
马尾松木荷不同比例混交林植被碳密度特征
2018-03-19 03:18:20赖国桢欧阳勋志向云西吴自荣
中南林业科技大学学报 2018年2期
赖国桢,曹 梦,潘 萍,欧阳勋志,国 瑞,向云西,吴自荣
(江西农业大学 林学院,江西 南昌 330045)
森林生态系统是陆地生态系统的主体,也是最大的陆地碳库,约占全球陆地总碳库的46%,在全球碳循环和减缓气候变暖中起着重要作用[1]。20世纪90年代以来,森林生态系统碳密度研究取得显著进展,相关研究表明影响森林碳密度的因素主要有森林类型、林龄、立地条件、人为干扰[2-7]等,合理的经营措施及科学的森林管理能够有效的提高森林的碳储量及碳密度[8]。
马尾松 Pinus massoniana 是亚热带主要针叶树种,具有适应性强、耐干旱与贫瘠的特点。不少学者从不同角度对马尾松混交林进行了研究,如王力[9]、殷沙[10]、黎芳[11]等分别对不同混交比例的马尾松林细根生物量及养分含量、凋落物及土壤持水性能以及林下植被多样性进行了比较研究,冯随起[12]对马尾松混交林的混交比例选择进行了探讨,陈模芳等[13]对不同群落类型的马尾松林地上部分生物量与碳储量进行了比较研究。赣南曾是我国水土流失较严重的地区之一,尤其是该区的兴国县,水土流失范围广、强度大,曾被称为“江南沙漠”。为恢复森林植被控制水土流失,20世纪70—90年代进行大面积的飞播马尾松种子造林,并且在90年代对部分纯林进行了不同比例补植木荷 Schima superba 的试验,而不同比例组成的混交林其植被碳密度有何差异目前尚不明确。因此,本文以赣南飞播马尾松造林具有代表性的兴国县为研究区,探讨马尾松、木荷不同混交比例下其植被碳密度的变化规律,为马尾松林开展碳汇林业的科学经营管理等方面提供参考依据。
1 研究区概况
兴国县位于江西省中南部,赣州市北部,地形以低丘、丘陵为主,最低海拔为127.9 m,最高海拔为1 204 m,主要土壤类型为红壤;属于亚热带季风湿润气候带,年平均气温18.8 ℃,平均无霜期284 d左右,年平均降水量为1 560 mm;主要森林类型为针叶林、针阔混交林、常绿阔叶林等,森林资源覆盖率达72.2%;据兴国县林业局相关统计资料,1973年至2001年间该县共飞播造林77 633.3 hm2,保存面积为64 000 hm2,约占全县现有有林地面积的29.5%[14]。
2 研究方法
2.1 标准地设置
由于研究区现保存下来的飞播马尾松林主要是20世纪80年代后期到90年代飞播的,多为中龄林(21~30 a),且主要集中在中低丘地貌(海拔100 ~ 300 m),90年代为优化林分结构,对部分飞播纯林(以中下坡为主)进行了不同比例木荷的补植试验。因此,本研究在中低丘分布区选择人为干扰程度较低且林分密度、立地条件等相似的中龄林作为研究对象,依据当时补种木荷的情况,并考虑与纯林对照,将飞播马尾松林按其混交比例分为类型Ⅰ(10马)、类型Ⅱ(8马2木)、类型Ⅲ(7马3木)、类型 Ⅳ(6马4木)4 种类型,同一类型的林分设置3个标准地,共12个,标准地面积均为900 m2(30 m×30 m)。各标准地成土母岩均为花岗岩,土壤类型为红壤,土层厚度>80 cm,坡位为中下坡或下坡,坡向为半阳或阳坡。由于所选标准地中胸径小于 5 cm的林木所占比例较少,有个别标准地还零星分布有少量的杉木 Cunninghamia lanceolate 和枫香 Liquidambar formosana,且二者所占比例均较小,因此,本研究将杉木归类为马尾松,枫香归类为木荷。各类型标准地基本概况见表1。
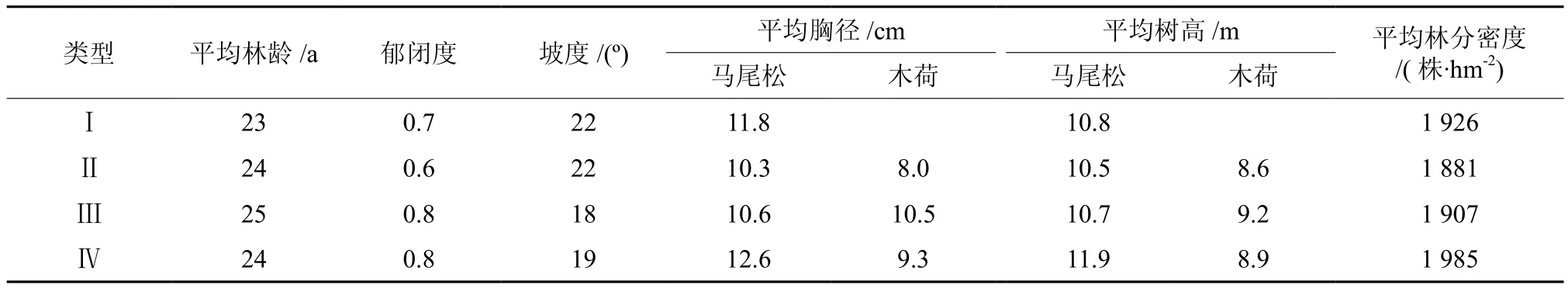
表 1 标准地基本概况Table 1 The basic situation of sample plots
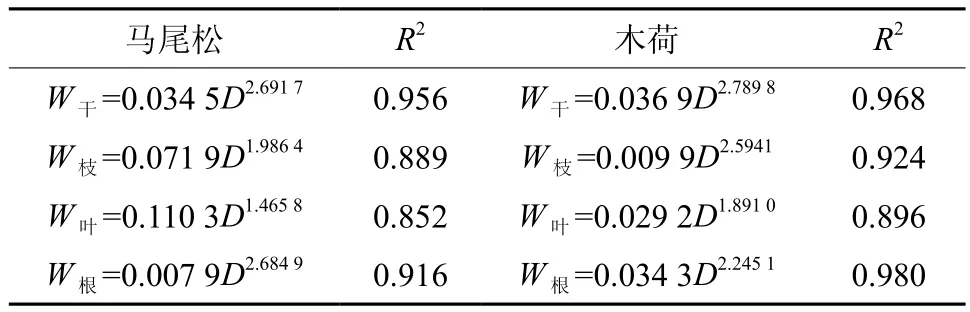
表 2 马尾松和木荷异速生长方程Table 2 The allometric growth equation of Pinus massoniana and Schima superba
2.2 植被碳密度计算方法
(1)乔木层生物量调查与碳密度计算。在标准地内进行每木检尺,检尺标准为胸径≥5 cm的树木,调查内容包括乔木的胸径、树高、林分郁闭度等。乔木层生物量根据每木检尺的结果,采用与乔木相对应的异速生长模型:

式中,W为单株乔木各个器官生物量(kg·株-1);D为胸径(cm);a,b为相关参数,以此方程求得单株木各器官(干、枝、叶、根)生物量,各器官总量相加得单株乔木生物量,标准地内乔木层总生物量即为各株生物量的总和。本研究分别采用肖欣和程煜[15-16]等拟合结果估算马尾松和木荷的生物量。
森林碳密度的普遍算法是用森林植被生物量乘以碳含量,因此植被的碳含量是研究森林碳密度的关键。目前对区域森林碳密度研究通常使用国际通用碳含量(0.45 ~ 0.5),此方法在针对大尺度的森林碳密度估算上行之有效,然而在较小的林分尺度,由于不同种类、不同年龄的植物,甚至同一植物的不同器官的碳含量都略有差别,使用国际通用碳含量会大大的增加实验的误差。因此,本研究采用王兵等人[17]测定的江西省大岗山林区马尾松及木荷各器官的碳含量计算其碳密度。
(2)林下植被生物量调查与碳密度计算。根据标准地具体情况,在标准地内分别设置3个2 m×2 m的灌木样方、3个1 m×1 m的草本样方和凋落物样方,调查并记录样方内灌木、草本的种类、株(丛)数和平均高等因子,采用收获法测定其生物量,灌木分为枝、叶、根,草本分为地上和地下,凋落物分为半分解(凋落物原状不完整,多分解成碎屑)和未分解(凋落物保持原状,外表没有分解痕迹),测量其鲜质量,并分别取样带回实验室,将样品置于80 ℃恒温箱直至其质量恒定,通过样品干质量与鲜质量可以计算出样品含水量,进而可以算出整个样方的生物量,最后推算出每块标准地的总生物量。将烘干后的样品磨成粉末,采用重铬酸钾氧化-外加热法测定样品中的碳含量,碳含量测定结果如表3所示,用所测得生物量乘以碳含量即得出碳密度。

表 3 林下植被层及凋落物层各组分碳含量†Table 3 The understory vegetation layer and litter layer carbon content of different components (g·kg-1)
2.3 数据处理
采用SPSS 21.0中对数据进行方差分析及相关性分析,方差分析差异显著时采用LSD法进行二次检验,采用Excel 2003进行数据统计及图表制作。
3 结果与分析
3.1 乔木层碳密度
如表4所示,各类型的飞播马尾松林干、枝、叶根碳密度范围分别为20.64 ~ 35.64、7.13 ~8.94、3.02 ~ 3.75、4.28 ~ 7.70 t·hm-2。可以看出,随着木荷混交比例的提高,树干和树根的碳密度均随之增大,而树枝和树叶碳密度则表现为先减小后增大。各组分分配比随着混交比例的变化也不尽相同,其中树干和树根所占比例随着混交比例的提高而增大,树枝与树叶分配比则随混交比例提高而表现出逐步下降的趋势。乔木层总碳密度则随混交比例的提高而依次增大,表现为类型Ⅳ>Ⅲ>Ⅱ>Ⅰ。经方差分析表明,不同混交比例对马尾松的干、枝、叶以及乔木层的碳密度均无显著影响(P>0.05),但树根的碳密度有显著差异(P<0.05);使用LSD法进行二次检验发现,类型Ⅳ树根碳密度显著高于类型Ⅰ、Ⅱ(P<0.05),而与类型Ⅲ两者间无显著差异(P>0.05),类型Ⅰ、Ⅱ、Ⅲ两两间无显著差异(P>0.05)。
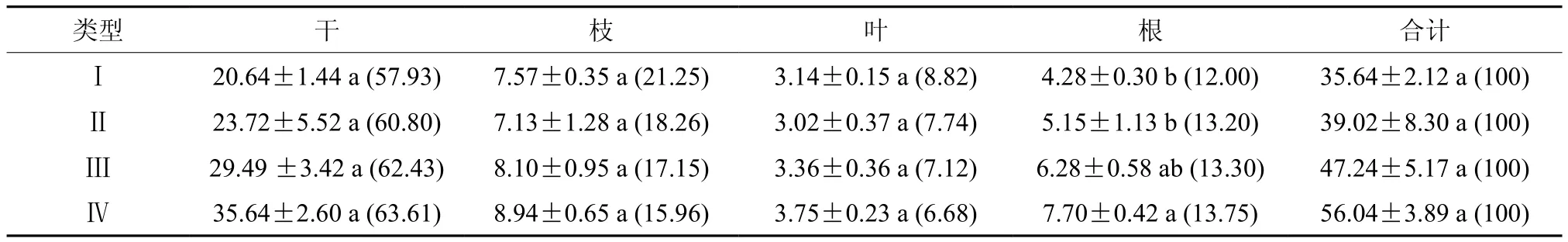
表 4 乔木层各组分碳密度及分配†Table 4 The tree layer carbon density and distribution at different organs t·hm-2,%
3.2 林下植被层碳密度
由表5可知,各类型灌木层碳密度为0.04 ~0.15 t·hm-2, 草 本 层 各 类 型 碳 密 度 为 0.57 ~0.77 t·hm-2;灌木层碳密度均随着混交比例的提高表现为先增大后减小的趋势,而草本层碳密度则随着混交比例的提高而相应减小。林下植被层总碳密度为0.61 ~ 0.86 t·hm-2,随着混交比例的提高林下植被层碳密度表现为类型Ⅰ>Ⅱ>Ⅲ>Ⅳ。灌木层碳密度占林下植被层碳密度的比例都较小,仅为6.58% ~ 19.02%,草本层为80.98% ~93.42%。方差分析表明,不同类型相同层次间碳密度均不存在显著性差异(P>0.05)。
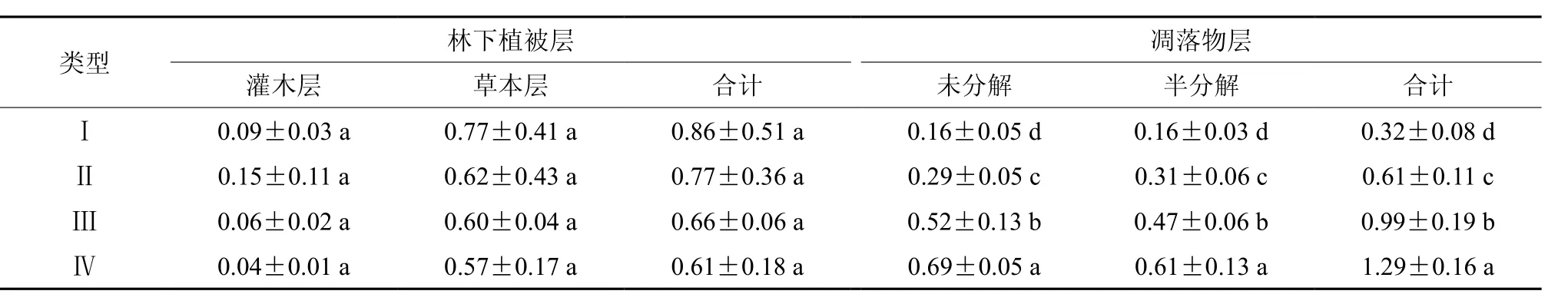
表 5 林下植被层及凋落物层碳密度†Table 5 The carbon density of understory vegetation layer and litter layer t·hm-2
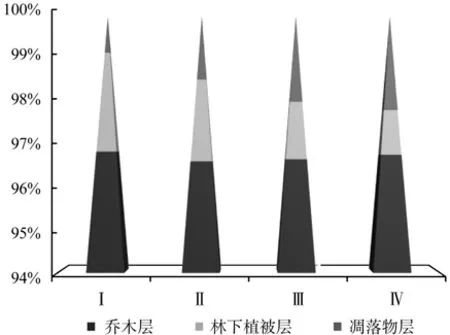
图1 植被碳密度分配Fig.1 The distribution of carbon density in vegetation
3.3 凋落物层碳密度
从表5可以看出,各类型凋落物未分解层碳密度为0.16 ~ 0.69 t·hm-2,半分解层凋落物碳密度为0.16 ~ 0.61 t·hm-2,不同分解层凋落物碳密度均表现为随着木荷混交比例的提高而增大的趋势。凋落物层总碳密度为0.32 ~ 1.29 t·hm-2,由大到小依次为类型Ⅳ>Ⅲ>Ⅱ>Ⅰ。经方差分析表明,不同类型同一分解层凋落物碳密度两两间均存在显著性差异(P<0.05)。
3.4 植被碳密度
如图1所示,各类型飞播马尾松林植被碳密度分别为 36.82、40.40、48.89 以及 57.94 t·hm-2,与乔木层碳密度相类似,即植被碳密度表现出随混交比例的提高而增大的趋势,究其原因是因为乔木层在植被碳密度中占主要部分,起到了决定性的作用。乔木层碳密度所占比重均在96%以上,远远高于林下植被层及凋落物层所占比重;随着混交比例的提高林下植被层所占比重相应减小,而凋落物层所占比重逐步增大。使用SPSS对不同类型的植被碳密度进行单因素方差分析,结果表明,不同的混交比例对植被碳密度无显著影响(P > 0.05)。
为了解木荷比例与林分各层碳密度关系,将木荷比例与林分各层碳密度及总碳密度进行相关性分析,见表6。结果表明,木荷比例与乔木层及林分总碳密度呈显著正相关(P<0.05),与凋落物层碳密度呈极显著正相关(P<0.01),与林下植被层碳密度相关性不显著(P>0.05)。

表 6 木荷比例与林分各层的相关性分析†Table 6 Correlations between the proportions of Schima superba and carbon density of different layers
4 结论与讨论
本研究表明,树干和树根碳密度随着混交比例的提高而增大,树枝和树叶碳密度则表现为先减小后增大;从碳密度分配上发现,树干和树根所占比例与碳密度的变化状况一致,而树枝与树叶分配比却表现为随着混交比例的提高而减小。出现这一现象的原因可能是马尾松木荷混交在促进乔木树干和树根生长的同时也影响了其自然整枝的强度,自然整枝的强度直接影响枝、叶的生物量,而自然整枝与混交比例及方式之间存在着较复杂关系。4种类型飞播马尾松林乔木层碳密度范围为35.64 ~ 56.04 t·hm-2,其中类型Ⅳ>Ⅲ>Ⅱ>Ⅰ,马尾松与木荷混交林的乔木层碳密度均大于马尾松纯林,这与一些学者的研究得出的混交林的碳密度大于纯林结果一致[18-19]。高必垒等[20]对不同混交比例的马尾松针阔混交林碳储量研究中表明马尾松所占比例在60% ~ 100%范围内乔木层碳密度随着阔叶树种混交比例的提高而逐步增大,此结果与本研究基本一致。可见,阔叶树比例的提高能够有效提高林分乔木层碳密度,这可能是因为马尾松木荷混交能够有效改善林分结构及凋落物组成,加速凋落物分解,促进部分酶的活性,从而提高林分的生产力[21]。
郭琦等[22]对杉木纯林以及杉木-火力楠Michelia macclurei Dandy 混交林林下植被生物量比较得出混交林灌草生物量均远大于纯林;汪永文等[23]对5种混交比例的马尾松与阔叶混交林研究发现,随着马尾松混交比例的增加,林下植被生物量呈现逐渐增大后降低趋势,当阔叶树占4成时林下植被生物量达到最大值。而本研究结果表明林下植被层碳密度随着木荷混交比例的提高而减小,即类型Ⅰ>Ⅱ>Ⅲ>Ⅳ,这可能是由于飞播马尾松林土壤种子库的物种数少而且主要是禾本科、莎草科的草本物种[24],随着木荷比例的增加,枯枝落叶增多而不利于土壤种子库现存物种的更新,同时,补植间伐等人为活动对林下植被造成一定程度的干扰,直接影响林下植被层生物量,从而导致林下植被碳密度的下降[25]。
凋落物层碳密度中类型Ⅰ最小,且表现为随木荷混交比例的提高而增大,即类型Ⅳ>Ⅲ>Ⅱ>Ⅰ。李红振等[26]对比研究了不同类型的白桦落叶松混交林,得出随着白桦所占比例的提高,混交林内凋落物层碳密度相应增大;殷沙等[10]对不同混交比例马尾松林凋落物进行了研究,得出随着混交比例的提高凋落物现存量逐步增大,碳密度也随之而增大,这与本研究结果一致。方差分析表明,木荷比例的差异对凋落物碳密度具有显著影响(P<0.05),随着木荷比例的提高凋落物层碳密度显著增大,这可能主要是因为木荷为常绿阔叶树种,枯枝落叶量较大,木荷比例提高导致其凋落物现存量也随之增加。
马尾松与木荷不同混交比例林分植被碳密度为36.82~ 57.49 t·hm-2,混交林林分碳密度均大于马尾松纯林碳密度,表现为类型Ⅳ>Ⅲ>Ⅱ>Ⅰ,与大多数研究结果一致[18-20]。相关性分析表明,木荷比例与乔木层及林分总碳密度呈显著正相关(P<0.05),与凋落物层碳密度呈极显著正相关(P<0.01),这主要是由于木荷混交比例的提高,能有效提高了凋落物的现存量以及改善土壤性状,促进林木的生长。综上所述,在飞播马尾松林中补植木荷能有效提高乔木层及林分植被总碳密度,但由于受研究区林分条件的限制,本研究只限于木荷比例占4成以下的马尾松、木荷混交林分,而随着木荷比例的继续增大,林分植被碳密度是否继续增大尚不清楚,这有待开展进一步研究。
[1]Ruiz-Peinado R, Bravo-Oviedo A, Lopez-Senespleda E, et al. Do thinnings influence biomass and soil carbon stocks in Mediterranean maritime pine woods?[J]. European Journal of Forest Research, 2013, 132(2): 253-262.
[2]胡 青, 汪思龙, 陈龙池, 等. 湖北省主要森林类型生态系统生物量与碳密度比较[J]. 生态学杂志, 2012,31(7):1626-1632.
[3]杨玉姣, 陈云明, 曹 扬. 黄土丘陵区油松人工林生态系统碳密度及其分配[J]. 生态学报, 2014, 34(8): 2128-2136.
[4]Cheng Chihhsin, Hung Chihyu, Chen Chioupeng, et al. Biomass carbon accumulation in aging Japanese cedar plantations in Xitou, Central Taiwan [J]. Botanical Studies , 2015, 54(1): 60-68.
[5]成向荣, 虞木奎, 吴统贵, 等. 立地条件对麻栎人工林碳储量的影响[J]. 生态环境学报, 2012, 21(10): 1674-1677.
[6]明安刚, 张治军, 谌红辉, 等. 抚育间伐对马尾松人工林生物量与碳贮量的影响[J]. 林业科学, 2013, 49(10): 1-6.
[7]武朋辉, 党坤良, 常 伟, 等. 抚育间伐对秦岭南坡锐齿栎天然林碳密度的影响[J]. 西北农林科技大学学报(自然科学版),2016, 44(10): 75-82.
[8]Fahey T J, Woodbury P B, Battles J J, et al. Forest carbon storage:ecology, management, and policy[J]. Frontiers in Ecology and the Environment, 2010, 8(5): 245-252.
[9]王 力, 汪永文, 张令峰, 等. 不同混交比例马尾松林细根生物量及其养分研究[J].安徽农业大学学报,2010,37(2):317-323.
[10]殷 沙, 赵 芳, 欧阳勋志. 马尾松木荷不同比例混交林枯落物和土壤持水性能比较分析[J]. 江西农业大学学报, 2015,37(3): 454-460.
[11]黎 芳, 潘 萍, 宁金魁, 等. 马尾松—木荷不同比例混交林林下植被物种组成及其多样性分析[J]. 西北林学院学报,2016, 31(6): 34-40.
[12]冯随起. 闽粤栲与马尾松混交比例选择及混交效果评价[J].福建林业科技, 2010, 37(3): 45-49.
[13]陈模芳, 丁贵杰, 瞿帅帅, 等. 不同马尾松群落类型的生物量及碳储量[J]. 中南林业科技大学学报, 2016, 36(7): 76-80.
[14]丁 松, 应学亮, 吕 丹, 等. 赣南飞播马尾松林林下植被盖度对土壤质量的影响[J]. 水土保持研究, 2014, 21(3): 31-36.
[15]肖 欣. 马尾松飞播林与天然林碳密度时空分布特征比较研究[D]. 南昌: 江西农业大学, 2015.
[16]程 煜, 洪 伟, 吴承祯, 等. 木荷地上部分生物量分布特征与生产力[J]. 应用与环境生物学报, 2009, 15(3): 318-322.
[17]王 兵, 魏文俊. 江西省森林碳储量与碳密度研究[J]. 江西科学, 2007, 25(6): 681-687.
[18]邹文魁. 马尾松、木荷混交林生物量结构研究[J]. 北华大学学报(自然科学版), 2008, 9(2): 161-164.
[19]何友均, 覃 林, 李智勇, 等. 西南桦纯林与西南桦×红椎混交林碳贮量比较[J]. 生态学报, 2012, 32( 23) : 7586-7594.
[20]高必垒, 陈 清, 冯 春, 等. 混交比例对马尾松针阔混交林植被碳贮量的研究[OL]. 中国科技论文在线, http://www.paper.edu.cn/releasepaper/content/201304-558.
[21]林惠章, 郭孝玉, 欧阳勋志, 等. 飞播马尾松纯林补植木荷后土壤酶活性与土壤养分变化[J]. 东北林业大学学报, 2015,43(7): 136-139.
[22]郭 琦, 王新杰. 不同混交模式杉木人工林林下植被生物量与土壤物理性质研究[J]. 中南林业科技大学学报, 2014, 34(5):70-74.
[23]汪永文, 王 力, 王丽丽, 等. 马尾松混交林林下植被结构及生物量特征研究[J]. 安徽农业大学学报, 2010,37(2):312-316.
[24]陈颖颖, 吴自荣, 潘 萍, 等. 飞播马尾松林土壤种子库的萌发特征及其与土壤理化性质的关系[J]. 土壤通报, 2016,47(1): 92-97.
[25]何圣嘉, 谢锦升, 周艳翔, 等. 南方红壤侵蚀区马尾松林下植被恢复限制因子与改造技术[J]. 水土保持通报, 2013, 33(3):118-124.
[26]李红振, 李凤日, 贾炜玮, 等. 大兴安岭不同类型白桦落叶松混交林枯落物水源涵养功能[J]. 东北林业大学学报, 2014,42(6): 43-46.
猜你喜欢
安徽农学通报(2023年16期)2023-10-15 11:49:37
内蒙古林业(2022年8期)2022-09-08 01:39:58
中国林副特产(2022年4期)2022-08-08 05:50:30
散文诗世界(2022年5期)2022-05-23 02:02:55
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
防护林科技(2019年12期)2019-04-14 04:01:18
新农民(2019年11期)2019-02-20 02:19:00
现代园艺(2018年2期)2018-03-15 08:01:03
陕西林业科技(2017年5期)2017-12-14 10:50:32
花卉(2016年14期)2016-07-31 18:46:24
