
秦川肉牛FABP3及FABP4基因SNP与肉质性状的关联性
2018-03-20 05:49余横伟桂林生昝林森西北农林科技大学动物科技学院陕西杨凌72002国家肉牛改良中心陕西杨凌72003陕西省肉牛工程技术研究中心陕西杨凌7200
西北农林科技大学学报(自然科学版) 2018年3期
余横伟,桂林生,胡 言,昝林森,2,3(西北农林科技大学 动物科技学院,陕西 杨凌7200;2国家肉牛改良中心,陕西 杨凌7200;3陕西省肉牛工程技术研究中心,陕西 杨凌 7200)
脂肪酸结合蛋白3(Fat acid binding proteins 3,FABP3)又称为心脏型脂肪酸结合蛋白(H-FABP)[1],可与血浆长链脂肪酸( Long-chain fatty acid,LCFA)结合。作为机体主要的能量来源, LCFA维持着心脏的正常功能,敲除小鼠的FABP3基因,会导致其 LCFA代谢过程发生紊乱[2]。在大鼠实验模型中发现,血清FABP3的水平与大鼠遭电击后的血压显著相关[3]。此外,在小鼠的心脏舒张过程中,FABP3基因的缺失能够明显抑制心脏对棕榈酸的吸收和氧化,而心肌细胞对葡萄糖的吸收和利用却提高了80%[4],这说明FABP3基因在维持机体内脂肪酸代谢的动态平衡方面发挥着重要的调控作用。
脂肪酸结合蛋白4(FABP4),又被称为A-FABP,同FABP3一样,也是FABPs家族的重要成员之一,其能够在各种细胞区室如过氧化物酶体、线粒体、内质网、脂质液滴、核内[5-6]结合脂肪酸和相关化合物。FABP4基因最早由Spiegelman等于1983年在脂肪细胞中检测到,与脂肪酸具有高度亲和性,参与LCFA的合成代谢[7]。FABP4基因在脂肪细胞中的表达受脂肪酸和胰岛素水平的调控[8]。以肥胖小鼠为模型,敲除FABP4基因能导致其胰岛素和血浆葡萄糖水平显著下降,且胰岛素抗性增强[9]。FABP4是肥胖相关代谢综合征的关键介质,研究表明该蛋白在2型糖尿病患者体内的表达水平显著高于正常人,导致患者血浆甘油三酯和胆固醇的水平降低,这说明胰腺β细胞受到保护,机体的脂质代谢被改善[10-11]。在3T3-L1细胞中发现,过表达FABP4基因可增强线粒体功能,进而促进脂肪细胞分化和脂质沉积,干扰FABP4基因则表现出相反的效果[12]。
鉴于FABP3和FABP4基因在机体脂质代谢过程中所发挥的关键作用,笔者推测其可以作为候选基因,用于畜禽肉用性状的选育研究中。因此,本研究以FABP3和FABP4为目标基因,检测其单核苷酸多态性(SNP),分析不同基因型及单倍型组合对秦川牛肉用新品系(以下简称“秦川肉牛”)肉用性状的影响,以期为秦川肉牛分子育种提供新的理论方法和技术途径。
1 材料与方法
1.1 血样的采集及DNA的提取
在西北农林科技大学国家肉牛改良中心和陕西省秦川肉牛良繁中心,随机选择18~24月龄健康的秦川肉牛571头,尾静脉采血10 mL/头,样品采集完毕后置于-80 ℃ 保存。使用TIANamp Blood DNA kit试剂盒(北京,天根)提取血样DNA,DNA质量和浓度分别采用0.5%琼脂糖凝胶电泳及紫外分光光度计进行检测。
1.2 FABP3和FABP4基因的PCR扩增
根据GenBank公布的牛FABP3基因序列(基因组DNA登录号为:AC_000159.1,mRNA登录号为:NM_174313.2)和FABP4的基因序列(基因组DNA登录号为:AC_000171.1,mRNA登录号为:NM_174314.2),利用Primer 5.0软件设计引物(表1),交由上海生工生物工程技术服务有限公司合成。
PCR反应体系为30.0 μL:含有核酸染料的dNTPs、TaqDNA聚合酶、10×Buffer的Mix 15.0 μL,ddH2O 11.8 μL,上游、下游引物(10 pmol/μL)各0.6 μL,模板DNA(50 ng/μL)2.0 μL。PCR反应条件如下:95 ℃预变性5 min;94 ℃ 30 s,退火30 s(退火温度见表1),72 ℃30 s,35个循环;72 ℃延伸10 min,4 ℃保存。PCR扩增产物用1.0%琼脂糖凝胶电泳检测。

表1 秦川肉牛FABP3和FABP4基因引物序列信息Table 1 Sequence information of primers of FABP3 and FABP4 genes in Qinchuan beef cattle
1.3 SNP位点的检测
将扩增的PCR产物送至上海生工生物工程技术服务有限公司进行纯化并测序,测序结果使用DNAMAN软件进行对比分析,寻找SNP位点。
1.4 秦川肉牛肉用性状指标的测定
肉用性状包括背膘厚(BF)、眼肌面积(ULA)和肌内脂肪(IMF)含量 3个指标,按照桂林生等[13]的检测方法进行测定。
1.5 数据统计与分析
根据基因型统计结果,计算基因型频率、等位基因频率、多态信息含量(PIC)、遗传杂合度(He)和有效等位基因数(Ne),并进行Hardy-Weinberg平衡适应性检验(HWE)[14]。同时,使用HAPLOVIEW3.32软件对FABP4基因的SNP位点进行连锁不平衡分析及单倍型分析。采用SPSS13.0软件中的GLM模型,分别对FABP3和FABP4基因突变位点的基因型与秦川肉牛肉用性状(背膘厚、眼肌面积和肌间脂肪含量)进行相关性分析,采用邓肯法 (Duncan’s) 进行均数间的多重比较。结果用“平均数±标准误”表示,以P<0.05 为差异显著水平进行差异显著性分析。GLM模型如下:Yijk=u+Gi+Aj+Eijk,其中Yijk为个体表型值,u为群体均值,Gi为标记基因型效应,Aj为年龄效应,Eijk为随机误差。
2 结果与分析
2.1 FABP3和FABP4基因PCR产物检测
图1、2和3为本试验3对引物的PCR扩增结果,目的片段的长度分别为505,491和521 bp。结果显示,扩增产物电泳条带清晰,特异性较好,可以用作后续测序分析。
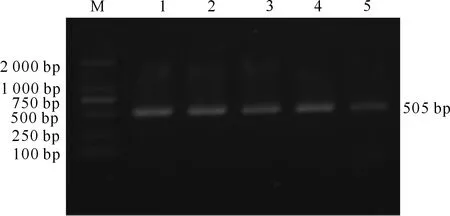
M.DNA Marker;1~5.PCR产物M.DNA Marker;1-5.PCR product图1 秦川肉牛FABP3基因部分第3内含子和部分3′非翻译区的PCR扩增Fig.1 Part of intron 3 and part of 3′UTR region individuals of FABP3 in Qinchuan beef cattle

M.DNA Marker;1~6.PCR产物M.DNA Marker;1-6.PCR product图2 秦川肉牛FABP4基因部分第1内含子和第2外显子的PCR扩增Fig.2 Part of intron 1 and exon 2 of FABP4 gene in Qinchuan beef cattleM.DNA Marker;1~5.PCR产物M.DNA Marker;1-5.PCR product图3 秦川肉牛FABP4基因部分第3内含子、第4外显子和部分3′非翻译区的PCR扩增Fig.3 Part of intron 3,exon 4 and part of 3′UTR region of FABP4 gene in Qinchuan beef cattle
2.2 FABP3和FABP4基因的SNP位点分析
牛FABP3基因位于第2号染色体上,基因全长60 606 bp,含4个外显子。本研究在FABP3基因第3内含子上检测出1个SNP,即g.60298C>T(图4)。牛FABP4基因位于第14号染色体上,基因全长4 389 bp,同样含4个外显子。本试验在FABP4基因第1内含子上检测到2个SNP,分别是g.2686A>T(图5)和g.2834C>G(图6);在第4外显子上检测出1个SNP,即g.4220A>G(图7)。在检测到的4个SNP中,g.2686A>T只有2种基因型,分别是AA和AT基因型,TT基因型缺失,其他3个位点均有3种基因型。

图4 秦川肉牛FABP3基因的SNP位点g.60298C>TFig.4 SNP (g.60298C>T) of FABP3 gene in Qinchuan beef图5 秦川肉牛FABP4基因的SNP位点g.2686A>TFig.5 SNP (g.2686A>T) of FABP4 gene in Qinchuan beef

图6 秦川肉牛FABP4基因的SNP位点g.2834C>GFig.6 SNP(g.2834C>G) of FABP4 gene in Qinchuan beef cattle图7 秦川肉牛FABP4基因的SNP位点g.4220A>GFig.7 SNP(g.4220A>G) of FABP4 gene in Qinchuan beef cattle

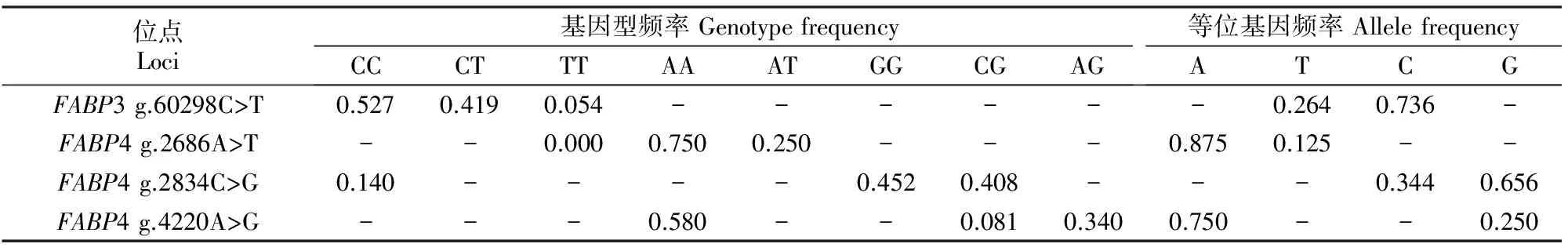
表2 秦川肉牛FABP3和FABP4基因SNP位点的基因型和等位基因分布频率Table 2 Genotype and allele frequencies of FABP3 and FABP4 genes for SNPs in Qinchuan beef cattle
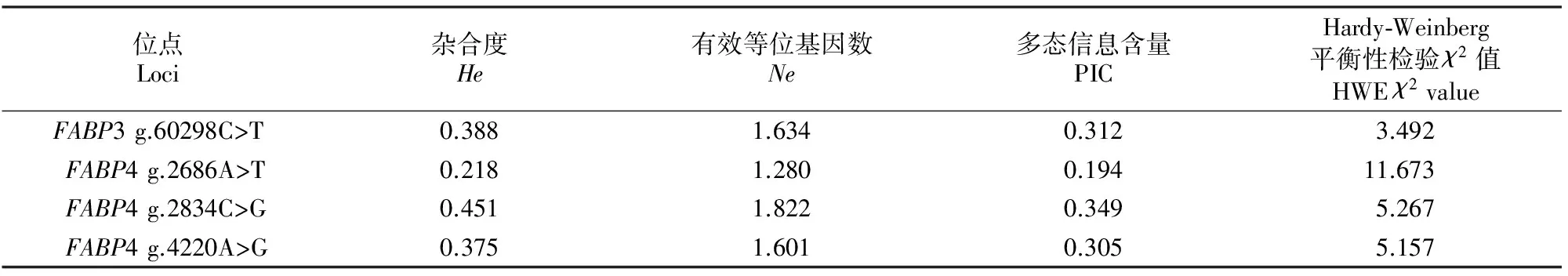
表3 秦川肉牛FABP3和FABP4基因4个SNPs的遗传参数Table 3 Genetic parameters among four SNPs loci within FABP3 and FABP4 genes in Qinchuan beef cattle
2.3 FABP3和FABP4基因SNP位点基因型对秦川肉牛肉用性状的影响
将秦川肉牛的3个肉用性状分别与FABP3基因突变位点的基因型进行关联分析。结果(表4)显示,g.60298C>T位点不同基因型个体的背膘和眼肌面积无显著差异,肌内脂肪含量差异显著(P<0.05),其中TT基因型个体显著高于CT和CC基因型个体。对FABP4基因而言,g.2834C>G位点上的CC基因型个体肉用性状较优,其眼肌面积极显著高于GG和CG基因型个体(P<0.01),肌内脂肪含量显著高于GG和CG基因型个体(P<0.05);g.4220A>G位点上的TT为优良基因型,其背膘厚显著高于CC和CT基因型(P<0.05)。
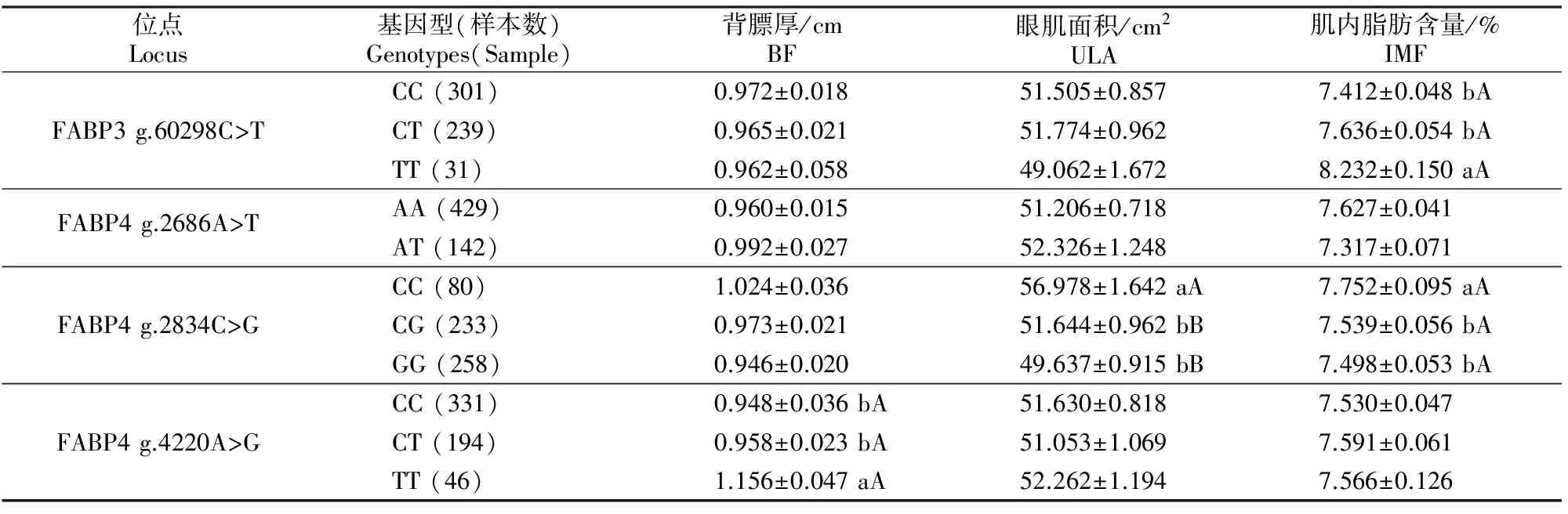
表4 FABP3和FABP4基因的SNP位点对秦川肉牛肉质性状的影响Table 4 Effects of SNPs of FABP3 and FABP4 genes on meat quality traits in Qinchuan beef cattle
注:同列数据后标不同小写字母表示差异显著(P<0.05),标不同大写字母表示差异极显著(P<0.01),标相同字母或未标字母者表示差异不显著(P>0.05)。下表同。
Notes:Different lowercase letter indicate significant difference (P<0.05),different uppercase letters indicate extremely significant difference (P<0.01),while same letters or no letter mean insignificant difference (P>0.05).The following below.
2.4 FABP4基因SNP位点的单倍型和连锁不平衡分析
通过HAPLOVIEW 3.32软件对FABP4基因上的3个SNP位点进行单倍型分析,总共检测到8种单倍型,分别为Hap1~Hap8(表5)。表5显示,单倍型Hap3的发生频率最高,为0.428;其次为单倍型Hap1和Hap4,发生频率分别为0.233和0.140。由于发生频率低于0.050的单倍型在统计学分析中没有意义,故删除单倍型Hap5、Hap6和Hap8,保留其他单倍型用于后续分析。
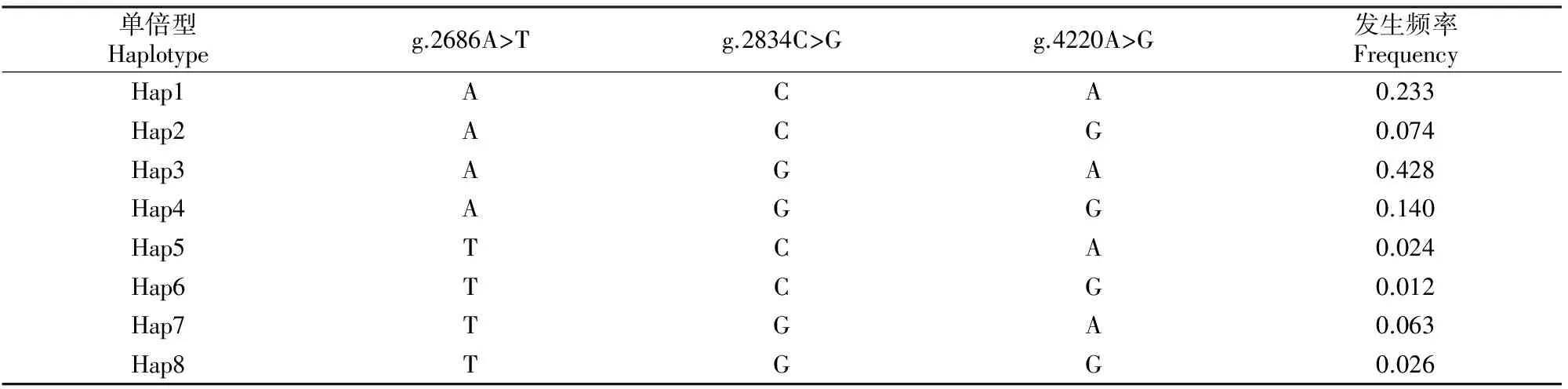
表5 秦川肉牛FABP4基因单倍型分析Table 5 Haplotypes of FABP4 genes and their frequencies in Qinchuan beef cattle
对FABP4基因中的g.2686A>T、g.2834C>G、g.4220A>G 3个SNP位点两两进行连锁不平衡分析,结果r2值均小于0.330(表6)。基于r2值大于0.330,表明2个SNP位点间具有较强的连锁不平衡效应的原则[15],可知以上3个SNP位点间不存在强的连锁性。
2.5 FABP4基因SNP位点单倍型组合对秦川肉牛肉用性状的影响
经分析得出,FABP4基因中3个SNP位点共有19种单倍型组合,将发生频率低于5.00%的不具有统计分析意义的12种单倍型组合删除,对剩余的7个单倍型组合进行关联性分析,结果见表7。由表7可知,7种单倍型组合背膘厚无显著差异;单倍型组合H1H1个体的眼肌面积极显著高于单倍型组合H3H3个体(P<0.01),并且显著高于单倍型组合H1H3、H1H4、H1H7和H3H4个体(P<0.05);肌内脂肪含量方面,单倍型组合H1H1个体极显著高于单倍型组合H3H7(P<0.01)。因此,在本试验群体中,H1H1可被视为一种最为优秀的单倍型组合。

表6 秦川肉牛FABP4基因SNP位点的连锁不平衡分析r2值Table 6 Estimated values of LD for SNPs of FABP4 gene in Qinchuan beef cattle
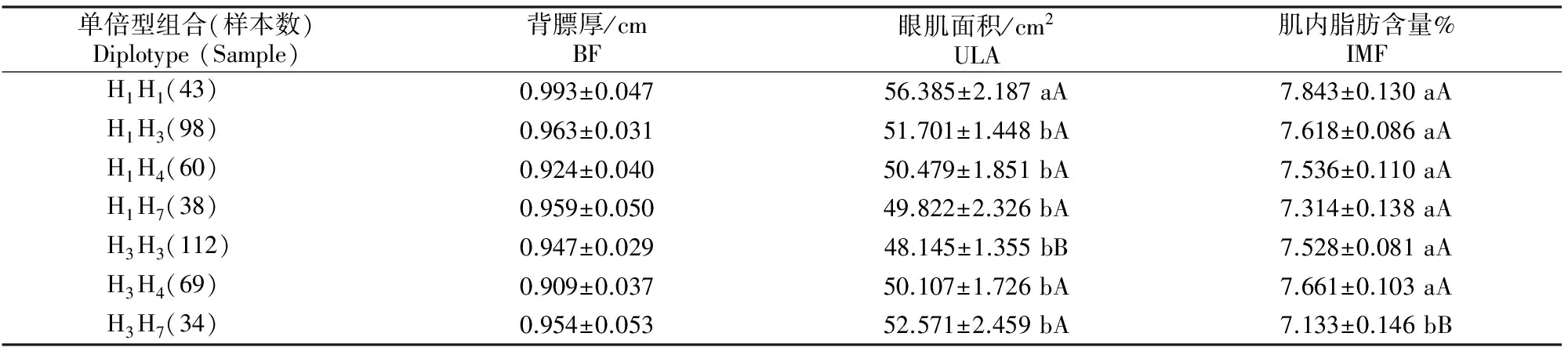
表7 FABP4基因单倍型组合对秦川肉牛肉质性状的影响Table 7 Effects of Diplotype of FABP4 genes on meat quality traits in Qinchuan beef cattle
3 讨 论
FABP3基因在动物的心脏、骨骼肌和乳腺组织中呈现高水平表达,参与脂肪酸的转运,促进脂肪酸的合成,对脂肪沉积、转运及基因的转录具有重要的调控作用[16]。在H-FABP基因中鉴定的新SNP位点与北京油鸡的大腿和胸肌的IMF含量相关[17]。Cho等[18]克隆出猪FABP3基因的mRNA,并检测出了6个SNP位点,通过关联性分析发现,位于第2外显子上编码80号位氨基酸的G>A突变可显著影响背膘厚。同样以猪为研究对象,Li等[19]在FABP3基因的第2内含子上发现了4个SNP位点,其均与背最长肌的嫩度、含水量和风味密切相关。Blecha等[20]在肉牛FABP3基因中检测出3个SNP位点,其中位点rs109315289显著影响牛的眼肌面积。本试验对秦川牛FABP3基因全长进行了多态性分析,结果在其第3内含子上发现1个SNP位点g.60298C>T,该位点不同基因型的肌内脂肪含量差异显著(P<0.05),基因型TT个体显著高于基因型CT和CC。
作为膜周边蛋白,FABP4可与长链脂肪酸、脂肪氧化产物和环氧化酶等疏水性化合物相结合,参与其代谢及信号转导过程,在调控糖脂代谢和胰岛素分泌方面发挥着重要作用[21]。目前,在人和小鼠上对该蛋白研究得比较全面。Matsumotoa等[22]在FABP4基因的启动子区域发现了2个SNP位点,分别为g.-259A>G和g.-287A>G,这2个SNP位点均能影响转录因子的结合,进而影响日本和牛的大理石花纹。Gao等[23]在用PCR-SSCP研究白猪Junmu 1号的过程中发现,A-FABP位点3 481 bp的点突变对肌内脂肪(IMF)含量有显著的影响。在3个中国山羊品种中,Xu等[24]通过SSCP技术,在FABP4基因第1内含子上检测到1个SNP位点,该位点与山羊的肌内脂肪含量紧密相关。Oh等[25]在韩国牛FABP4基因外显子上检测到3个SNP位点,其均显著影响牛肉脂肪酸组成。本试验在秦川肉牛FABP4基因上检测到g.2686A>T、g.2834C>G和g.4220A>G 3个SNP位点,除g.2686A>T外,其余2个SNP位点与秦川肉牛肉用性状紧密关联,其中g.2834C>G可以显著影响眼肌面积和肌内脂肪含量,g.4220A>G则可以显著影响背膘厚。单倍型组合关联性分析发现,单倍型组合H1H1个体眼肌面积和肌内脂肪含量显著高于其余单倍型组合。
值得注意的是,本试验检测出的可显著影响秦川肉牛肉用性状的3个SNP位点(g.60298C>T、 g.2686A>T和g.2834C>G)中,g.60298C>T和g.2834C>G分别定位于FABP3和FABP4基因的内含子上,且为同义突变。这2个位点影响肉用性状的机制可能是:内含子SNP位点可能与同一染色体上其他基因上存在的SNP位点呈现完全连锁状态[26];内含子SNP位点可能影响剪接供体位点或其附近区域[27],导致翻译过程发生改变。
4 结 论
本研究发现,FABP3基因g.60298C>T和FABP4基因g.2834C>G、g.4220A>G在秦川肉牛中存在多态性,与肉用性状紧密关联。FABP4基因3个SNP位点的优势单倍型组合为H1H1(ACA-ACA),其个体眼肌面积和肌间脂肪含量极显著或显著高于其他单倍型组合,故可将FABP3和FABP4基因作为影响肉用性状的候选基因用于标记辅助选择,以加速秦川肉牛定向高效选育和群体遗传改良。
[1] 姜延志,刘晓研,李芳琼,等.猪H-FABP基因多态性及其与肌内脂肪含量的相关研究 [J].畜牧兽医学报,2010,41(7):792-796.
Jian Y Z,Liu X Y,Li F Q,et al.The genetic relationship between polymorphism ofH-FABPgene andint-ramuscular fat content in pig [J].Acta Veterinaria et Zootechnica Sinica,2010,41(7):792-796.
[2] Binas B,Danneberg H,McWhir J,et al.Requirement for the heart-type fatty acid binding protein incardiac fatty acid utilization [J].The FASEB Journal,2013,8:805-812.
[3] Caglar O,Hasim A,Cevat Y,et al.Heart-type fatty acid binding protein and cardiac- troponin I may have a diagnostic value in electrocution:A rat model [J].Journal of Forensic and Legal Medicine,2016,39:76-79.
[4] Chaap F G,Binas B,Danneberg H,et al.Impaired long-chain fatty acid utilization by cardiamyo-cytes isolated from mice lacking the heart-type fatty acid binding protein gene [J].Circulation research,1985,4:329-337.
[5] Kralisch S,Fasshauer M.Adipocyte fatty acid binding protein:a novel adipokine involved in the pathogenesis of metabolic and vascular disease? [J].Diabetologia,2013,1:10-21.
[6] Furuhashi M,Furuhashi M,Saitoh S,et al.Fatty acid-binding protein 4 (FABP4):pathophysiological insights and potent clinical biomarker of metabolic and cardiovascular diseases [J].Clinical Medicine Insights:Cardiology,2015,3:23-33.
[7] Piegelman B M,Frank M,Green H.Molecular cloning of mRNA from 3T3 adipocytes regulation of mRNA content for glycerop hosphate dehydrogenase and other differentiation-dependent proteins during adipocyte development [J].The Journal of Biologica Chemistry,1983,258(16):10083-10089.
[8] Haunerland N H,Spener F.Fatty acid-binding proteins-insights from genetic manipulations [J].Progress in Lipid Research,2004,43(4):328-349.
[9] Hotamisligil G S,Johnson R S,Distel R J,et al.Uncoupling of obesity from insulin resistance through a targeted mutation in aP2,the adipocyte fatty acidbinding protein [J].Science,1996,274(5291):1377-1379.
[10] Young C K,Yong K C,Won Y L,et al.Serum adipocyte-specific fatty acidbinding protein is associated with nonalcoholic fatty liver disease in apparently healthy subjects [J].The Journal of Nutritional Biochemistry,2011,22(3):289-292.
[11] Ysal K T,Scheja L,Wiesbrock S M,et al.Improved glucose and lipid metabolism in genetically obese mice lacking aP2 [J].Endocrinology,2000,141(9):3388-3396.
[12] 谈 笑.FABP4对3T3-L1脂肪细胞中脂肪酸代谢和线粒体功能的影响 [D].陕西杨凌:西北农林科技大学,2014.
Tan X.Regulation of FABP4 on fatty acid metabolism and mitochondrial function in 3T3-L1 adipocytes [D].Yangling,Shaanxi:Northwest A&F University,2014.
[13] 桂林生,昝林森,王洪宝,等.SIRT1和SIRT2基因多态性及合并基因型与秦川牛肉用性状的关联分析 [J].畜牧兽医学报,2015,46(10):1741-1749.
Gui L S,Zan L S,Wang H B,et al.Effects of single and combined genotypes ofSIRT1 andSIRT2 genes on meat quality traits in Qinchuan cattle [J].Chinese Journal of Animal and Veterinary Sciences,2015,46(10):1741-1749.
[14] Hardy H G.Mendelian proportions in a mixed population [J].Science,1908(28):49-50.
[15] Flint G S A.Structure of linkage disequilibrium in plants [J].Annu Rev Plant Biol,2003(54):357-374.
[16] 李 娟,杜新华,陈 燕,等.牛FABP3转基因小鼠的遗传及表达特性研究 [J].畜牧兽医学报,2015,46(10):1750-1758.
Li J,Du X H,Chen Y,et al.Study on genetic characteristic and expression of bovineFABP3 gene in transgenic mice [J].Chinese Journal of Animal and Veterinary Sciences,2015,46(10):1750-1758.
[17] Ye M H,Chen J L,Zhao G P,et al.Associations of A-FABP and H-FABP markers with the contnt of intramuscular fat in Beijing-You chicken [J].Anim Biotechnol,2010,21(1):14-24.
[18] Cho K H,Kim M J,Jeon G J,et al.Association of genetic variants forFABP3 gene with back-fat thickness and intramuscular fat content in pig [J].Molecular Biology Reports,2011,38:2161-2166.
[19] Li X P,Kim S W,Choi J S,et al.Investigation of porcineFABP3 andLEPRgene polymorphisms and mRNA expression for variation in intramuscular fat content [J].Molecular Biology Reports,2010,37:3931-3939.
[20] Blecha I M Z,Siqueira F,Ferreira A B R,et al.Identification and evaluation of polymorphisms inFABP3 andFABP4 in beef cattle [J].Genetics and Molecular Research,2015,14(4):16353-16363.
[21] Zhang J,Yang J H,Huang Y L,et al.Effects of different diet forms on growth performance and nutrient digestibility in hepu Geese [J].Chinese Journal of Animal Nutrition,2010,22(3):586-594.
[22] Matsumotoa H,Nogib T,Tabuchib I,et al.The SNPs in the promoter regionsof the bovineFADS2 andFABP4 genes are associated with beef quality traits [J].Livestock Science,2014,163:34-40.
[23] Gao Y,Zhang Y H,Zhang S,et al.Association ofA-FABPgene polymorphism in intron 1 with meat quality traits in Junmu No.1 white swine [J].Gene,2011,11:170-173.
[24] Xu Q L,Tang G W,Zhang Q L,et al.TheFABP4 gene polymorphism is associated with meat tenderness in three Chinese native sheep breeds [J].Czech Journal of Animal Science,2011,56:1-6.
[25] Oh D Y,Lee Y S,La B M,et al.Identification of the SNP (Single Nucleotide Polymorphism) for fatty acid composition associated with beef flavorrelatedFABP4 (Fatty Acid Binding Protein 4) in Korean cattle [J].Animal Science,2012,25:913-920.
[26] Li M X,Sun X M,Hua L S,et al.SIRT1 gene polymorphi-sms are associated with growth traits in Nanyang cattle [J].Molecular and Cellular Probes,2013,27:215-220.
[27] Cao F,Zhang H,Feng J,et al.Association study of three microsatellite polymorphisms located in introns 1,8,and 9 ofDISC1 with schizophrenia in the Chinese Han population [J].Genet Test Mol Biomarkers,2013,17:407-411.
猜你喜欢
今日农业(2022年2期)2022-11-16
今日农业(2022年1期)2022-06-01
做人与处世(2022年6期)2022-05-26
做人与处世(2022年4期)2022-05-26
情感读本·道德篇(2022年2期)2022-04-12
今日农业(2021年21期)2021-11-26
作文大王·低年级(2021年8期)2021-09-14
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01
湖南饲料(2019年5期)2019-10-15
生物工程学报(2019年6期)2019-07-10
