龄林分的RS的季节动态基本一致,均与T5的变化趋势基本一致,表现为生长季高、非生长季低的单峰型变化格局(图1)。不过,在6月—8月期间,RS实测值明显低于基于T5的模型估测值,因此可以将RS的季节动态分为两个阶段:土壤含水率控制阶段(2014年6月—2014年8月,即B阶段)和土壤温度控制阶段(2014年4月—2014年5月和2014年9月—2015年3月,即A1和A2阶段)(图1)。在土壤温度控制阶段内,T5对1年、10年、25年和56年林分RS变异性的解释率分别为85%、93%、88%和90%;而在土壤含水率控制阶段,W5对1年、10年、25年和56年林分RS变异性的解释率分别为56%、63%、79%和57%(表2)。
2.2 不同年龄林分土壤呼吸的差异及其影响因子
不同年龄林分的全年T5平均值和4月—11月期间W5平均值均无显著差异(表3),但生长季和非生长季T5平均值的林分间差异显著。重复测量方差分析结果显示,年龄(P=0.013)和测定时间(P<0.001)对RS均有显著影响,但两者的交互作用不显著(P=0.076)。不同年龄林分的全年、生长季和非生长季RS平均值均存在显著差异(P<0.05),分别波动在1.75—2.09g C m-2d-1,3.21—3.72g C m-2d-1、0.27—0.45g C m-2d-1之间(表3),均以10年样地最低、25年或56年样地最高。
表3不同年龄样地土壤5cm温度(T5)和土壤5cm含水率(W5)以及土壤呼吸(RS)的多重比较(平均值±标准误差)
Table3Multiplecomparisonbetweeneachofagesstandsinsoiltemperature(T5)andwatercontentatthe5cmdepth(W5)andsoilrespiration(mean±SE)
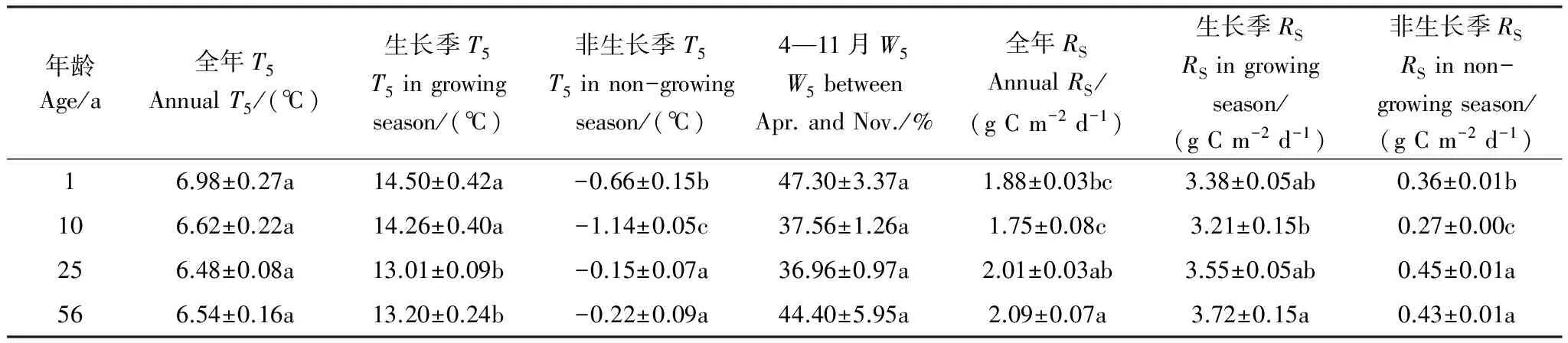
年龄Age/a全年T5AnnualT5/(℃)生长季T5T5ingrowingseason/(℃)非生长季T5T5innon-growingseason/(℃)4—11月W5W5betweenApr.andNov./%全年RSAnnualRS/(gCm-2d-1)生长季RSRSingrowingseason/(gCm-2d-1)非生长季RSRSinnon-growingseason/(gCm-2d-1)16.98±0.27a14.50±0.42a-0.66±0.15b47.30±3.37a1.88±0.03bc3.38±0.05ab0.36±0.01b106.62±0.22a14.26±0.40a-1.14±0.05c37.56±1.26a1.75±0.08c3.21±0.15b0.27±0.00c256.48±0.08a13.01±0.09b-0.15±0.07a36.96±0.97a2.01±0.03ab3.55±0.05ab0.45±0.01a566.54±0.16a13.20±0.24b-0.22±0.09a44.40±5.95a2.09±0.07a3.72±0.15a0.43±0.01a
表中同列不同小写字母表示不同年龄差异显著(P<0.05)
4个年龄林分全年RS通量波动在639.7g C m-2a-1与762.3g C m-2a-1之间;生长季RS通量波动在590.6g C m-2a-1至684.8g C m-2a-1之间;非生长季RS通量波动在49.1g C m-2a-1至80.6g C m-2a-1;全年和生长季RS通量均表现为10年样地<1年样地<25年样地<56年样地的顺序,非生长季RS通量表现为10年样地<1年样地<56年样地<25年样地的顺序(图3)。全年、生长季和非生长季RS通量随林龄的变异系数分别为7.6%、6.3%和21.1%。
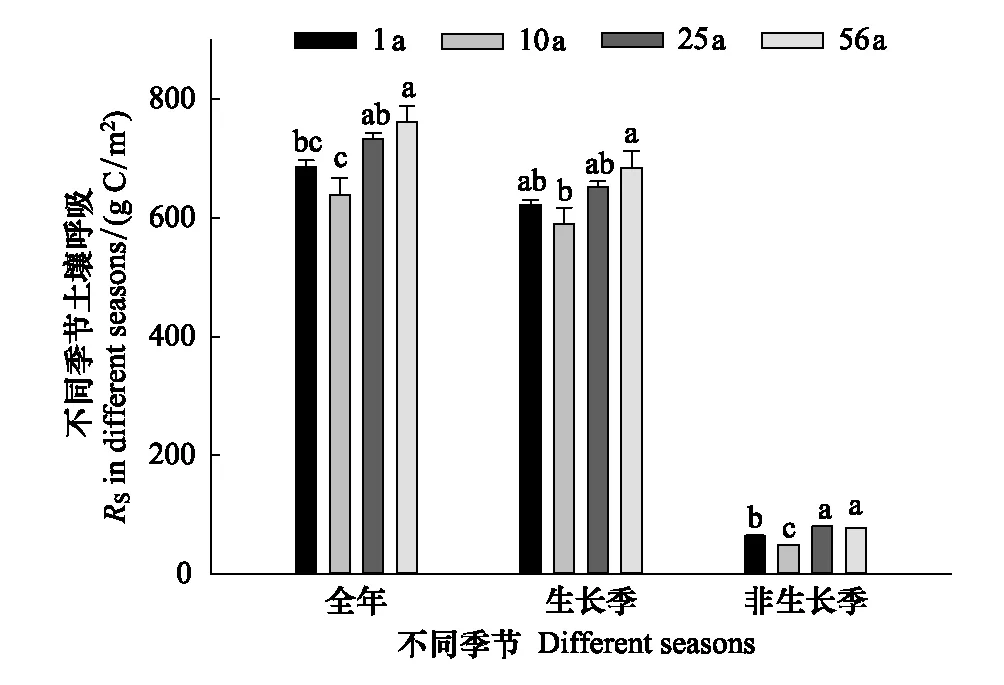
图3 不同年龄林分全年、生长季和非生长季RS通量比较(平均值±标准误差,n=3)Fig.3 Comparisons of RS in the whole year, growing season and non-growing season among different aged stands (mean±SE, n=3)不同字母表示不同年龄林分差异显著,P<0.05
不同年龄林分RS与T5、W5、不同土深(0—10、0—20、0—40cm)的细根生物量以及SOC密度之间的关系分析表明:生长季RS通量与0—10cm土层中直径<0.5mm的细根生物量呈显著正相关关系(R2=0.390,P=0.03),与0—20cm土层的SOC密度也呈显著正相关关系(R2=0.434,P=0.02)。然而,非生长季RS通量与相应的T5呈显著的正相关关系(R2=0.959,P<0.01)(图4)。
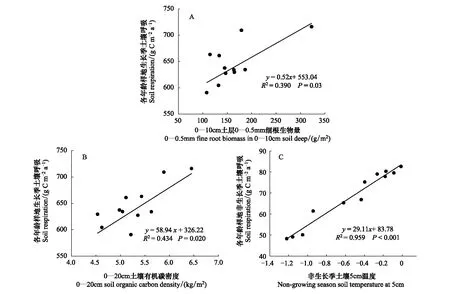
图4 不同年龄林分生长季或非生长季RS通量与影响因子之间的关系Fig.4 Relationships between the RS in the growing or non-growing season and affecting factors for all aged stands(A)为生长季RS通量与0—10cm土层直径< 0.5mm的细根生物量的关系,(B)为生长季RS通量与0—20cm土层SOC密度的关系,(C)为非生长季RS通量与5cm深土壤温度(C)之间的关系
3 结论与讨论
3.1 不同年龄林分土壤呼吸的季节变化及其影响因子
本研究中4个皆伐后自然恢复的不同年龄林分的RS呈现一致的季节动态(图1),与在南亚热带森林[27]和北方森林[12]的研究结果类似,这可能是RS受到土壤温度和土壤含水率的强烈控制的缘故[28-29]。在同一研究区内,气候条件(气温和降水量等)基本一致,土壤温度和土壤含水率的变化模式相似,因而不同年龄林分的RS表现出相似的季节变化趋势[30]。除6月—8月外,4种年龄林分RS的季节变化与T5变化趋势一致,两者之间呈极显著的指数函数关系(表2),表明土壤温度是影响该时段土壤呼吸时间动态的主要因子,这与其他温带森林的研究结果一致[12,31-32]。
然而,6月—8月期间,4个年龄林分的RS与T5之间的关系均不显著(P> 0.05;图1)。这并不能说明这段时段内RS与T5就没有相关关系,可能是由于这段时间内T5波动较小,使得RS与T5统计关系不显著。相反,这一时段的W5变化剧烈(图2),从而上升为RS的主导因子:RS与W5有显著的二次函数关系(R2波动在0.56—0.79之间;表2)。这一结果与Li等对黄土高原森林等11种群落RS的研究结果相似[33]。有研究证实:当土壤含水率不是限制条件时,RS主要受土壤温度的控制;而当RS受到土壤含水率的胁迫时,RS的变化在温度的驱动下受土壤含水率的控制[9,34-35]。当土壤含水率较低时,土壤中可溶性底物的扩散受到抑制;在一定土壤含水率范围内,RS随着土壤含水率的增加而增大。当土壤含水率超过一定的数值后,土壤中气体的扩散速率降低和氧气缺乏,就会抑制土壤中CO2的排放[36-37]。因此,土壤含水率过低或过高都会抑制凋落物分解、根呼吸以及土壤微生物的活动,从而限制RS的大小[38-40]。
3.2 不同年龄林分土壤呼吸的差异及其影响因子
本研究中4个年龄林分全年RS通量波动在639.7—762.3g C m-2a-1之间,处于以往温带森林研究结果范围(122—1754g C m-2a-1[28,41])之内。4个年龄林分全年和生长季RS通量均差异显著,表现出随皆伐后森林恢复的年龄增加,RS通量呈先减后增的趋势(图3)。皆伐1年后生长季RS通量比56年生林分低(表3),这是由于皆伐后植被细根生物量迅速减少所导致的(皆伐迹地和56年生林分的直径<0.5mm的细根生物量分别为1.22g/m2和1.75g/m2)。在皆伐后森林恢复初期,一方面土壤SOC因分解加快而减少[42],另一方面因林分郁闭而使抵达地面的太阳辐射减少,土壤温度降低(表2),而且植被根系尚未得以恢复,从而使10a林分的RS通量仍然显著小于56a(图3)。随着森林的进一步恢复,植被根系得以恢复,同时林地凋落物产量不断增加[27,43],SOC的增加为微生物呼吸提供了大量底物,从而使得RS随之增大。在这一时段,SOC密度增加起着重要的作用。
本研究中4个年龄林分非生长季RS通量波动在49.1—80.6g C m-2之间(图3),占全年RS通量的比例分别为9.4%、7.7%、11.0%和10.2%;这与Wang等[10]报道的温带森林中非生长季RS占全年RS通量的比例(8%—25%)相符。生长季和非生长季中不同年龄RS的大小趋势基本一致(表2,图3),但不同年龄林分之间非生长季RS通量的变异系数(21.1%)是生长季的(6.3%)3.35倍。这说明虽然非生长季RS通量占全年RS通量的比例较小,但仍然加大了不同年龄林分全年RS通量的差异,因此在评价森林恢复过程中RS通量的时间动态时不能简单地仅考虑生长季的观测值。
冬季土壤呼吸主要来自微生物呼吸[44]。一般认为,非生长季RS的土壤温度临界值处在-7℃至-5℃之间,低于这个温度微生物呼吸会因土壤中自由水的缺乏而受到抑制[45-46]。研究发现T5高于-3.5℃就能维持活跃的土壤呼吸。不同年龄林分非生长季RS通量的差异主要是由非生长季T5驱动引起的。25年和56年生林分非生长季T5显著高于其他样地(表3),可能是因为它们的凋落物层较厚,能部分阻挡土壤至大气的长波辐射[47]。皆伐迹地可能由于采伐残留物对土壤有一定的保温作用而使其T5高于10年生林分(表3),因而其非生长季RS通量较高。可见,不同年龄林分非生长季RS通量的差异主要是由于枯落物层不同引起土壤温度差异所导致的。
本研究中RS随着森林恢复呈先减后增的趋势,与北方森林RS随森林恢复先增后降[12,48-49]和逐渐降低[50]的研究结果不一致。主要原因可能是本研究所采用的年龄序列中的最小年龄样地为皆伐迹地,最大年龄为56年生林分(本地区的老龄林超过200年),因此没能包括整个次生演替系列。而在森林演替过程中,生态系统的物种组成、土壤理化性质[51-52]、微生物群落多样性[53]、土壤胞外酶[54]等都会发生变化,进而影响RS对森林演替响应的模式。因此,土壤微生物呼吸在森林恢复过程中的变化机理还需深入研究。
致谢:本文数据的测定和分析得到王兴昌、周正虎、张著及黑龙江帽儿山森林生态系统国家野外科学观测研究站工作人员的帮助,特此致谢。
[1] Luo Y Q, Zhou X H. Soil Respiration and the Environment. London: Academic Press, 2006.
[2] 顾峰雪, 黄玫, 张远东, 李洁, 闫慧敏, 郭瑞, 钟秀丽. 氮输入对中国东北地区土壤碳蓄积的影响. 生态学报, 2016, 36(17): 5379- 5390.
[3] 王新源, 李玉霖, 赵学勇, 毛伟, 崔夺, 曲浩, 连杰, 罗永清. 干旱半干旱区不同环境因素对土壤呼吸影响研究进展. 生态学报, 2012, 32(15): 4890- 4901.
[4] Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7- 20.
[5] Raich J W, Potter C S, Bhagawati D. Interannual variability in global soil respiration, 1980- 94. Global Change Biology, 2002, 8(8): 800- 812.
[6] Bond-Lamberty B, Thomson A. Temperature-associated increases in the global soil respiration record. Nature, 2010, 464(7288): 579- 582.
[7] Vicca S, Bahn M, Estiarte M, Van loon E E, Vargas R, Alberti G, Ambus P, Arain M A, Beier C, Bentley L P, Borken W, Buchmann N, Collins S L, de Dato G, Dukes J S, Escolar C, Fay P, Guidolotti G, Hanson P J, Kahmen A, Kröel-Dulay G, Ladreiter-Knauss T, Larsen K S, Lellei-Kovacs E, Lebrija-Trejos E, Maestre F T, Marhan S, Marshall M, Meir P, Miao Y, Muhr J, Niklaus P A, Ogaya R, Peuelas J, Poll C, Rustad L E, Savage K, Schindlbacher A, Schmidt I K, Smith A R, Sotta E D, Suseela V, Tietema A, van Gestel N, van Straaten O, Wan S, Weber U, Janssens I A. Can current moisture responses predict soil CO2efflux under altered precipitation regimes? A synthesis of manipulation experiments. Biogeosciences, 2014, 11(11): 2991- 3013.
[8] Zhu B, Cheng W X. Rhizosphere priming effect increases the temperature sensitivity of soil organic matter decomposition. Global Change Biology, 2011, 17(6): 2172- 2183.
[9] Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles, 1995, 9(1): 23- 36.
[10] Wang C K, Yang J Y, Zhang Q Z. Soil respiration in six temperate forests in China. Global Change Biology, 2006, 12(11): 2103- 2114.
[11] 刘绍辉, 方精云. 土壤呼吸的影响因素及全球尺度下温度的影响. 生态学报, 1997, 17(5): 469- 476.
[12] Tang J W, Bolstad P V, Martin J G. Soil carbon fluxes and stocks in a great lakes forest chronosequence. Global Change Biology, 2009, 15(1): 145- 155.
[13] Feldpausch T R, Rondon M A, Fernandes E C M, Riha S J, Wandelli E. Carbon and nutrient accumulation in secondary forests regenerating on pastures in central Amazonia. Ecological Applications, 2004, 14(sp4): 164- 176.
[14] De Kovel C G F, Van Mierlo A (J) E M, Wilms Y J O, Berendse F. Carbon and nitrogen in soil and vegetation at sites differing in successional age. Plant Ecology, 2000, 149(1): 43- 50.
[15] 牛莉平, 许嘉巍, 靳英华, 尹海霞, 王嫒林, 刘羽霞, 张英洁. 温带森林不同演替阶段下的土壤CO2排放通量昼间变化. 生态学报, 2017, 37(8).
[16] Ewel K C, Cropper W P Jr, Gholz H L. Soil CO2evolution in Florida slash pine plantations. I. Changes through time. Canadian Journal of Forest Research, 1987, 17(4): 325- 329.
[17] Wang C K, Bond-Lamberty B, Gower S T. Soil surface CO2flux in a boreal black spruce fire chronosequence. Journal of Geophysical Research, 2002, 107(D3): 8224.
[18] Klopatek J M. Belowground carbon pools and processes in different age stands of Douglas-fir. Tree Physiology, 2002, 22(2/3): 197- 204.
[19] Chen X W, Li B L, Lin Z S. The acceleration of succession for the restoration of the mixed-broadleaved Korean pine forests in Northeast China. Forest Ecology and Management, 2003, 177(1/3): 503- 514.
[20] Deng X Z, Jiang Q O, Zhan J Y, He S J, Lin Y Z. Simulation on the dynamics of forest area changes in Northeast China. Journal of Geographical Sciences, 2010, 20(4): 495- 509.
[21] Luo Y Q, Wan S Q, Hui D F, Wallace L L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 2001, 413(6856): 622- 625.
[22] Saiz G, Byrne K A, Butterbach-Bahl K, Kiese R, Blujdes V, Farrell E P. Stand age-related effects on soil respiration in a first rotation Sitka spruce chronosequence in central Ireland. Global Change Biology, 2006, 12(6): 1007- 1020.
[23] 史宝库, 金光泽, 汪兆洋. 小兴安岭5种林型土壤呼吸时空变异. 生态学报, 2012, 32(17): 5416- 5428.
[24] 高伟峰, 史宝库, 金光泽. 模拟氮沉降对典型阔叶红松林土壤呼吸的影响. 南京林业大学学报: 自然科学版, 2016, 40(1): 8- 14.
[25] Wang C K, Yang J Y. Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests. Global Change Biology, 2007, 13(1): 123- 131.
[26] 杨金艳, 王传宽. 东北东部森林生态系统土壤呼吸组分的分离量化. 生态学报, 2006, 26(6): 1640- 1647.
[27] Yan J H, Wang Y P, Zhou G Y, Zhang D Q. Estimates of soil respiration and net primary production of three forests at different succession stages in South China. Global Change Biology, 2006, 12(5): 810- 821.
[28] Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B: Chemical and Physical Meteorology, 1992, 44(2): 81- 99.
[29] Russell C A, Voroney R P. Carbon dioxide efflux from the floor of a boreal aspen forest. I. Relationship to environmental variables and estimates of C respired. Canadian Journal of Soil Science, 1998, 78(2): 301- 310.
[30] 范跃新, 杨玉盛, 郭剑芬, 杨智杰, 陈光水, 谢锦升, 钟小剑, 徐玲琳. 中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化. 植物生态学报, 2014, 38(11): 1155- 1165.
[31] Savin M C, Görres J H, Neher D A, Amador J A. Biogeophysical factors influencing soil respiration and mineral nitrogen content in an old field soil. Soil Biology and Biochemistry, 2001, 33(4/5): 429- 438.
[32] Luan J W, Liu S R, Wang J X, Zhu X L, Shi Z M. Rhizospheric and heterotrophic respiration of a warm-temperate oak chronosequence in China. Soil Biology and Biochemistry, 2011, 43(3): 503- 512.
[33] Li H J, Yan J X, Yue X F, Wang M B. Significance of soil temperature and moisture for soil respiration in a Chinese mountain area. Agricultural and Forest Meteorology, 2008, 148(3): 490- 503.
[34] Conant R T, Dalla-Betta P, Klopatek C C, Klopatek J M. Controls on soil respiration in semiarid soils. Soil Biology and Biochemistry, 2004, 36(6): 945- 951.
[35] Reichstein M, Tenhunen J D, Roupsard O, Ourcival J M, Rambal S, Dore S, Valentini R. Ecosystem respiration in two Mediterranean evergreen Holm Oak forests: drought effects and decomposition dynamics. Functional Ecology, 2002, 16(1): 27- 39.
[36] Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 1998, 4(2): 217- 227.
[37] Xu M, Qi Y. Soil-surface CO2efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Global Change Biology, 2001, 7(6): 667- 677.
[38] Taggart M, Heitman J L, Shi W, Vepraskas M. Temperature and water content effects on carbon mineralization for sapric soil material. Wetlands, 2012, 32(5): 939- 944.
[39] Brye K R, Riley T L. Soil and plant property differences across a chronosequence of humid-temperate tallgrass prairie restorations. Soil Science, 2009, 174(6): 346- 357.
[40] 蒋延玲, 周广胜, 赵敏, 王旭, 曹铭昌. 长白山阔叶红松林生态系统土壤呼吸作用研究. 植物生态学报, 2005, 29(3): 411- 414.
[41] Bond-Lamberty B, Wang C K, Gower S T. A global relationship between the heterotrophic and autotrophic components of soil respiration? Global Change Biology, 2004, 10(10): 1756- 1766.
[42] 康永祥, 夏国威, 刘建军, 周威, 陈光平. 秦岭小陇山锐齿栎林皆伐迹地土壤呼吸特征. 应用生态学报, 2014, 25(2): 342- 350.
[43] Irvine J, Law B E. Contrasting soil respiration in young and old-growth ponderosa pine forests. Global Change Biology, 2002, 8(12): 1184- 1194.
[44] 王娓, 汪涛, 彭书时, 方精云. 冬季土壤呼吸: 不可忽视的地气CO2交换过程. 植物生态学报, 2007, 31(3): 394- 402.
[45] Brooks P D, Schmidt S K, Williams M W. Winter production of CO2and N2O from alpine tundra: environmental controls and relationship to inter-system C and N fluxes. Oecologia, 1997, 110(3): 403- 413.
[46] Schimel J P, Clein J S. Microbial response to freeze-thaw cycles in tundra and taiga soils. Soil Biology and Biochemistry, 1996, 28(8): 1061- 1066.
[47] Paul K I, Polglase P J, Smethurst P J, O′connell A M, Carlyle C J, Khanna P K. Soil temperature under forests: a simple model for predicting soil temperature under a range of forest types. Agricultural and Forest Meteorology, 2004, 121(3/4): 167- 182.
[48] Litvak M, Miller S, Wofsy S C, Goulden M. Effect of stand age on whole ecosystem CO2exchange in the Canadian boreal forest. Journal of Geophysical Research, 2003, 108(D3): 8225.
[49] Payeur-Poirier J L, Coursolle C, Margolis H A, Giasson M A. CO2fluxes of a boreal black spruce chronosequence in eastern North America. Agricultural and Forest Meteorology, 2012, 153: 94- 105.
[50] Sun O J, Campbell J, Law B E, Wolf V. Dynamics of carbon stocks in soils and detritus across chronosequences of different forest types in the Pacific Northwest, USA. Global Change Biology, 2004, 10(9): 1470- 1481.
[51] 贾建伟, 王磊, 唐玉姝, 李艳丽, 张文佺, 王红丽, 付小华, 乐毅全. 九段沙不同演替阶段湿地土壤微生物呼吸的差异性及其影响因素. 生态学报, 2010, 30(17): 4529- 4538.
[52] 康冰, 刘世荣, 蔡道雄, 卢立华, 何日明, 高妍夏, 迪玮峙. 南亚热带不同植被恢复模式下土壤理化性质. 应用生态学报, 2010, 21(10): 2479- 2486.
[53] 周正虎, 王传宽. 生态系统演替过程中土壤与微生物碳氮磷化学计量关系的变化. 植物生态学报, 2016, 40(12): 1257- 1266.
[54] Enowashu E, Poll C, Lamersdorf N, Kandeler E. Microbial biomass and enzyme activities under reduced nitrogen deposition in a spruce forest soil. Applied Soil Ecology, 2009, 43(1): 11- 21.
猜你喜欢
防护林科技(2020年10期)2020-12-22 11:46:42
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
辽宁林业科技(2017年4期)2017-06-22 10:23:36
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
大气科学(2015年5期)2015-12-04 03:04:44
亚热带资源与环境学报(2015年3期)2015-01-22 07:05:36
